Cellule (biologie)
Cet article concerne l'unité de base du vivant. Pour la subdivision d'aile d'insectes, voir Cellule (entomologie). Pour les autres significations, voir Cellule.


a. cellules qui ne se divisent pas, avec réseau de chromatine et nucléoles fortement colorés ;
b. noyaux se préparant à la division ;
c. Cellules en cours de division présentant des figures de mitose ;
e. Paire de cellules filles peu après la division. »


La cellule — du latin cellula « chambre de moine »[1] — est l'unité biologique structurelle et fonctionnelle fondamentale de tous les êtres vivants connus. C'est la plus petite unité vivante capable de se reproduire de façon autonome. La science qui étudie les cellules est appelée biologie cellulaire.
Une cellule est constituée d'une membrane plasmique contenant un cytoplasme, lequel est formé d'une solution aqueuse (cytosol) dans laquelle se trouvent de nombreuses biomolécules telles que des protéines et des acides nucléiques, organisées ou non dans le cadre d'organites. De nombreux êtres vivants ne sont constitués que d'une seule cellule : ce sont les organismes unicellulaires, comme les bactéries, les archées et la plupart des protistes. D'autres sont constitués de plusieurs cellules : ce sont les organismes multicellulaires, comme les plantes et les animaux. Ces derniers contiennent un nombre de cellules très variable d'une espèce à l'autre ; le corps humain en compte ainsi de l'ordre de cent mille milliards (1014), mais est colonisé par un nombre de un[3] à dix[4] fois plus grand de bactéries, qui font partie de son microbiote et sont bien plus petites que les cellules humaines. La plupart des cellules des plantes et des animaux ne sont visibles qu'au microscope, avec un diamètre compris entre 10 et 100 µm.
L'existence des cellules a été découverte en 1665 par le naturaliste anglais Robert Hooke. La théorie cellulaire a été formulée pour la première fois en 1839 par le botaniste allemand Matthias Jakob Schleiden et l'histologiste allemand Theodor Schwann : elle expose que tous les êtres vivants sont constitués d'une ou plusieurs cellules, que les cellules sont les unités fondamentales de toutes les structures biologiques, qu'elles dérivent toujours d'autres cellules préexistantes, et qu'elles contiennent l'information génétique nécessaire à leur fonctionnement ainsi qu'à la transmission de l'hérédité aux générations de cellules suivantes. Les premières cellules sont apparues sur Terre il y a au moins 3,7 milliards d'années[5], et peut-être dès 4 Ga[6].
Étymologie
[modifier | modifier le code]Le nom de « cellule » est dû à son découvreur Robert Hooke, qui leur a donné le nom latin cellula en référence aux petites chambres occupées par les moines dans les monastères. Cellula est dérivé de cella, qui désignait en latin une pièce, ou un cellier — qui vient de son dérivé cellarium (« garde-manger »).
Cella est issu de l’indo-européen commun *k̂el (« couvrir ») dont proviennent aussi, indirectement, le français celer (« cacher ») ou l'anglais hell (« monde souterrain, enfer »)...
Principales structures cellulaires
[modifier | modifier le code]On considère généralement deux types fondamentaux de cellules selon qu'elles possèdent ou non un noyau enveloppé d'une membrane nucléaire :
- les procaryotes, dont l'ADN est libre dans le cytoplasme, comprenant les bactéries et les archées, et qui, bien qu'ils puissent former des colonies très nombreuses, sont toujours des organismes unicellulaires ;
- les eucaryotes, qui ont une organisation interne complexe, de nombreux organites, et dont le noyau est délimité par une membrane nucléaire. Les eucaryotes comprennent un grand nombre de formes d'organismes unicellulaires et organismes multicellulaires.
| Procaryotes | Eucaryotes | |
|---|---|---|
| Représentants | Bactéries, archées | Protistes, mycètes, plantes, animaux |
| Taille typique | ~ 1 à 5 μm | ~ 10 à 100 μm |
| Type de noyau | Nucléoïde ; pas de véritable noyau | Vrai noyau avec membrane nucléaire |
| ADN | Généralement circulaire | Molécules linéaires (chromosomes) avec histones |
| Transcription/Traduction génétique | Biosynthèse des protéines intégralement dans le cytoplasme | Transcription et traduction séparées spatialement :
|
| Ribosomes | Grande et petite sous-unités : | Grande et petite sous-unités : |
| Compartiments cellulaires | Peu de structures intracellulaires | Nombreuses structures : système endomembranaire, cytosquelette |
| Motilité cellulaire | Flagelle constitué de flagelline | Flagelle et cils constitués de tubuline |
| Métabolisme | anaérobie ou aérobie selon les cas | Généralement aérobie |
| Mitochondries | Aucune | D'aucune à plusieurs milliers |
| Chloroplastes | Aucun | Dans les algues et les plantes chlorophylliennes |
| Organisation unicellulaire ou multicellulaire |
Cellules généralement isolées (unicellulaires) | Cellules isolées, colonies, organismes complexes avec des cellules spécialisées (multicellulaires) |
| Division cellulaire | Scissiparité (division simple) | Mitose (multiplication conforme de la cellule) Méiose (formation de gamètes) |
| Matériel génétique | Chromosome unique et plasmides | Chromosomes multiples |
Procaryotes
[modifier | modifier le code]Les procaryotes sont la première forme de vie apparue sur Terre, définie comme étant autosuffisante et pourvue de tous les processus biologiques vitaux, y compris les mécanismes de signalisation cellulaire. Plus petites et plus simples que les cellules d'eucaryotes, les cellules de procaryotes sont dépourvues de système endomembranaire et des organites qui le constituent, à commencer par le noyau. Les bactéries et les archées sont les deux domaines du vivant regroupant les procaryotes. L'ADN d'un procaryote forme un unique chromosome en contact direct avec le cytoplasme. La région nucléaire du cytoplasme est appelée nucléoïde et n'est pas nettement séparée du reste de la cellule. La plupart des procaryotes sont les plus petits des êtres vivants connus, avec un diamètre compris entre 0,5 et 2 µm.
Structure d'un procaryote typique, montrant les éléments suivants :
|

Une cellule de procaryote contient trois régions distinctes :
- l'extérieur de la cellule porte des flagelles et des pili qui sortent de la surface cellulaire chez un grand nombre de procaryotes. Ces structures permettent à ces cellules de se déplacer (motilité cellulaire) et de communiquer entre elles ;
- la surface de la cellule est délimitée par l'enveloppe cellulaire (en), formée généralement d'une paroi cellulaire recouvrant une membrane plasmique, certaines bactéries étant recouvertes d'une couche supplémentaire appelée capsule. L'enveloppe confère sa rigidité à la cellule et sépare l'intérieur de la cellule de l'environnement de cette dernière en jouant le rôle d'un filtre protecteur. La plupart des procaryotes sont dotés d'une paroi cellulaire, mais il existe des exceptions telles que les genres Mycoplasma (une bactérie) et Thermoplasma (une archée). La paroi cellulaire des bactéries est constituée de peptidoglycane et agit comme barrière supplémentaire contre les agents extérieurs. Elle empêche également la cellule de se dilater et d'éclater (cytolyse) sous l'effet de sa pression osmotique interne dans un environnement hypotonique. Certains eucaryotes sont également pourvus d'une paroi cellulaire, par exemple les cellules végétales et les cellules fongiques ;
- l'intérieur de la cellule possède une région qui contient le génome (ADN) et contient également des ribosomes et divers types d'inclusions. Les ribosomes sont responsables de l'aspect granulaire du cytoplasme des procaryotes. Le matériel génétique est libre dans le cytoplasme. Les procaryotes peuvent porter des éléments extrachromosomiques appelés plasmides, qui sont généralement circulaires. Ces plasmides encodent des gènes supplémentaires, comme des gènes de résistance aux antibiotiques. Des plasmides bactériens linéaires ont été identifiés chez plusieurs espèces de spirochètes, notamment le genre Borrelia, et particulièrement Borrelia burgdorferi, responsable de la maladie de Lyme. À la place du noyau des eucaryotes, les procaryotes possèdent une région nucléaire diffuse appelée nucléoïde.
Eucaryotes
[modifier | modifier le code]
Cellule animale :
|

Structure schématique d'une cellule végétale. On y observe notamment les éléments suivants :
|

Les plantes, les animaux, les mycètes, les protozoaires et les algues sont des eucaryotes. Ces cellules sont en moyenne 15 fois plus grandes qu'un procaryote typique, et peuvent être jusqu'à mille fois plus volumineuses. La principale caractéristique qui distingue les eucaryotes des procaryotes est leur compartimentation en organites spécialisés au sein desquels se déroulent des processus métaboliques spécifiques. Parmi ces organites se trouve le noyau, qui héberge l'ADN de la cellule. C'est la présence de ce noyau qui donne son nom à ce type de cellules, eucaryote étant forgé à partir de racines grecques signifiant « à vrai noyau ». Par ailleurs :
- la fonction biologique de la membrane plasmique des eucaryotes est semblable à celle des procaryotes, mais avec une structure qui peut être un peu différente. Il existe des eucaryotes avec et sans paroi cellulaire ; dans la cellule végétale, il s'agit d'une paroi pectocellulosique constituée de polysaccharides, notamment de cellulose, d'hémicellulose et de pectine ;
- le matériel génétique des eucaryotes est organisé en une ou plusieurs molécules d'ADN linéaires associées à des histones pour former des chromosomes. Tout l'ADN chromosomique d'un eucaryote est stocké dans le noyau, séparé du cytoplasme par la membrane nucléaire. Certains organites des eucaryotes contiennent également de l'ADN, par exemple les mitochondries ;
- de nombreuses cellules d'eucaryotes sont pourvues de cils primaires, qui jouent un rôle important dans la perception par la cellule d'informations chimiques, mécaniques et thermiques sur son environnement. Les cils primaires doivent donc être vus comme des antennes cellulaires sensibles qui coordonnent un grand nombre de voies métaboliques de signalisation cellulaire, parfois en couplant la signalisation à la motilité ou à la division cellulaire, voire à la différenciation cellulaire[7] ;
- certaines cellules d'eucaryotes peuvent se déplacer par motilité au moyen de cils vibratiles ou de flagelles. Ces derniers forment des systèmes plus simples chez les eucaryotes que chez les procaryotes.
Éléments constitutifs d'une cellule
[modifier | modifier le code]
Toutes les cellules, qu'il s'agisse de procaryotes ou d'eucaryotes, possèdent une membrane plasmique qui les enveloppe, régule les flux de matière entrants et sortants (transport membranaire) et maintient un potentiel électrochimique de membrane. Contenu dans cette membrane se trouve le cytoplasme, qui est une solution aqueuse riche en sels dissous occupant l'essentiel du volume de la cellule. Toutes les cellules possèdent un matériel génétique constitué d'ADN, ainsi que de l'ARN qui intervient essentiellement dans la biosynthèse des protéines et des enzymes, ces dernières étant responsables du métabolisme de la cellule ; les érythrocytes (globules rouges du sang) font exception, car leur cytoplasme est dépourvu de presque tous les organites constituant normalement une cellule d'eucaryote, ce qui leur permet d'accroître la quantité d'hémoglobine qu'ils peuvent contenir, et ne possèdent donc pas de noyau, dans lequel se trouverait l'ADN. Il existe une très grande variété de biomolécules dans les cellules.
Membrane
[modifier | modifier le code]
En haut : membrane archéenne ; 1 chaîne latérale terpénoïde ; 2 liaison éther ; 3 L-glycérol ; 4 groupe phosphate. Au milieu : membrane archéenne et bactérienne ; 5 acide gras ; 6 liaison ester ; 7 D-glycérol ; 8 groupe phosphate. En bas : 9 bicouche lipidique chez les eucaryotes, les bactéries et la plupart des archées ; 10 monocouche lipidique chez certaines archées.
La membrane plasmique, ou membrane cellulaire, est une membrane biologique qui entoure et délimite le cytoplasme d'une cellule. Chez les animaux, la membrane matérialise la surface de la cellule, tandis que, chez les plantes et les procaryotes, elle est généralement recouverte d'une paroi cellulaire. Ainsi, chez les plantes, les algues et les mycètes, la cellule est incluse dans une paroi pectocellulosique, qui fournit un squelette à l'organisme[8]. Des dépôts de composés tels que la subérine ou la lignine modulent les propriétés physico-chimiques de la paroi, la rendant plus rigide ou plus imperméable, par exemple.
La membrane a pour fonction de séparer le milieu intracellulaire de l'environnement de la cellule en le protégeant de ce dernier. Elle est constituée d'une bicouche lipidique chez les eucaryotes, les bactéries et la plupart des archées, ou d'une monocouche d'étherlipides chez certaines archées. Chez les eucaryotes, il s'agit essentiellement de phospholipides, qui ont la propriété d'être amphiphiles, c'est-à-dire de posséder une tête polaire hydrophile et des queues aliphatiques hydrophobes. Une très grande variété de protéines, dites protéines membranaires, sont incluses dans la membrane plasmique, où elles jouent le rôle de canaux et de pompes assurant le transport membranaire entrant et sortant de la cellule. On dit que la membrane plasmique est semiperméable car sa perméabilité est très variable en fonction de l'espèce chimique considérée : certaines peuvent la traverser librement, d'autres ne peuvent la traverser que de façon limitée ou dans un seul sens, d'autres enfin ne peuvent pas la traverser du tout. La surface cellulaire contient également des récepteurs membranaires qui assurent la transduction de signal dans le cadre de mécanismes de signalisation cellulaire, ce qui permet à la cellule de réagir par exemple à la présence d'hormones.
Cytosquelette
[modifier | modifier le code]
Le cytosquelette intervient pour définir et maintenir la forme de la cellule (tenségrité), positionner les organites dans le cytoplasme, réaliser l'endocytose d'éléments extracellulaires, assurer la cytokinèse lors de la division cellulaire, et déplacer certaines régions du cytoplasme lors de la croissance et de la mobilité cellulaires (transport intracellulaire). Le cytosquelette des eucaryotes est composé de microfilaments, de filaments intermédiaires et de microtubules. Un grand nombre de protéines sont associées à ces structures, chacune d'entre elles contrôlant la structure de la cellule en orientant, liant et alignant les filaments. Le cytosquelette des procaryotes est moins connu mais intervient pour maintenir la forme et la polarité ainsi que pour assurer la cytokinèse de ces cellules[9]. La protéine constitutive des microfilaments est une petite protéine monomérique appelée actine, tandis que celle constitutive des microtubules est une protéine dimérique appelée tubuline. Les filaments intermédiaires sont des hétéropolymères dont les monomères varient en fonction du type de cellule et du tissu ; ce sont notamment la vimentine, la desmine, les lamines A, B et C, les kératines et les protéines des neurofilaments (NF-L et NF-M).
Matériel génétique
[modifier | modifier le code]
Le matériel génétique des cellules peut se trouver sous la forme d'ADN ou d'ARN (cellule sans noyau). C'est la séquence nucléotidique de l'ADN qui porte toute l'information génétique (génotype) d'une cellule. Cet ADN est transcrit en ARN, un autre type d'acide nucléique, qui assure diverses fonctions : transport de l'information génétique de l'ADN vers les ribosomes sous forme d'ARN messager, et traduction de l'ARN messager en protéines sous forme à la fois d'ARN de transfert et d'ARN ribosomique, ce dernier agissant comme un ribozyme.
Le matériel génétique des procaryotes est généralement constitué d'une molécule d'ADN circulaire unique formant un chromosome dans une région diffuse du cytoplasme appelée nucléoïde. Celui des eucaryotes est réparti sur plusieurs molécules d'ADN linéaires formant des chromosomes contenus dans un noyau cellulaire différencié. Les cellules d'eucaryotes contiennent également de l'ADN dans certains organites tels que les mitochondries et, chez les plantes, les chloroplastes.
Une cellule humaine contient de ce fait de l'ADN dans son noyau et dans ses mitochondries. On parle respectivement de génome nucléaire et de génome mitochondrial. Le génome nucléaire humain est réparti sur 46 molécules d'ADN linéaires formant autant de chromosomes. Ceux-ci sont organisés par paires, en l'occurrence 22 paires de chromosomes homologues et une paire de chromosomes sexuels. Le génome mitochondrial humain est contenu sur un chromosome circulaire et possède 38 gènes : 14 gènes encodent des sous-unités constituant cinq protéines (NADH déshydrogénase, cytochrome b, cytochrome c oxydase, ATP synthase et humanine), deux gènes encodent des ARN ribosomiques mitochondriaux (ARNr 12S et ARNr 16S), et 22 gènes encodent vingt ARN de transfert mitochondriaux.
Du matériel génétique exogène peut également être introduit dans une cellule par transfection. Ceci peut être permanent si l'ADN exogène est inséré de façon stable dans le génome de la cellule, ou transitoire dans le cas contraire. Certains virus insèrent également leur matériel génétique dans le génome de leur cellule hôte : c'est la transduction.
Organites
[modifier | modifier le code]
Les organites sont des compartiments cellulaires réalisant des fonctions biologiques spécialisées, de façon analogue aux organes du corps humain. Les cellules d'eucaryotes et de procaryotes possèdent des organites, mais ceux des procaryotes sont plus simples et ne sont généralement pas matérialisés par une membrane.
Il existe différents types d'organites dans une cellule. Certains sont généralement uniques, comme l'appareil de Golgi, tandis que d'autres sont présents en de très nombreux exemplaires — des centaines, voire des milliers — comme les mitochondries, les chloroplastes, les peroxysomes et les lysosomes. Le cytosol est le fluide gélatineux qui entoure les organites dans le cytoplasme.
Organites présents chez tous les êtres vivants
[modifier | modifier le code]- Ribosomes — Il s'agit de complexes moléculaires formés de protéines et d'ARN. Chaque ribosome est constitué de deux sous-unités et fonctionne comme une chaîne d'assemblage réalisant la biosynthèse des protéines à partir d'acides aminés. Les ribosomes peuvent se trouver ou bien en suspension dans le cytoplasme ou bien liés à la membrane plasmique chez les procaryotes ou au réticulum endoplasmique rugueux chez les eucaryotes[10].
Organites des cellules d'eucaryotes
[modifier | modifier le code]
Éléments du système endomembranaire :
|


- Noyau — C'est l'organite le plus visible des cellules d'eucaryotes et qui contient leur matériel génétique. C'est dans le noyau que se trouvent les chromosomes et que se déroulent la réplication de l'ADN ainsi que la transcription de l'ADN en ARN. Le noyau est de forme grossièrement sphérique et est séparé du cytoplasme par une double membrane appelée membrane nucléaire. Celle-ci a notamment pour fonction d'isoler et de protéger le matériel génétique cellulaire des agents chimiques susceptibles de l'endommager ou d'interférer avec ses fonctions biologiques. La biosynthèse des protéines commence dans le noyau par la transcription de l'ADN en ARN messager, lequel quitte ensuite le noyau en direction du cytoplasme, où des ribosomes assurent sa traduction en protéines. Les ribosomes sont assemblés dans une région particulière du noyau appelée nucléole avant de gagner le cytoplasme. Les procaryotes étant dépourvus de noyau, l'ADN et toutes ses fonctions biologiques prennent place directement dans le cytoplasme.
- Réticulum endoplasmique — Cet organite est une extension cytoplasmique de la membrane nucléaire. Il assure la biosynthèse et le transport de molécules marquées pour des transformations et des destinations spécifiques, contrairement aux molécules non marquées, qui dérivent librement dans le cytoplasme. Le réticulum endoplasmique rugueux est recouvert de ribosomes qui produisent des protéines essentiellement destinées à être sécrétées hors de la cellule où à demeurer dans la membrane plasmique ; il produit également des lipides destinés au système endomembranaire dans son ensemble, dont le réticulum endoplasmique fait partie. Le réticulum endoplasmique lisse est dépourvu de ribosomes à sa surface et intervient essentiellement dans la production de lipides, dans la détoxication de certains xénobiotiques ainsi que dans diverses fonctions sécrétrices selon les types de cellules.
- Appareil de Golgi — Cet organite est le lieu de transformation finale des protéines nouvellement synthétisées — modifications post-traductionnelles — essentiellement par glycosylation et phosphorylation.
- Vacuoles — Elles concentrent les déchets cellulaires et, chez les plantes, stockent l'eau. Elles sont souvent décrites comme des volumes remplis de liquide et délimités par une membrane. Certaines cellules, notamment les amibes du genre Amoeba, possèdent des vacuoles contractiles qui peuvent pomper l'eau hors de la cellule si celle-ci en contient trop. Les vacuoles des cellules d'eucaryotes sont généralement plus grandes chez les plantes que chez les animaux[11].
- Centrosome — Cet organite produit les microtubules de la cellule, qui sont un composant essentiel du cytosquelette. Il organise le transport à travers le réticulum endoplasmique et l'appareil de Golgi. Les centrosomes sont constitués chacun de deux centrioles, qui se séparent lors de la division cellulaire et contribuent la formation du fuseau mitotique. Les cellules animales possèdent un centrosome unique ; on en trouve également chez certaines algues et certains mycètes.
- Lysosomes et peroxysomes — Les lysosomes contiennent essentiellement des hydrolases acides, qui sont des enzymes digestives. Ces organites ont donc pour fonction de dégrader les éléments cellulaires endommagés ou inutilisés, ainsi que les particules alimentaires, les virus et les bactéries phagocytés. Les peroxysomes contiennent des enzymes qui éliminent les peroxydes nocifs pour la cellule. Une cellule ne pourrait contenir ce type d'enzymes destructrices si elles n'étaient pas contenues dans des organites délimités par un système de membranes.
- Mitochondries et chloroplastes — Ce sont les organites assurant la production d'énergie métabolique de la cellule. Les mitochondries sont des organites qui se reproduisent par réplication et sont présents dans le cytoplasme de toutes les cellules d'eucaryotes sous des formes, des tailles et en nombre très variables. C'est dans les mitochondries que se déroule la respiration cellulaire, produisant de l'énergie métabolique sous forme d'ATP et du pouvoir réducteur sous forme de NADH et de FADH2 à travers un ensemble de voies métaboliques — β-oxydation, cycle de Krebs, chaîne respiratoire, phosphorylation oxydative — qui convertissent en énergie les nutriments cellulaires tels que les acides gras et les oses. Les mitochondries se multiplient par scissiparité (division simple) comme les procaryotes. Les chloroplastes ne se trouvent que chez les plantes et les algues et assurent la production de glucides à partir du rayonnement solaire par photosynthèse.
Structures extracellulaires
[modifier | modifier le code]De nombreuses cellules possèdent également des structures entièrement ou partiellement situées à l'extérieur de la membrane plasmique. Ces structures ne sont donc pas protégées de l'environnement de la cellule par une membrane semiperméable. L'assemblage de ces structures implique que leurs constituants soient transportés hors de la cellule par des processus spécifiques.
Paroi cellulaire
[modifier | modifier le code]
De nombreux types de cellules de procaryotes et d'eucaryotes possèdent une paroi cellulaire. Celle-ci protège la cellule des actions chimiques et mécaniques de son environnement et ajoute une couche protectrice supplémentaire par-dessus la membrane plasmique. Les différents types de cellules tendent à produire des parois de nature chimique différente : la paroi pectocellulosique des plantes est constituée essentiellement de cellulose, la paroi des mycètes est constituée essentiellement de chitine, et la paroi bactérienne est constituée essentiellement de peptidoglycane.
Structures spécifiques aux procaryotes
[modifier | modifier le code]
- Capsule — Il s'agit d'une capsule gélatineuse présente chez certaines bactéries par-dessus la paroi bactérienne et la membrane plasmique. Elle peut être constituée de polysaccharides comme chez les pneumocoques et les méningocoques, de polypeptides comme chez le bacille du charbon, ou encore d'acide hyaluronique comme chez les streptocoques. Elles ne sont pas colorées par les procédés standard et peuvent être marquées à l'encre de Chine ou au bleu de méthyle.
- Flagelle — C'est l'organite essentiel de la motilité cellulaire. Le flagelle bactérien prend naissance dans le cytoplasme, traverse la membrane plasmique et les différentes parois qui peuvent éventuellement la recouvrir, et s'étend largement dans le milieu extracellulaire. Les flagelles sont de nature intégralement protéique. Ils sont de types différents chez les bactéries, les archées et les eucaryotes.
- Fimbriae — Il s'agit de l'appellation des pili en bactériologie. Elles se présentent comme des cils à la surface de la bactérie. Les fimbriae sont constituées d'une protéine appelée piline et interviennent dans les processus d'adhérence cellulaire. Il existe des types de pili spécifiques impliqués dans la conjugaison des bactéries.
Processus cellulaires
[modifier | modifier le code]Métabolisme et cycle cellulaire
[modifier | modifier le code]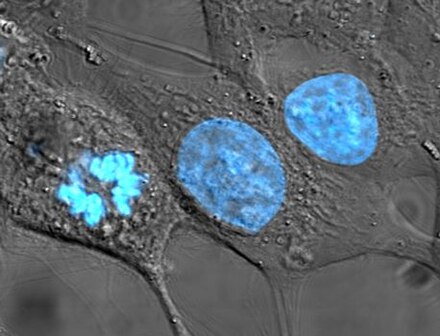
Entre deux divisions successives du cycle cellulaire, les cellules se développent grâce à leur métabolisme. Le métabolisme cellulaire est le processus par lequel chaque cellule exploite les nutriments qu'elle absorbe afin de se maintenir en vie et de se reproduire. Le métabolisme se divise en deux grandes parties : d'une part le catabolisme, par lequel les cellules dégradent les molécules complexes en espèces chimiques plus simples afin de produire de l'énergie métabolique sous forme par exemple d'ATP et du pouvoir réducteur sous forme par exemple de NADH et de FADH2 ; d'autre part l'anabolisme qui utilise l'énergie et le pouvoir réducteur produits par le catabolisme afin de synthétiser des biomolécules et de réaliser d'autres fonctions biologiques.
Le cycle cellulaire est l'ensemble des processus biologiques conduisant à la division d'une cellule mère en deux cellules filles. Chez les procaryotes, qui ne possèdent pas de noyau, la réplication des cellules se fait par scissiparité, c'est-à-dire par division simple. Chez les eucaryotes, en revanche, le cycle cellulaire est divisé en trois grandes phases : l'interphase, la mitose et la cytokinèse. Au cours de l'interphase, les cellules grossissent en accumulant les substances nécessaires à la préparation de la division cellulaire et à la réplication de l'ADN. Puis le noyau se scinde en deux au cours de la mitose, et enfin le cytoplasme achève de se scinder à son tour en deux, avec un noyau dans chacune des deux parties, au cours de la cytokinèse. Des mécanismes appelés points de contrôle (en) assurent que la division se déroule de façon conforme[12].
Réplication
[modifier | modifier le code]
La division cellulaire est le processus par lequel une cellule unique, dite cellule mère, donne naissance à deux cellules, dites cellules filles. Ceci permet la croissance des organismes multicellulaires et la multiplication des organismes unicellulaires. Les cellules de procaryotes se divisent par scissiparité (division simple) tandis que les cellules d'eucaryotes se divisent d'abord au niveau de leur noyau — phase de mitose — puis au niveau de l'ensemble du cytoplasme — phase de cytokinèse. Une cellule diploïde peut également donner des cellules haploïdes, généralement au nombre de quatre, à travers le processus de méiose ; les cellules haploïdes interviennent comme gamètes chez les organismes multicellulaires en fusionnant avec d'autres gamètes pour redonner des cellules diploïdes.
La réplication de l'ADN, qui est la base moléculaire de la réplication du génome d'une cellule, intervient toujours lorsqu'une cellule se divise par mitose ou par scissiparité ; elle a lieu à la phase S du cycle cellulaire. Au cours de la méiose, l'ADN n'est répliqué qu'une seule fois alors que la cellule se divise deux fois : la réplication de l'ADN intervient lors de la première division de la méiose, mais pas lors de la division subséquente. La réplication, comme tous les autres processus cellulaires, requiert des protéines et des enzymes spécialisées pour être menée à bien.
Dans le cas des organismes unicellulaires, il est généralement accepté que les cellules prolifèrent spontanément, sans avoir besoin de stimulation. Dans le cas des organismes multicellulaires, cette question fait l'objet d'un débat[14]. De nombreux auteurs défendent l'idée que ces cellules requièrent une stimulation pour proliférer, d'autres au contraire considèrent que la quiescence est le résultat de contraintes agissant sur les cellules quiescentes[15],[16]. Pour la modélisation du comportement des cellules, les deux points de vue sont couramment utilisés[17].
Biosynthèse des protéines
[modifier | modifier le code]
L'une des activités biochimiques principales des cellules est de produire de nouvelles protéines. Ces dernières sont essentielles à la régulation et à la maintenance de l'activité cellulaire. La synthèse des protéines se décompose en plusieurs étapes : transcription de l'ADN en ARN messager, modifications post-transcriptionnelles de l'ARN messager, traduction de l'ARN messager en protéines, modifications post-traductionnelles des protéines nouvellement synthétisées, et enfin repliement des protéines dans leur conformation fonctionnelle, appelée état natif.
Lors de la transcription, des ARN polymérases produisent un brin d'ARN complémentaire au brin d'ADN codant. L'information génétique est portée par la séquence nucléotidique de l'ADN, reproduite sur l'ARN messager lors de la transcription. Cette séquence est ensuite lue par des ribosomes afin de polymériser les acides aminés dans l'ordre spécifié par la succession de groupes de trois nucléotides sur l'ARN messager, chacun de ces triplets, appelés codons, correspondant à un acide aminé donné ; c'est cette correspondance entre codons et acides aminés qu'on appelle code génétique.
Locomotion ou motilité
[modifier | modifier le code]Les unicellulaires peuvent se déplacer afin de trouver de la nourriture ou d'échapper à leurs prédateurs. Flagelles et cils sont les principaux moyens de motilité cellulaire.
Chez les organismes multicellulaires, les cellules peuvent se déplacer par exemple lors de la cicatrisation des plaies, lors de la réponse immunitaire, ou encore lors de la formation de métastases tumorales. Ainsi, les leucocytes (globules blancs) se déplacent jusqu'à la plaie afin d'y tuer les micro-organismes susceptibles d'y provoquer des infections. La motilité cellulaire fait intervenir de nombreux récepteurs, des mécanismes de réticulation, d'assemblage, de liaison ou encore d'adhérence de protéines, ainsi que des protéines motrices, parmi d'autres types de protéines[18]. Le processus se déroule en trois temps : saillie de la pointe avant de la cellule, adhérence de l'avant de la cellule et « désadhérence » du reste de la surface cellulaire, et contraction du cytosquelette pour tirer la cellule vers l'avant. Chacune de ces étapes est gérée par des forces produites par des segments particuliers du cytosquelette[18]. Comme pour la prolifération, la question de savoir si la motilité des cellules d'un multicellulaire est spontanée, comme pour les unicellulaires, ou doit faire l'objet de stimulations fait l'objet d'un débat[15].
Pluricellularité
[modifier | modifier le code]Un organisme multicellulaire est constitué de plusieurs cellules, par opposition à un organisme unicellulaire.
Spécialisation cellulaire
[modifier | modifier le code]Au sein des organismes multicellulaires, les cellules se spécialisent en différents types cellulaires (en) adaptés chacun à des fonctions physiologiques particulières. Chez les mammifères par exemple, on trouve par exemple des cellules de la peau, des myocytes (cellules musculaires), des neurones (cellules nerveuses), des cellules sanguines, des fibroblastes (cellules des tissus conjonctifs), ou encore des cellules souches. Des cellules de types différents d'un même organisme ont une fonction physiologique et une apparence propres, mais partagent le même génome. Des cellules ayant le même génotype peuvent présenter des phénotypes différents en raison d'une expression génétique différenciée : les gènes qu'elles contiennent ne sont pas exprimés de la même façon les uns par rapport aux autres, certains le sont davantage dans un type cellulaire que dans un autre.
Tous les types cellulaires d'un organisme donné dérivent d'une cellule unique dite totipotente, c'est-à-dire capable de se différencier en n'importe quel type cellulaire lors du développement de l'organisme. La différenciation cellulaire est influencée par divers facteurs environnementaux (par exemple les interactions cellule-cellule (en)) et des différences intrinsèques (par exemple la distribution non uniforme des molécules lors de la division).
Origine de la multicellularité
[modifier | modifier le code]
La multicellularité a émergé à partir d'organismes unicellulaires un grand nombre de fois au cours de l'évolution et ne s'observe pas uniquement chez les eucaryotes : certains procaryotes tels que des cyanobactéries, des myxobactéries, actinomycètes, Magnetoglobus multicellularis ou encore des archées du genre Methanosarcina, présentent des organisations multicellulaires. Cependant, ce sont bien chez les eucaryotes que des organismes multicellulaires sont apparus, et ce parmi six groupes : les animaux, les mycètes, les algues brunes, les algues rouges, les algues vertes et les plantes. La multicellularité peut être apparue à partir de colonies d'organismes interdépendants, voire à partir d'organismes en symbiose.
Les plus anciennes traces de multicellularité ont été identifiées chez des organismes apparentés aux cyanobactéries qui vivaient il y a entre 3 et 3,5 milliards d'années. Parmi d'autres fossiles d'organismes multicellulaires, on compte également Grypania spiralis[19],[20], dont la nature biologique exacte demeure cependant débattue, ainsi que les fossiles des schistes paléoprotérozoïques du groupe fossile de Franceville[21], au Gabon.
L'évolution d'organismes multicellulaires à partir d'ancêtres unicellulaires a été reproduite au laboratoire à travers des expériences d'évolution expérimentale utilisant la prédation comme vecteur de pression de sélection.
Origine des cellules
[modifier | modifier le code]L'origine des cellules est intimement liée à l'origine de la vie, aux origines de l'histoire évolutive du vivant.
Origines des premières cellules
[modifier | modifier le code]
Il existe plusieurs théories expliquant l'origine des petites molécules ayant conduit à l'apparition de la vie sur Terre. Elles auraient pu être apportées depuis l'espace par des météorites (météorite de Murchison), être formées dans les monts hydrothermaux sous les océans ou sous l'effet de la foudre dans une atmosphère réductrice (expérience de Miller-Urey). On dispose de peu de données expérimentales permettant de savoir ce qu'étaient les premières substances capables de se reproduire à l'identique. On pense que l'ARN a été la première molécule capable d'auto-réplication, car elle est capable à la fois de stocker l'information génétique et de catalyser des réactions chimiques (ribozymes), ce qui a été formulé dans le cadre de l'hypothèse du monde à ARN ; il existe cependant d'autres substances capables d'auto-réplication qui auraient pu précéder l'ARN dans cette fonction, par exemple des argiles comme la montmorillonite, qui sont capables de catalyser la polymérisation de l'ARN et la formation de membranes lipidiques[23],[24], ou encore les acides nucléiques peptidiques[25].
Les premières cellules sont apparues il y a au moins 3,5 milliards d'années[26],[27]. On considère actuellement que ces premières cellules étaient hétérotrophes. Les premières membranes cellulaires étaient probablement plus simples et plus perméables que les membranes actuelles. Les lipides forment spontanément des bicouches lipidiques dans l'eau, donnant des micelles et des vésicules ; ils auraient pu précéder l'ARN mais les premières membranes cellulaires auraient également pu être produites par des ribozymes ou même requérir des protéines structurelles pour se former[28].
Origine des eucaryotes
[modifier | modifier le code]On pense que les cellules d'eucaryotes dériveraient d'une communauté symbiotiques de procaryotes. Les organites comprenant de l'ADN tels que les mitochondries et les chloroplastes proviendraient respectivement de protéobactéries aérobies et de cyanobactéries devenues endosymbiotes d'un procaryote hôte.
Méthodes d'étude de la cellule in vitro
[modifier | modifier le code]Microscopie
[modifier | modifier le code]La microscopie optique ou photonique (résolution de +/- 0,25µm en lumière visible) permet l'observation de la structure des cellules eucaryotes. En effet, les microscopes photoniques peuvent grossir environ 1000 fois la taille du spécimen. Mais ce type de microscope n'est pas assez puissant pour l'étude des organites de la cellule.
La microscopie électronique (résolution de quelques Angströms) révèle l'ultrastructure de celles-ci et permet une observation plus poussée de la structure des cellules procaryotes comme eucaryotes. Dans la pratique, les microscopes électroniques modernes peuvent atteindre une résolution de 2 nm, ce qui représente une résolution 100 fois plus grande que celle des microscopes photoniques. Les microscopes électroniques ont permis de découvrir un grand nombre d'organites et une quantité d'autres structures subcellulaires invisibles au microscope optique. Toutefois, des percées techniques majeures ont donné un second souffle au microscope photonique : comme l'utilisation de marqueur fluorescent, de la microscopie confocale et avec déconvolution, qui permettent de donner des images 3D de la cellule avec une bonne netteté[29].
Marquage de molécules
[modifier | modifier le code]Pour étudier l'organisation subcellulaire des cellules au microscope, les tissus peuvent, en fonction de la méthode choisie, être vivants, ce qui permet une observation dynamique, ou fixés et préparés en coupes histologiques, ce qui permet en général une observation plus précise, mais figée et ponctuelle.
Localisation subcellulaire par l'utilisation de gènes rapporteurs tels que la GFP (green fluorescing protein) et la luciférase, par immunocytochimie, ou grâce à des molécules radioactives.
Différentes colorations, vitales ou non, permettent l'observation des structures au microscope optique : rouge neutre pour les vacuoles, violet dahlia ou cristal pour le noyau...
Étude des constituants cellulaires
[modifier | modifier le code]Isolement de structures : par choc osmotique, ou grâce à des détergents puis par centrifugation.
Purification des protéines : par électrophorèse, centrifugation, chromatographie, dialyse, etc.
Numération des cellules
[modifier | modifier le code]Il est fréquent de devoir compter le nombre de cellules vivantes dans une boîte de culture et de le comparer au nombre de cellules total, par exemple pour déterminer la toxicité d'un produit. L'une de ces méthodes de numération est réalisée grâce au test MTT.
Nombre et taille des cellules
[modifier | modifier le code]Le nombre des cellules propres à un organisme humain adulte est compris entre 1012 et 1016 ; une étude de 2013[30] l'estime à 3,72 × 1013. Selon une étude de 2014[4], les bactéries présentes dans ce même organisme (le microbiote, principalement dans le tube digestif), sont dix fois plus nombreuses (de l'ordre de 1015).
Les cellules les plus lourdes de tout le monde vivant sont les jaunes d'œuf d'autruche, de masse 1,2 à 1,9 kg, et les plus longues certains neurones de calmar géant ou de calmar colossal, dont l'axone peut atteindre 12 m[31].
La taille des cellules à paroi (plantes, bactéries, champignons, algues et certaines archées) varie de moins d'un micromètre (certaines bactéries) à plus d'un centimètre (algues géantes). Une étude de 2019[32],[33] montre que cette taille est directement liée à la rigidité de la paroi, définie comme le produit de son épaisseur par son module d'incompressibilité.
Histoire du concept
[modifier | modifier le code]- 1665 : Robert Hooke découvrit des cellules mortes dans du liège; ces cellules lui firent penser aux cellules d'un monastère, d'où le nom. Puis il observa des cellules dans des plantes vivantes, en utilisant les premiers microscopes. Hooke utilisa le mot cellule dans son ouvrage Micrographia, mais le sens était alors différent de celui d'aujourd'hui.
- 1668 : Francesco Redi montra que, contrairement à la conception de l'époque, les vers n'apparaissaient pas spontanément dans les cadavres mais provenaient d'œufs pondus par des mouches.
- 1839 : Theodor Schwann et Matthias Jakob Schleiden découvrirent que les plantes et les animaux sont tous faits de cellules, concluant que la cellule est l'unité commune de structure et de développement, ce qui fonda la théorie cellulaire. Schwann donna son nom aux cellules de Schwann.
- 1855 : Rudolf Virchow affirma que les cellules naissent du résultat de la division cellulaire (omnis cellula ex cellula), ce qui repose en termes cellulaires la question de l'œuf et de la poule. C'est précisément cette partie qui est attaquée par les tenants du créationnisme ou de son dernier avatar, le dessein intelligent.
- 1859 : Louis Pasteur (1822-1895) réfuta la croyance selon laquelle des formes de vie peuvent apparaître spontanément par génération spontanée.
- 1931 : Ernst Ruska construisit le premier microscope électronique en transmission à l'université Humboldt de Berlin, dont la résolution en 1935 était deux fois supérieure à celles des microscopes optiques, révélant des organites et des détails jusqu'alors invisibles.
- 1981 : Lynn Margulis publia Symbiosis in cell evolution, formulant la théorie endosymbiotique.
Théorie cellulaire
[modifier | modifier le code]- La cellule est l'unité constitutive des organismes vivants. Elle en est aussi l'unité fonctionnelle.
- L'organisme dépend de l'activité des cellules isolées ou groupées en tissus pour assurer les différentes fonctions.
- Les activités biochimiques des cellules sont coordonnées et déterminées par certaines structures présentes à l'intérieur des cellules.
- La multiplication des cellules permet le maintien des organismes et leur multiplication.
- Cette théorie est formulée en 1838 par Schleiden et Schwann : la cellule est unité de vie (tout ce qui est vivant est cellulaire). Cette théorie évoque également la présence d'organites à l'intérieur de ces mêmes cellules.
- La cellule c'est l'unité anatomique, morphologique et physiologique de tout être vivant.
- Cette théorie est ensuite complétée par Rudolf Virchow : chaque cellule trouve son origine auprès d'une autre cellule comme elle, on dit alors que les cellules sont les unités de reproduction du vivant, cela met donc fin à la théorie de la génération spontanée.
Notes et références
[modifier | modifier le code]- Félix Gaffiot, « Dictionnaire Latin Français », sur Le Gaffiot numérisé, Hachette, (consulté le ), p. 285.
- (en) Edmund Beecher Wilson, The cell in Development and Inheritance, New York City, The Macmillan Company, , 2e éd., Figure 2.
- (en) Ron Sender, Shai Fuchs et Ron Milo, « Are We Really Vastly Outnumbered? Revisiting the Ratio of Bacterial to Host Cells in Humans », Cell, vol. 164, no 3, , p. 337–340 (DOI 10.1016/j.cell.2016.01.013).
- (en) « Mouth bacteria can change its diet, supercomputers reveal », sur MedicalXpress, (consulté le ).
- (en) Allen P. Nutman, Vickie C. Bennett, Clark R. L. Friend, Martin J. Van Kranendonk et Allan R. Chivas, « Rapid emergence of life shown by discovery of 3,700-million-year-old microbial structures », Nature, vol. 537, , p. 535-538 (DOI 10.1038/nature19355).
- (en) Elizabeth Pennisi, « Life may have originated on Earth 4 billion years ago, study of controversial fossils suggests », Science, (DOI 10.1126/science.aar7944).
- (en) Peter Satir et Søren T. Christensen, « Structure and function of mammalian cilia », Histochemistry and Cell Biology, vol. 129, no 6, , p. 687-693 (PMID 18365235, PMCID 2386530, DOI 10.1007/s00418-008-0416-9, lire en ligne)
- Geoffrey M. Cooper, La cellule : une approche moléculaire, De Boeck Université, 1999, p. 502
- (en) Katharine A. Michie et Jan Löwe, « Dynamic Filaments of the Bacterial Cytoskeleton », Annual Review of Biochemistry, vol. 75, , p. 467-492 (PMID 16756499, DOI 10.1146/annurev.biochem.75.103004.142452, lire en ligne)
- (en) Jean-François Ménétret, Julia Schaletzky, William M. Clemons Jr., Andrew R. Osborne, Sigrid S. Skånland, Carilee Denison, Steven P. Gygi, Don S. Kirkpatrick, Eunyong Park, Steven J. Ludtke, Tom A. Rapoport et Christopher W. Akey, « Ribosome Binding of a Single Copy of the SecY Complex: Implications for Protein Translocation », Molecular Cell, vol. 28, no 6, , p. 1083-1092 (PMID 18158904, DOI 10.1016/j.molcel.2007.10.034, lire en ligne)
- Jean-Claude Callen, Roland Perasso, Biologie cellulaire : Des molécules aux organismes, 2e édition, Dunod, Paris, 2005, p. 36-37. (ISBN 2-10-049236-5)
- (en) Stephen J. Elledge, « Cell Cycle Checkpoints: Preventing an Identity Crisis », Science, vol. 274, no 5293, , p. 1664-1672 (PMID 8939848, DOI 10.1126/science.274.5293.1664, Bibcode 1996Sci...274.1664E, lire en ligne)
- (en) « Subdiffraction multicolor imaging of the nuclear periphery with 3D structured illumination microscopy », Science, vol. 320, no 5881, , p. 1332-1336 (PMID 18535242, PMCID 2916659, DOI 10.1126/science.1156947, lire en ligne)
- (en) Edward Parr, « The default state of the cell: Quiescence or proliferation? », BioEssays, vol. 34, no 1, , p. 36-37 (ISSN 1521-1878, PMID 22006841, PMCID 3266487, DOI 10.1002/bies.201100138, lire en ligne, consulté le )
- (en) Ana M. Soto, Giuseppe Longo, Maël Montévil et Carlos Sonnenschein, « The biological default state of cell proliferation with variation and motility, a fundamental principle for a theory of organisms », Progress in Biophysics and Molecular Biology, from the Century of the Genome to the Century of the Organism: New Theoretical Approaches, vol. 122, no 1, , p. 16-23 (DOI 10.1016/j.pbiomolbio.2016.06.006, lire en ligne, consulté le )
- Carlos Sonnenschein, Ana M. Soto, Michel Morange et Thomas Heams, La société des cellules : Nouvelle approche du cancer, Editions Syllepse, (ISBN 9782849500385)
- (en) Maël Montévil, Lucia Speroni, Carlos Sonnenschein et Ana M. Soto, « Modeling mammary organogenesis from biological first principles: Cells and their physical constraints », Progress in Biophysics and Molecular Biology, from the Century of the Genome to the Century of the Organism: New Theoretical Approaches, vol. 122, no 1, , p. 58-69 (DOI 10.1016/j.pbiomolbio.2016.08.004, lire en ligne, consulté le )
- (en) Revathi Ananthakrishnan et Allen Ehrlicher, « The Forces Behind Cell Movement », International Journal of Biological Sciences, vol. 3, no 5, , p. 303-317 (PMID 17589565, PMCID 1893118, DOI 10.7150/ijbs.3.303, lire en ligne)
- (en) M. R. Walter, John H. Oehler et Dorothy Z. Oehler, « Megascopic Algae 1300 Million Years Old from the Belt Supergroup, Montana: A Reinterpretation of Walcott's Helminthoidichnites », Journal of Paleontology, vol. 50, no 5, , p. 872-881 (JSTOR 1303584, lire en ligne)
- (en) T. M. Han et B. Runnegar, « Megascopic eukaryotic algae from the 2.1-billion-year-old negaunee iron-formation, Michigan », Science, vol. 257, no 5067, , p. 232-235 (PMID 1631544, DOI 10.1126/science.1631544, Bibcode 1992Sci...257..232H, lire en ligne)
- (en) Abderrazak El Albani, Stefan Bengtson, Donald E. Canfield, Andrey Bekker, Roberto Macchiarelli, Arnaud Mazurier, Emma U. Hammarlund, Philippe Boulvais, Jean-Jacques Dupuy, Claude Fontaine, Franz T. Fürsich, François Gauthier-Lafaye, Philippe Janvier, Emmanuelle Javaux, Frantz Ossa Ossa, Anne-Catherine Pierson-Wickmann, Armelle Riboulleau, Paul Sardini, Daniel Vachard, Martin Whitehouse et Alain Meunier, « Large colonial organisms with coordinated growth in oxygenated environments 2.1 Gyr ago », Nature, vol. 466, no 7302, , p. 100-104 (PMID 20596019, DOI 10.1038/nature09166, lire en ligne)
- (en) Robert Riding, « The term stromatolite: towards an essential definition », Lethaia, vol. 32, no 4, , p. 321-330 (DOI 10.1111/j.1502-3931.1999.tb00550.x, lire en ligne)
- (en) Harvard University, « Clay-armored bubbles may have formed first protocells: Minerals could have played a key role in the origins of life », sur ScienceDaily, (consulté le )
- (en) Anand Bala Subramaniam, Jiandi Wan, Arvind Gopinath et Howard A. Stone, « Semi-permeable vesicles composed of natural clay », Soft Matter, vol. 7, no 6, , p. 2600-2612 (DOI 10.1039/c0sm01354d, Bibcode 2011SMat....7.2600S, lire en ligne)
- (en) Leslie E. Orgel, « The origin of life—a review of facts and speculations », Trends in Biochemical Sciences, vol. 23, no 12, , p. 491-495 (PMID 9868373, DOI 10.1016/S0968-0004(98)01300-0, lire en ligne)
- (en) J. William Schopf, Anatoliy B. Kudryavtsev, Andrew D. Czaja et Abhishek B. Tripathi, « Evidence of Archean life: Stromatolites and microfossils », Precambrian Research, vol. 158, nos 3-4, , p. 141-155 (DOI 10.1016/j.precamres.2007.04.009, lire en ligne)
- (en) J. William Schopf, « Fossil evidence of Archaean life », Philosophical Transactions: Biological Sciences, vol. 361, no 1470, , p. 869-885 (PMID 16754604, PMCID 1578735, DOI 10.1098/rstb.2006.1834, JSTOR 20209690, lire en ligne)
- (en) Gareth Griffiths, « Cell evolution and the problem of membrane topology », Nature Reviews, vol. 8, no 12, , p. 1018-1024 (PMID 17971839, DOI 10.1038/nrm2287, lire en ligne)
- Campbell Biologie 9e édition Pearson, Chap. 6, Exploration de la Cellule
- An estimation of the number of cells in the human body., Ann Hum Biol. 2013 Nov-Dec;40(6):463-71. doi: 10.3109/03014460.2013.807878. Epub 2013 Jul 5.
- Frédéric Flamant, De l'œuf à la poule, Belin, , 159 p..
- « Pour la taille d’une cellule, c’est la rigidité de l’enveloppe qui compte », sur INSB, (consulté le ).
- (en) Valeria Davì, Louis Chevalier, Haotian Guo, Hirokazu Tanimoto, Katia Barrett et al., « Systematic mapping of cell wall mechanics in the regulation of cell morphogenesis », PNAS, vol. 116, no 28, , p. 13833-13838 (DOI 10.1073/pnas.1820455116).
Voir aussi
[modifier | modifier le code]Articles connexes
[modifier | modifier le code]- Biologie cellulaire
- Cellule souche
- Cellule végétale
- Culture cellulaire
- Cytota (taxon regroupant la vie cellulaire)
- Dernier ancêtre commun universel
(DACU, LUCA en anglais) - Électrophysiologie
- Histologie
- Mécanique cellulaire
- Organisme unicellulaire
- Respiration cellulaire
Bibliographie
[modifier | modifier le code]- Jean Claude Callen, Biologie cellulaire : Des molécules aux organismes.
- Christian Sardet, Les cellules : Une histoire de la vie, Paris, Les Éditions Ulmer, , 224 p. (ISBN 9782379223563, lire en ligne) :
« Comprendre l'origine et le fonctionnement des cellules, c'est découvrir l'unité de la vie sur Terre. Un voyage fascinant dans le temps et l'infiniment petit ! »
- Christian Sardet, Plancton : Aux origines du vivant, Paris, Les Éditions Ulmer, , 215 p. (ISBN 978-2-84138-634-5)
Liens externes
[modifier | modifier le code]
- Ressources relatives à la santé :
- Notices dans des dictionnaires ou encyclopédies généralistes :
- Dossier Sagascience du CNRS : la cellule animale
- Qu'est ce qu'une cellule : La cellule en microcinéma
- Schéma détaillé d'une cellule animale.
- Schéma détaillé d'une cellule végétale.
- Exploration 3D d'une cellule : sinauer.com, aimediaserver.com
- (de) Cell Biology - Graphics
- Voir la vidéo the inner life of the cell faite par BioVision (harvard)
- Voir la vidéo sur la cellule dans l'encyclopédie médicale Vulgaris