Spirochaetota

| Domaine | Bacteria |
|---|
- Spirochaetota Whitman et al. 2018
- Spirochaetaeota Oren et al. 2015
- Spirochaetae Cavalier-Smith 2002
- Spirochaetes Garrity & Holt 2001
- Spirochaetae (Ehrenberg 1855) Cavalier-Smith 1998
Les Spirochaetota (en français les Spirochètes) sont un embranchement (ou phylum, ou encore division) du règne des Bacteria. Son nom provient de Spirochaeta qui est le genre type de cet embranchement. Ce sont des bactéries hélicoïdales à Gram négatif, très flexibles, mobiles à l'aide d'une structure intracellulaire spécifique appelée flagelle périplasmique. Certains genres sont des parasites animaux et en particulier humains comme par exemple Borrelia, Treponema ou encore Leptospira, pouvant entrer dans le cadre des maladies émergentes ou ré-émergentes. Ces maladies parfois graves ou mortelles sont difficiles à diagnostiquer en raison de symptômes variés, variables et peu spécifiques[2][source insuffisante].
En 2022 selon la LPSN (1er décembre 2022)[3] cet embranchement ne comporte qu'une seule classe, les Spirochaetia Paster 2020.
Taxonomie
[modifier | modifier le code]La taxonomie des Spirochaetota a connu de nombreuses évolutions. L'ancien ordre des Spirochétales était autrefois inclus dans les protozoaires. Cet ordre des Spirochaetales est considéré comme un ordre à part depuis le milieu du XXe siècle[4]. Historiquement, les spirochètes ont formé un phylum appelé Spirochaetae ne contenant qu'une seule et unique classe Spirochaetes, un seul ordre des Spirochaetales divisé en trois familles. Ces anciennes partitions ne sont plus reconnues depuis 2014LPSN (6 octobre 2023)[5].
Parmi les membres importants, citons :
- les espèces du genre Leptospira, qui provoquent la leptospirose ;
- le genre Borrelia, parmi lequel :
- Borrelia burgdorferi, l'un des agents de la maladie de Lyme (le plus courant en Amérique du Nord) ;
- Borrelia duttoni
- l'espèce Treponema pallidum, agent de la syphilis.
Systématique
[modifier | modifier le code]Le nom correct complet (avec auteur) de ce taxon est Spirochaetota Garrity & Holt 2021[5].
Le genre type est : Spirochaeta Ehrenberg 1835[5].
Étymologie
[modifier | modifier le code]L'étymologie du nom de ce phylum est la suivante : Spi.ro.chae.to’ta. N.L. fem. n. Spirochaeta, genre type de ce phylum; L. neut. pl. n. suff. -ota, suffixe pour définir un phylum; N.L. neut. pl. n. Spirochaetota, Le phylum des Spirochaeta[5].
Synonymie
[modifier | modifier le code]Spirochaetota a pour synonymes[5] :
- Spirochaetae (Ehrenberg 1855) Cavalier-Smith 1998
- Spirochaetaeota Oren et al. 2015
- Spirochaetes Garrity & Holt 2001
- Spirochaetota Whitman et al. 2018
Liste des classes
[modifier | modifier le code]Selon la base de référence LPSN (6 octobre 2023)[5], ce phylum ne comporte qu'une seule classe :
- Spirochaetia Paster 2020
Liste des ordres
[modifier | modifier le code]Selon la LPSN (6 octobre 2023)[5], il existe 4 ordres publiés de manière valide et au nom correct :
- ordre des Brachyspirales Gupta et al. 2014
- ordre des Brevinematales Gupta et al. 2014
- ordre des Leptospirales Gupta et al. 2014
- ordre des Spirochaetales Buchanan 1917
Histoire évolutive
[modifier | modifier le code]Un spirochète (a priori intestinal) très ancien a été découvert en République dominicaine, fossilisé, dans de l'ambre daté du Miocène (il y a 20 millions d'années, bien avant l'apparition du genre Homo). Dénommé Pillotina sp. il a été trouvé dans un fossile de termite (dans une vésicule associée à une membrane) de l'espèce Mastotermes electrodominicus (de la famille des Mastotermitidae). Selon Ø. Brorson & al. (2009)[6] ce fossile bien conservé[7] et des observations contemporaines in vivo[8] laissent penser que des spirochètes vivent depuis des dizaines de millions d'années sur Terre comme symbiotes (non-pathogène) d'insectes ; selon cette équipe, des symbioses tissulaires spirochètes-animaux précèdent probablement de millions d'années la nécrotrophie tissulaire induite par les spirochètes associée aux états pathologiques (ex : spirochétoses de type maladie de Lyme ou syphilis[6].
Histoire scientifique
[modifier | modifier le code]Des bactéries spiralées ont été découvertes à l'intérieur de tiques par Dutton qui au tout début du XXe siècle s'intéressait à des fièvres apparaissant chez des humains après morsure de tique, dans l'Est du Congo, avant la publication (en 1905 de Dutton et Todd[9]. Ces bactéries spiralées se développaient dans ce cas à partir d'un grand nombre de « granules » observés chez des tiques infectées exposées à une température de 25 °C ou plus.
Peu après (en 1911), E. Hindle découvre que l'injection à des souris de laboratoire de la « forme granulaire » (pas de bactérie hélicoïdale apparente dans le milieu de culture) de ces bactéries spirochètes (entre-temps dénommées Borrelia duttoni) provoquaient chez ces animaux des symptômes d'infection[10].
40 ans plus tard, E.G Hampp EG (en 1951) note que des formes kystiques de Borrelia vincentii restées « granulaires » durant 31 mois se sont converties en spirochètes hélicoïdaux mobiles une fois transférées dans un milieu favorable à la croissance de la bactérie[11], ce qui plaidait pour un cycle de vie pléiomorphe (à plusieurs formes) et laisse penser que la forme kystique a aussi une signification médicale.
D'autres chercheurs n'ont pas réussi à trouver ces propagules (spirochètes en dormance ou enkystés) ou les ont considéré comme étant des spirochètes mourants. Cette littérature scientifique précoce sur le sujet a ensuite été oubliée jusque dans les années 2010[12], ce qui était injustifié selon Ø Brorson et ses collègues (2009)[6] .
Morphologie, structure
[modifier | modifier le code]Les spirochètes sont des cellules longues de forme hélicoïdale (en « tire-bouchon »), mais elles peuvent aussi former des amas coloniaux en forme de boule (voir fin de la vidéo en microscopie, plus bas), ou changer d'aspect dans certaines circonstances.
Elles se distinguent des autres bactéries par la présence de longs filaments axiaux internes, d'un diamètre uniforme de 18 nm, situés dans le sens de la longueur, entre la membrane cellulaire et la paroi cellulaire, appelés filaments axiaux. Du point de vue chimique et structural, ces filaments axiaux sont semblables aux flagelles bactériens. Leur nombre varie de deux à plusieurs centaines et leur localisation est unique dans le monde bactérien. Souvent, ils se chevauchent au centre de la cellule.
Ces pseudo-flagelles — par un mécanisme encore mal compris (plusieurs modèles sont proposés) — confèrent aux spirochètes une mobilité et rapidité exceptionnelles dans les milieux visqueux. Les spirochètes peuvent s'enfoncer rapidement dans un milieu légèrement gélatineux ou se déplacer par reptation sur des milieux solides.
Leur enveloppe externe, tubulaire et très élastique, est également originale. Elle est composée d'un assemblage de lipides, protéines et sucres et chez certaines espèces crénelée et très complexe (cinq couches chez Pillotina). Elle peut se séparer de la cellule en conditions adverses, par exemple en contact avec un détergent de type laurylsulfate de sodium ou dodécylsulfate de sodium (la cellule prend alors une forme sphérique, a priori non viable).
Aspects morphologiques
[modifier | modifier le code]
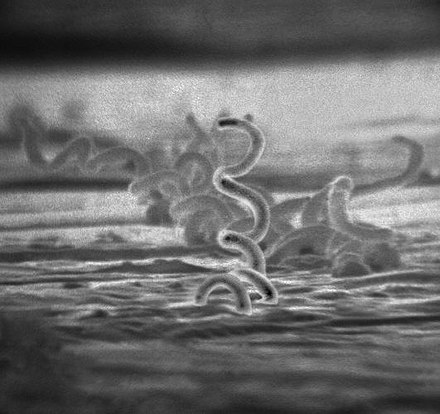



La microscopie électronique à transmission permet d’observer trois principaux types de « structure classique » :
- Les bactéries à Gram + ;
- Les bactéries à Gram - ;
- Les spirochètes.
Mobilité
[modifier | modifier le code]Dans les années 2000, les capacités motrices exceptionnelles (et bidirectionnelles) de certains spirochètes (borrélies) ont été mises en évidence. Leur mode de locomotion, et leur forme spiralée — dans certains milieux (visqueux) de l'organisme —, les aident à facilement distancer les macrophages phagocytaires qui devraient normalement les détruire.
En 2008, l'observation en temps réel en vidéomicroscopie à contraste de phase a montré[13],[14] que Borrelia burgdorferi se déplace à une vitesse moyenne de 1636 micromètres par minute (moyenne pour 28 bactéries observées), avec une vitesse maximale de 2 800 µm/min (2,8 mm/min) chez 3 des bactéries observées, soit la vitesse la plus rapide enregistrée pour un spirochète, et de deux ordres de grandeur au-dessus de la vitesse d'un polynucléaire neutrophile humain, réputé être la plus rapide de nos cellules mobiles.
Ces spirochètes disposent de 2 séries de flagelles leur permettant de se déplacer dans des directions opposées le long de leur axe (en avant ou en arrière). Au contraire des autres bactéries mobiles connues, il s'agit de flagelles internes (organites dites « endoflagelles » ou « flagelles périplasmiques »).
Ces flagelles baignent dans le milieu interne (periplasme) de la cellule bactérienne. Chaque flagelle est attaché par l'une de ses deux extrémités à l'une des extrémités de la cellule. Leur longueur diffère selon l'espèce de spirochète. Chez certaines espèces les flagelles (dont le nombre varie aussi selon l'espèce) se chevauchent au centre de la cellule. Les flagelles sont animés d'un mouvement rotatoire dans le milieu interne (espace périplasmique).
Ces flagelles internes sont très peu efficaces dans un liquide très fluide : à la manière d'un ver de terre dans l'eau, la bactérie doit y effectuer des flexions et rotations qui lui font consommer beaucoup d'énergie. Elle n'y progresse qu'à une vitesse d'environ 4,25 μm/s, ou 255 μm/min [15]. Cependant, à la manière du ver de terre dans un substrat qui lui oppose une résistance, les borrélias deviennent très rapides quand le degré de « viscoélasticité » du milieu augmente ; la bactérie devient ainsi « très » rapide (jusqu'à environ 2 000 μm/min, soit 2 mm/min) et bien mieux capable de se diriger dans un fluide plus visqueux comparable au milieu extracellulaire de notre organisme (par exemple dans la méthylcellulose dont les propriétés de viscosité imitent celles de la matrice extracellulaire, de celles des différents acides hyaluroniques qui composent nos humeurs vitreuses et de celles des lubrifiants articulaires dont synovie). La vitesse des Spirochètes observés dans de tels « gels » est directement liée à la viscoélasticité du milieu, plus qu'à sa teneur en nutriments. Ainsi a-t-on observé des spirochètes restant mobiles durant 3 heures dans un tel milieu, malgré une teneur en nutriments de 100 fois inférieure à la norme retenue pour les milieux de culture[16], de même que sur les surfaces où elles sont soupçonnées d'être présentes[17],[18].
Le mécanisme expliquant la motilité semble également varier selon les espèces de spirochètes. Le modèle mis au point pour les leptospirae, qui reste convaincant pour certains spirochètes n'explique pas le mouvement de Borrelia burgdorferi qui « nage » différemment et très efficacement dans les gels. Les mouvements de cette borrélie ont été observés en microscopie, et, la comparaison avec ceux de mutants de la même bactérie associée à des études moléculaires sur les protéines et les gènes impliqués dans la mobilité devraient bientôt éclairer ce qui la différencie des autres spirochètes, sous cet aspect. Ces études et les premières séquences génomiques disponibles montrent déjà que les filaments protéiques qui forment les flagelles internes des spirochètes comptent parmi les plus complexes des flagelles bactériens connus et qu'ils sont organisés et régulés par un grand nombre de gènes impliqués dans la motilité des spirochètes. Ceci laisse penser que ces borrélies ont depuis longtemps adapté leur vie à ces milieux plus visqueux.
La tomographie appliquée à la microscopie pourrait permettre de mieux comprendre comment fonctionne le « moteur biologique flagellaire » qui permet aux spirochètes de se déplacer si rapidement dans les liquides ou fluides visqueux (pour l'instant à partir d'un organisme (Treponema) simple : Treponema primitia[19]).
Paléoécologie
[modifier | modifier le code]Des biologistes bactériologistes comme Lynn Margulis estiment que certains des spirochaetes ont probablement depuis longtemps coévolué avec l'homme et son système immunitaire.
En 2009, L. Margulis, avec six autres scientifiques, a estimé dans un article qu'« une recherche détaillée qui corrélant l'histoire des spirochètes symbiotiques aux changements du système immunitaire des vertébrés est grandement nécessaire ». Cet article exhorte une « nouvelle recherche de l'histoire naturelle des mammifères et des tiques ainsi que de la transmission vénérienne des spirochètes en relation avec la détérioration du système immunitaire humain »[20].
Photographies
[modifier | modifier le code]-
Deux spirochètes, dans un frottis sanguin, au milieu de globules rouges
-
Groupe de spirochètes dans une goutte d'eau provenant d'un caniveau


Notes et références
[modifier | modifier le code]- Oren A & Garrity GM « Valid publication of the names of forty-two phyla of prokaryotes » Int J Syst Evol Microbiol. 2021;71(10):005056. Accès libre.
- Capucine Kosossey Vrain (2004), La leptospirose canine : revue bibliographique ; thèse de doctorat vétérinaire (École Nationale vétérinaire d’Alfort, France) [PDF], 135 p.
- List of Prokaryotic names with Standing in Nomenclature (LPSN), consulté le 1er décembre 2022
- Bharti AR et al. (2003) Leptospirosis: a zoonotic disease of global importance. Lancet, 3, 757-771.
- List of Prokaryotic names with Standing in Nomenclature (LPSN), consulté le 6 octobre 2023
- Brorson Ø, Brorson S.H, Scythes J, MacAllister J, Wier A & Margulis L (2009) Destruction of spirochete Borrelia burgdorferi round-body propagules (RBs) by the antibiotic tigecycline. Proc. Natl. Acad. Sci. USA, 106, 18656–18661. (résumé)
- Wier A, et al. (2002) Spirochete and protist symbionts of a termite (Mastotermes electrodominicus) in Miocene amber. Proc Natl Acad Sci USA 99:1410–1413.
- Wier A, MacAllister J & Margulis L (2007) Hibernacular behavior of spirochetes inside membrane-bounded vesicles of the termite protist Staurojoenina assimilis. Symbiosis 44:75– 83.
- Dutton JE & Todd JL (1905) The nature of human tick fever in the eastern part of the Congo Free State. Liverpool Sch Trop Med Hyg, Mem XVII, pp 1–18.
- Hindle E (1911) On the life-cycle of Spirochaeta gallinarum. Parasitology 4:463– 477.
- Hampp EG (1951) Further studies on the significance of spirochetal granules. J Bacteriol 62:347–349.
- Margulis L, et al. (2009) Spirochete round bodies, syphilis, Lyme disease & AIDS : Resurgence of ‘‘the Great Imitator’’? Symbiosis 47:51–58
- Stephen E. Malawista1 et Anne de Boisfleury Chevance ; Clocking the Lyme Spirochete ; PLoS ONE. 2008 ; 3(2) : e1633. Mis en ligne le 20 février 2008 ; DOI : 10.1371/journal.pone.0001633. Lire l'étude en ligne (PubMed) (Licence Creative Commons).
- Vidéo présentant des spirochètes en mouvement ; source : Lyme disease research database.
- Goldstein, SF ; Charon, NW ; Kreiling, JA. Borrelia burgdorferi swims with a planar waveform similar to that of eukaryotic flagella. Proc Natl Acad Sci U S A. 1994;91:3433–7.
- Kimsey, RB ; Spielman, A. Motility of Lyme disease spirochetes in fluids as viscous as the extracellular matrix. J Infect Dis. 1990;162:1205–8. Résumé PubMed.
- Li, C ; Motaleb, A ; Sal, M ; Goldstein, SF ; Charon, NW. Spirochete periplasmic flagella and motility. J Mol Microbiol Biotechnol. 2000;2:345–54. Résumé (PubMed).
- Charon, NW ; Goldstein, SF. Genetics of motility and chemotaxis of a fascinating group of bacteria: the spirochetes. Annu Rev Genet. 2002;36:47–73. Résumé (PubMed).
- Electron cryotomography of T. primitia and its periplasmic flagellar motor, Nature. Pour plus de détails voir : Gavin E. Murphy, Jared R. Leadbetter et Grant J. Jensen, In situ structure of the complete Treponema primitia flagellar motor ; Nature 442, 1062-1064 (31 août 2006) ; 10.1038/nature05015 (résumé).
- (en) Syphilis, Lyme disease & AIDS: Resurgence of “the great imitator”?, SYMBIOSIS vol. 47, no 1 (2009), pp. 51-58.
Voir aussi
[modifier | modifier le code]Articles connexes
[modifier | modifier le code]Liens externes
[modifier | modifier le code]- Classe Spirochaetes
- (en) Référence Tree of Life Web Project : Spirochaetes
- (en) Référence Catalogue of Life : Spirochaetes Cavalier-Smith, 2002 (consulté le )
- (en) Référence NCBI : Spirochaetes (taxons inclus)
- Ordre Spirochaetales
- (en) Référence Tree of Life Web Project : Spirochaetales
- (en) Référence Catalogue of Life : Spirochaetales Buchanan, 1917 (consulté le )
- (fr + en) Référence ITIS : Spirochaetales
- (en) Référence NCBI : Spirochaetales (taxons inclus)
Bibliographie
[modifier | modifier le code]- Judith Miklossy (2011), Alzheimer's disease - a neurospirochetosis. Analysis of the evidence following Koch's and Hill's criteria ; J Neuroinflammation ; 8:90 ; en ligne : 2011-08-04. Doi:10.1186/1742-2094-8-90 ; PMCID:PMC3171359
- Zelger B, Eisendle K & Mensing C (2007), Detection of spirochetal microorganisms by focus-floating microscopy in necrobiotic xanthogranuloma , J Am Acad Dermatol ; 57:1026–30. (en)