Cinesina
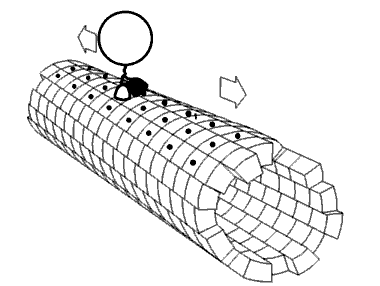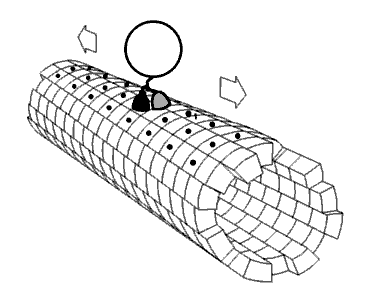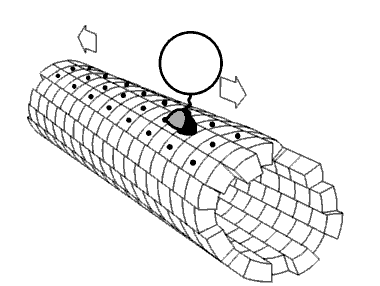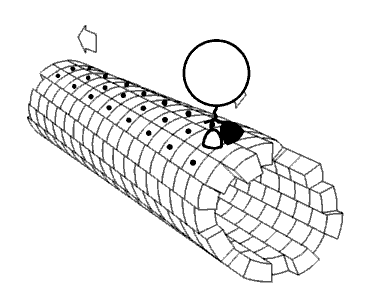

As cinesinas ou quinesinas (ás veces escrito kinesinas, do grego kinetos, móbil) son un tipo de proteínas motoras presentes nas células eucarióticas. As cinesinas móvense ao longo dos microtúbulos do citoesqueleto movidas pola enerxía fornecida pola hidrólise do ATP, polo que as cinesinas son ATPases. O movemento activo das cinesinas é necesario para diversas funcións celulares, como a mitose, meiose e o transporte de cargamentos pola célula, como no transporte axonal. A maioría das cinesinas camiñan cara ao extremo máis (+) dos microtúbulos, ao cal xeralmente significa que transportan o seu cargamento desde a parte central da célula cara á periferia. Esta forma de transporte coñécese como transporte anterógrado. Outras proteínas motoras, como a dineína, polo contrario, móvense cara ao extremo menos (-) dos microtúbulos.
As cinesinas
[editar | editar a fonte]As cinesinas descubríronse como motores de transporte intracelular anterógrado baseado nos microtúbulos.[1] O primeiro membro que se coñeceu desta familia foi a cinesina-1, illada como un motor de transporte de orgánulos axonal rápido heterotetramérico, formado por dúas subunidades motoras idénticas (KHC) e dúas "cadeas lixeiras" (KLC) por medio da purificación por afinidade con microtúbulos de extractos de células neuronais.[2] Posteriormente purificouse un motor baseado nos microtúbulos diferente heterotrimérico dirixido ao extremo máis (+), formado por dúas subunidades motoras relacionadas coa KHC e unha subunidade accesoria "KAP", procedente de extractos de embrións e ovos de equinodermos,[3] que é máis coñecido polo seu papel no transporte de complexos proteicos (partículas IFT) ao longo dos axonemas durante a bioxénese dos cilios.[4] Os estudos de xenética molecular e xenómica atoparon que as cinesinas forman unha superfamilia moi diversa de proteínas motoras que son responsables de moitos fenómenos de motilidade intracelular nas células eucarióticas.[5][6][7][8] Por exemplo, os xenomas de mamíferos codifican máis de 40 cinesinas distintas,[9] organizadas en polo menos 14 familias denominadas desde a cinesina-1 á cinesina-14.[10]
Estrutura
[editar | editar a fonte]Os membros da superfamilia da cinesina varían en forma pero a prototípica cinesina-1 é un heterotetrámero cuxas subunidades motoras (cadeas pesadas ou KHCs) forman unha proteína dímera que se une a dúas cadeas lixeiras (KLCs).
A cadea pesada da cinesina-1 consta dunha cabeza globular (o dominio motor) no extremo amino terminal conectada por medio dun curto colo de unión flexible co talo, que é un longo dominio superenrolado central en hélice alfa, que remata no dominio da cola carboxilo terminal, que se asocia coas cadeas lixeiras. Os talos de dúas KHCs entrelázanse para formar un superenrolamento que dirixe a dimerización das dúas KHCs. Na maioría dos casos o cargamento transportado únese ás cadeas lixeiras da cinesina polo motivo de secuencia TPR da KLC, pero nalgúns casos o cargamento únese ao dominio C-terminal das cadeas pesadas.[11]
Dominio do motor cinesina
[editar | editar a fonte]| Cinesina | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 | |||||||||
| Identificadores | |||||||||
| Símbolo | Kinesin motor domain | ||||||||
| Pfam | PF00225 | ||||||||
| InterPro | IPR001752 | ||||||||
| SMART | SM00129 | ||||||||
| PROSITE | PS50067 | ||||||||
| SCOPe | 1bg2 / SUPFAM | ||||||||
| |||||||||
A cabeza é o máis característico das cinesinas e a súa secuencia de aminoácidos está ben conservada entre as diversas cinesinas. Cada cabeza ten dous sitios de unión separados: un para o microtúbulo e o outro para o ATP. A unión do ATP e a súa hidrólise e liberación de ADP, fan cambiar a conformación tridimensional dos dominios de unión aos microtúbulos e a orientación do colo de unión con respecto á cabeza; isto ten como resultado o movemento da cinesina. Varios elementos estruturais da cabeza, como un dominio central en folla beta e os dominios Switch I e II, crese que están implicados na mediación das interaccións entre os dous sitios de unión e o dominio do colo. As cinesinas están relacionadas estruturalmente coas proteínas G, as cales hidrolizan o GTP en vez da ATP. As dúas familias comparten varios elementos estruturais, especialmente os dominios Switch I e II.
Transporte do cargamento
[editar | editar a fonte]No interior da célula as pequenas moléculas como os gases e a glicosa difunden facilmente a todas partes, pero moitas moléculas grandes sintetizadas na célula e compoñentes intracelulares como vesículas e orgánulos como as mitocondrias son demasiado grandes para difundir ata os seus destinos nun citosol que está ateigado de moléculas e compoñentes. As proteínas motoras desempeñan o papel de transportadoras de cargamentos no interior da célula, que levan aos destinos requiridos. As cinesinas son unha destas proteínas que transportan cargamentos "camiñando" unidireccionalmente ao longo dos microtúbulos hidrolizando unha molécula de ATP en cada paso que dan.[13] Críase que a enerxía do ATP impulsaba a cabeza da cinesina cara a adiante ata o seguinte punto de unión no microtúbulo.[14] Porén, propúxose tamén que a cabeza difunde cara a adiante e o que impulsa o cargamento cara a adiante é a forza de unión ao microtúbulo.[15]
Hai evidencias significativas de que os cargamentos son transportados in vivo por diversas proteínas motoras que actúan cooperativamente.[16][17][18][19]
Dirección do movemento
[editar | editar a fonte]As proteínas motoras viaxan nunha dirección específica ao longo dos microtúbulos. Isto débese a que o microtúbulo é polar e as cabezas só se unen ao microtúbulo nunha orientación, mentres que a unión do ATP dálle unha dirección a cada paso que se avanza por medio dun proceso chamado unión en cremalleira do colo de unión (neck linker zippering).[20]
A maioría das cinesinas camiñan cara ao extremo máis (+) do microtúbulo, o cal implica un transporte desde o centro á periferia da célula. Esta forma de transporte coñécese como transporte anterógrado ou ortógrado. Son excepcións as proteínas da familia cinesina-14, como a NCD de Drosophila melanogaster, a KAR3 dos lévedos de xemación, e a ATK5 da planta Arabidopsis thaliana, que camiñan na dirección oposta, cara ao extremo menos (-).[21]
Outro tipo distinto de proteínas motoras, as dineínas, móvense en dirección ao extremo menos, polo que fan un transporte retrógrado (da periferia ao centro).
Mecanismos de movemento propostos
[editar | editar a fonte]As cinesinas realizan o seu transporte utilizando como camiño un microtúbulo, e para explicar este movemento propuxéronse dous mecanismos:
- No mecanismo "man sobre man" as cabezas de cinesina dan un paso superando á outra cabeza, que queda máis atrás, de modo que se van alternado na posición máis avanzada.
- No mecanismo "eiruga", sempre lidera a mesma cabeza de cinesina, que se move un paso cara a adiante antes de que a outra cabeza a alcance (parecido a como se moven as eirugas).
A pesar que non se despexaron todas as controversias entre estes dous modelos, hai cada vez maiores probas experimentais que apuntan a que o mecanismo máis probable é o de "man sobre man".[22][23]
A unión e hidrólise do ATP fai que a cinesina se mova por un "mecanismo de balanceo" sobre un punto pivote.[24][25] Este mecanismo explica as observacións de que a unión do ATP ao estado unido ao microtúbulo sen nucleótido dá lugar á inclinación do domonio motor cinesina en relación co microtúbulo. Antes desta inclinación o colo de unión non pode adoptar a súa conformación orientada cara a adiante unida á cabeza do motor. A inclinación inducida polo ATP proporciona a oportunidade de que o colo de unión se una nesta conformación orientada cara a adiante. Este modelo está baseado en modelos de microscopia crioelectrónica (Cryo-EM) da estrutura da cinesina unida ao microtúbulo, o cal representa os estados inicial e final do proceso, pero non pode resolver os detalles precisos da transición entre as dúas estruturas.
Modelos teóricos da cinesina
[editar | editar a fonte]Propuxéronse varios modelos teóricos para a proteína motor molecular cinesina.[26][27][28] Os investigadores atoparon moitas dificultades teóricas polas incertezas que aínda hai sobre o papel das estruturas proteicas, o modo preciso en que a enerxía do ATP se transforma en traballo mecánico, e o pepel exercido polas flutuacións térmicas. Esta é unha área bastante activa de investigación. Necesítase unha maior investigación especialmente para saber a mellor forma de ligar os datos obtidos das investigacións experimentais coa arquitectura molecular da proteína.
Cinesina e mitose
[editar | editar a fonte]Nos últimos anos, encontrouse que os motores moleculares baseados en microtúbulos (como moitas cinesinas) exercen unha función na mitose. As cinesinas son importantes para que o fuso mitótico teña a lonxitude axeitada e están implicadas no escorregamento de microtúbulos no fuso durante a profase e metafase mitóticas, e a despolimerización dos extremos menos (-) dos microtúbulos durante a anafase.[29] Especificamente, a familia de proteínas da cinesina-5 actúa dentro do fuso para facer escorregar os microtúbulos, mentres que a familia cinesina-13 actúa despolimerizando os microtúbulos.
Membros da superfamilias das cinesinas
[editar | editar a fonte]A superfamilia das cinesinas humanas inclúe as seguintes proteínas, que na nomenclatura estandarizada desenvolvida polos investigadores que traballan no campo das cinesinas se organizan en 14 familias, que van da cinesina-1 á cinesina-14, e que son:[10]
- 1A – KIF1A, 1B – KIF1B = cinesina-3
- 2A – KIF2A, 2C – KIF2C = cinesina-13
- 3B – KIF3B, 3C – KIF3C = cinesina-2
- 4A – KIF4A, 4B – KIF4B = cinesina-4
- 5A – KIF5A, 5B – KIF5B, 5C – KIF5C = cinesina-1
- 6 – KIF6 = cinesina-9
- 7 – KIF7 = cinesina-4
- 9 – KIF9 = cinesina-9
- 11 – KIF11 = cinesina-5
- 12 – KIF12 = cinesina-12
- 13A – KIF13A, 13B – KIF13B = cinesina-3
- 14 – KIF14 = cinesina-3
- 15 – KIF15 = cinesina-12
- 16B – KIF16B = cinesina-3
- 17 – KIF17 = cinesina-2
- 18A – KIF18A, 18B – KIF18B = cinesina-8
- 19 – KIF19 = cinesina-8
- 20A – KIF20A, 20B – KIF20B = cinesina-6
- 21A – KIF21A, 21B – KIF21B = cinesina-4
- 22 – KIF22 = cinesina-10
- 23 – KIF23 = cinesina-6
- 24 – KIF24 = cinesina-13
- 25 – KIF25 = cinesina-14
- 26A – KIF26A, 26B – KIF26B = cinesina-11
- 27 – KIF27 = cinesina-4
- C1 – KIFC1, C2 – KIFC2, C3 – KIFC3 = cinesina-14
Cadeas lixeiras da cinesina-1:
Proteína asociada á cinesina-2:
- KAP-1, KAP3 ou KIFAP3
Notas
[editar | editar a fonte]- ↑ Vale RD (2003). "The molecular motor toolbox for intracellular transport". Cell 112 (4): 467–80. PMID 12600311. doi:10.1016/S0092-8674(03)00111-9.
- ↑ Vale RD, Reese TS, Sheetz MP (1985). "Identification of a novel force-generating protein, kinesin, involved in microtubule-based motility". Cell 42 (1): 39–50. PMC 2851632. PMID 3926325. doi:10.1016/S0092-8674(85)80099-4.
- ↑ Cole DG, Chinn SW, Wedaman KP, Hall K, Vuong T, Scholey JM (1993). "Novel heterotrimeric kinesin-related protein purified from sea urchin eggs". Nature 366 (6452): 268–70. Bibcode:1993Natur.366..268C. PMID 8232586. doi:10.1038/366268a0.
- ↑ Rosenbaum JL, Witman GB (2002). "Intraflagellar transport". Nat. Rev. Mol. Cell Biol. 3 (11): 813–25. PMID 12415299. doi:10.1038/nrm952.
- ↑ Yang JT, Laymon RA, Goldstein LS (1989). "A three-domain structure of kinesin heavy chain revealed by DNA sequence and microtubule binding analyses". Cell 56 (5): 879–89. PMID 2522352. doi:10.1016/0092-8674(89)90692-2.
- ↑ Aizawa H, Sekine Y, Takemura R, Zhang Z, Nangaku M, Hirokawa N (1992). "Kinesin family in murine central nervous system". J. Cell Biol. 119 (5): 1287–96. PMC 2289715. PMID 1447303. doi:10.1083/jcb.119.5.1287.
- ↑ Enos AP, Morris NR (1990). "Mutation of a gene that encodes a kinesin-like protein blocks nuclear division in A. nidulans". Cell 60 (6): 1019–27. PMID 2138511. doi:10.1016/0092-8674(90)90350-N.
- ↑ Meluh PB, Rose MD (1990). "KAR3, a kinesin-related gene required for yeast nuclear fusion". Cell 60 (6): 1029–41. PMID 2138512. doi:10.1016/0092-8674(90)90351-E.
- ↑ Hirokawa N, Noda Y, Tanaka Y, Niwa S (2009). "Kinesin superfamily motor proteins and intracellular transport". Nat. Rev. Mol. Cell Biol. 10 (10): 682–96. PMID 19773780. doi:10.1038/nrm2774.
- ↑ 10,0 10,1 Lawrence CJ, Dawe RK, Christie KR, Cleveland DW, Dawson SC, Endow SA, Goldstein LS, Goodson HV, Hirokawa N, Howard J, Malmberg RL, McIntosh JR, Miki H, Mitchison TJ, Okada Y, Reddy AS, Saxton WM, Schliwa M, Scholey JM, Vale RD, Walczak CE, Wordeman L (2004). "A standardized kinesin nomenclature". J. Cell Biol. 167 (1): 19–22. PMC 2041940. PMID 15479732. doi:10.1083/jcb.200408113.
- ↑ Hirokawa N, Pfister KK, Yorifuji H, Wagner MC, Brady ST, Bloom GS (1989). "Submolecular domains of bovine brain kinesin identified by electron microscopy and monoclonal antibody decoration". Cell 56 (5): 867–78. PMID 2522351. doi:10.1016/0092-8674(89)90691-0.
- ↑ PDB - 1BG2; Kull FJ, Sablin EP, Lau R, Fletterick RJ, Vale RD (1996). "Crystal structure of the kinesin motor domain reveals a structural similarity to myosin". Nature 380 (6574): 550–5. Bibcode:1996Natur.380..550J. PMC 2851642. PMID 8606779. doi:10.1038/380550a0.
- ↑ Schnitzer MJ, Block SM (1997). "Kinesin hydrolyses one ATP per 8-nm step". Nature 388 (6640): 386–390. Bibcode:1997Natur.388..386S. PMID 9237757. doi:10.1038/41111.
- ↑ Vale RD, Milligan RA (2000). "The way things move: looking under the hood of molecular motor proteins". Science 288 (5463): 88–95. Bibcode:2000Sci...288...88V. PMID 10753125. doi:10.1126/science.288.5463.88.
- ↑ Mather WH, Fox RF (2006). "Kinesin's biased stepping mechanism: amplification of neck linker zippering". Biophys. J. 91 (7): 2416–26. Bibcode:2006BpJ....91.2416M. PMC 1562392. PMID 16844749. doi:10.1529/biophysj.106.087049.
- ↑ Gross SP, Vershinin M, Shubeita GT (2007). "Cargo transport: two motors are sometimes better than one". Current Biology : CB 17 (12): R478–86. PMID 17580082. doi:10.1016/j.cub.2007.04.025.
- ↑ Hancock WO (2008). "Intracellular transport: kinesins working together". Current Biology : CB 18 (16): R715–7. PMID 18727910. doi:10.1016/j.cub.2008.07.068.
- ↑ Kunwar A, Vershinin M, Xu J, Gross SP (2008). "Stepping, strain gating, and an unexpected force-velocity curve for multiple-motor-based transport". Current Biology : CB 18 (16): 1173–83. PMID 18701289. doi:10.1016/j.cub.2008.07.027.
- ↑ Klumpp S, Lipowsky R (2005). "Cooperative cargo transport by several molecular motors". Proceedings of the National Academy of Sciences of the United States of America 102 (48): 17284–9. Bibcode:2005PNAS..10217284K. PMC 1283533. PMID 16287974. arXiv:q-bio/0512011. doi:10.1073/pnas.0507363102.
- ↑ Rice S, Lin AW, Safer D, Hart CL, Naber N, Carragher BO, Cain SM, Pechatnikova E, Wilson-Kubalek EM, Whittaker M, Pate E, Cooke R, Taylor EW, Milligan RA, Vale RD (1999). "A structural change in the kinesin motor protein that drives motility". Nature 402 (6763): 778–84. Bibcode:1999Natur.402..778R. PMID 10617199. doi:10.1038/45483.
- ↑ Ambrose JC, Li W, Marcus A, Ma H, Cyr R (2005). "A minus-end-directed kinesin with plus-end tracking protein activity is involved in spindle morphogenesis". Mol. Biol. Cell 16 (4): 1584–92. PMC 1073643. PMID 15659646. doi:10.1091/mbc.E04-10-0935.
- ↑ Yildiz A, Tomishige M, Vale RD, Selvin PR (2004). "Kinesin Walks Hand-Over-Hand". Science 303 (5658): 676–8. Bibcode:2004Sci...303..676Y. PMID 14684828. doi:10.1126/science.1093753.
- ↑ Asbury CL (2005). "Kinesin: world’s tiniest biped". Current Opinion in Cell Biology 17 (1): 89–97. PMID 15661524. doi:10.1016/j.ceb.2004.12.002.
- ↑ Sindelar CV, Downing KH (2010). "An atomic-level mechanism for activation of the kinesin molecular motors". Proc Natl Acad Sci U S A 107 (9): 4111–6. Bibcode:2010PNAS..107.4111S. PMC 2840164. PMID 20160108. doi:10.1073/pnas.0911208107.
- ↑ Lay Summary (18 February 2010). "Life’s smallest motor, cargo carrier of the cells, moves like a seesaw". PhysOrg.com. Consultado o 31 May 2013.
- ↑ Atzberger PJ, Peskin CS (2006). "A Brownian Dynamics model of kinesin in three dimensions incorporating the force-extension profile of the coiled-coil cargo tether". Bull. Math. Biol. 68 (1): 131–60. PMID 16794924. doi:10.1007/s11538-005-9003-6.
- ↑ Peskin CS, Oster G (1995). "Coordinated hydrolysis explains the mechanical behavior of kinesin". Biophys. J. 68 (4 Suppl): 202S–210S; discussion 210S–211S. PMC 1281917. PMID 7787069.
- ↑ Mogilner A, Fisher AJ, Baskin RJ (2001). "Structural changes in the neck linker of kinesin explain the load dependence of the motor's mechanical cycle". J. Theor. Biol. 211 (2): 143–57. PMID 11419956. doi:10.1006/jtbi.2001.2336.
- ↑ Goshima G, Vale RD (2005). "Cell cycle-dependent dynamics and regulation of mitotic kinesins in Drosophila S2 cells". Mol. Biol. Cell 16 (8): 3896–907. PMC 1182325. PMID 15958489. doi:10.1091/mbc.E05-02-0118.
Véxase tamén
[editar | editar a fonte]Outros artigos
[editar | editar a fonte]Bibliografía
[editar | editar a fonte]- Lawrence CJ, Dawe RK, Christie KR, Cleveland DW, Dawson SC, Endow SA, Goldstein LS, Goodson HV, Hirokawa N, Howard J, Malmberg RL, McIntosh JR, Miki H, Mitchison TJ, Okada Y, Reddy AS, Saxton WM, Schliwa M, Scholey JM, Vale RD, Walczak CE, Wordeman L (2004). "A standardized kinesin nomenclature". J. Cell Biol. 167 (1): 19–22. PMC 2041940. PMID 15479732. doi:10.1083/jcb.200408113.
Ligazóns externas
[editar | editar a fonte]- Animated model of kinesin walking Arquivado 15 de marzo de 2024 en Wayback Machine.
- Ron Vale's seminar:"Cytoskeletal Motor Proteins"
- Kinesin and Dynein Microtubule Movement
- The Inner Life of a Cell, 3D animation featuring a Kinesin transporting a vesicle Arquivado 07 de decembro de 2008 en Wayback Machine.
- The Kinesin Homepage
- MeshName Kinesin
- Número EC 3.6.4.4 ExPASY
- Número EC 3.6.4.5 ExPASY
- 3D electron microscopy structures of kinesin from the EM Data Bank(EMDB)