Célula

La célula (del latín cellula, diminutivo de cella, ‘celda’)[1] es la unidad morfológica y funcional de todo ser vivo. De hecho, la célula es el elemento de menor tamaño que puede considerarse vivo.[2] De este modo, puede clasificarse a los organismos vivos según el número de células que posean: si solo tienen una, se les denomina unicelulares (como pueden ser los protozoos o las bacterias, organismos microscópicos); si poseen más, se les llama pluricelulares. En estos últimos el número de células es variable: de unos pocos cientos, como en algunos nematodos, a cientos de billones (1014), como en el caso del ser humano. Las células suelen poseer un tamaño de 10 µm y una masa de 1 ng, si bien existen células mucho mayores.

La teoría celular, propuesta en 1838 para los vegetales y en 1839 para los animales,[3] por Matthias Jakob Schleiden y Theodor Schwann, postula que todos los organismos están compuestos por células, y que todas las células derivan de otras precedentes. De este modo, todas las funciones vitales emanan de la maquinaria celular y de la interacción entre células adyacentes; además, la tenencia de la información genética, base de la herencia, en su ADN permite la transmisión de aquella de generación en generación.[4]
La aparición del primer organismo vivo sobre la Tierra suele asociarse al nacimiento de la primera célula. Si bien existen muchas hipótesis que especulan cómo ocurrió, usualmente se describe que el proceso se inició gracias a la transformación de moléculas inorgánicas en orgánicas bajo unas condiciones ambientales adecuadas; tras esto, dichas biomoléculas se asociaron dando lugar a entes complejos capaces de autorreplicarse. Existen posibles evidencias fósiles de estructuras celulares en rocas datadas en torno a 4 o 3,5 miles de millones de años (gigaaños o Ga).[5][6][nota 1] Se han encontrado evidencias muy fuertes de formas de vida unicelulares fosilizadas en microestructuras en rocas de la formación Strelley Pool, en Australia Occidental, con una antigüedad de 3,4 Ga.[cita requerida] Se trataría de los fósiles de células más antiguos encontrados hasta la fecha. Evidencias adicionales muestran que su metabolismo sería anaerobio y basado en el sulfuro.[7]
Tipos celulares
Existen dos grandes tipos celulares:
- Célula procariota, propia de los procariontes, que comprende las células de arqueas y bacterias.
- Célula eucariota, propia de los eucariontes, tales como la célula animal, célula vegetal, y las células de hongos y protistas.
Historia y teoría celular
La historia de la biología celular ha estado ligada al desarrollo tecnológico que pudiera sustentar su estudio. De este modo, el primer acercamiento a su morfología se inicia con la popularización del microscopio rudimentario de lentes compuestas en el siglo XVII, se suplementa con diversas técnicas histológicas para microscopía óptica en los siglos XIX y XX y alcanza un mayor nivel resolutivo mediante los estudios de microscopía electrónica, de fluorescencia y confocal, entre otros, ya en el siglo XX. El desarrollo de herramientas moleculares, basadas en el manejo de ácidos nucleicos y enzimas permitieron un análisis más exhaustivo a lo largo del siglo XX.[8]
Descubrimiento

Las primeras aproximaciones al estudio de la célula surgieron en el siglo XVII;[9] tras el desarrollo a finales del siglo XVI de los primeros microscopios.[10] Estos permitieron realizar numerosas observaciones, que condujeron en apenas doscientos años a un conocimiento morfológico relativamente aceptable. A continuación se enumera una breve cronología de tales descubrimientos:
- 1665: Robert Hooke publicó los resultados de sus observaciones sobre tejidos vegetales, como el corcho, realizadas con un microscopio de 50 aumentos construido por él mismo. Este investigador fue el primero que, al ver en esos tejidos unidades que se repetían a modo de celdillas de un panal, las bautizó como elementos de repetición, «células» (del latín cellulae, celdillas). Pero Hooke solo pudo observar células muertas por lo que no pudo describir las estructuras de su interior.[11]
- Década de 1670: Anton van Leeuwenhoek observó diversas células eucariotas (como protozoos y espermatozoides) y procariotas (bacterias).
- 1745: John Needham describió la presencia de «animálculos» o «infusorios»; se trataba de organismos unicelulares.

- Década de 1830: Theodor Schwann estudió la célula animal; junto con Matthias Schleiden postularon que las células son las unidades elementales en la formación de las plantas y animales, y que son la base fundamental del proceso vital.
- 1831: Robert Brown describió el núcleo celular.
- 1839: Purkinje observó el citoplasma celular.
- 1857: Kölliker identificó las mitocondrias.
- 1858: Rudolf Virchow postuló que todas las células provienen de otras células.
- 1860: Pasteur realizó multitud de estudios sobre el metabolismo de levaduras y sobre la asepsia.
- 1880: August Weismann descubrió que las células actuales comparten similitud estructural y molecular con células de tiempos remotos.
- 1931: Ernst Ruska construyó el primer microscopio electrónico de transmisión en la Universidad de Berlín. Cuatro años más tarde, obtuvo una resolución óptica doble a la del microscopio óptico.
- 1981: Lynn Margulis publica su hipótesis sobre la endosimbiosis serial, que explica el origen de la célula eucariota.[12]
Teoría celular
El concepto de célula como unidad anatómica y funcional de los organismos surgió entre los años 1830 y 1880, aunque fue en el siglo XVII cuando Robert Hooke describió por vez primera la existencia de las mismas, al observar en una preparación vegetal la presencia de una estructura organizada que derivaba de la arquitectura de las paredes celulares vegetales. En 1830 se disponía ya de microscopios con una óptica más avanzada, lo que permitió a investigadores como Theodor Schwann y Matthias Schleiden definir los postulados de la teoría celular, la cual afirma, entre otras cosas:
- Que la célula es una unidad morfológica de todo ser vivo: es decir, que en los seres vivos todo está formado por células o por sus productos de secreción.
- Este primer postulado sería completado por Rudolf Virchow con la afirmación Omnis cellula ex cellula, la cual indica que toda célula deriva de una célula precedente (biogénesis). En otras palabras, este postulado constituye la refutación de la teoría de generación espontánea o ex novo, que hipotetizaba la posibilidad de que se generara vida a partir de elementos inanimados.[13]
- Un tercer postulado de la teoría celular indica que las funciones vitales de los organismos ocurren dentro de las células, o en su entorno inmediato, y son controladas por sustancias que ellas secretan. Cada célula es un sistema abierto, que intercambia materia y energía con su medio. En una célula ocurren todas las funciones vitales, de manera que basta una sola de ellas para que haya un ser vivo (que será un individuo unicelular). Así pues, la célula es la unidad fisiológica de la vida.
- El cuarto postulado expresa que cada célula contiene toda la información hereditaria necesaria para el control de su propio ciclo y del desarrollo y el funcionamiento de un organismo de su especie, así como para la transmisión de esa información a la siguiente generación celular.[14]
Definición
Se define a la célula como la unidad morfológica y funcional de todo ser vivo. De hecho, la célula es el elemento de menor tamaño que puede considerarse vivo. Como tal posee una membrana de fosfolípidos con permeabilidad selectiva que mantiene un medio interno altamente ordenado y diferenciado del medio externo en cuanto a su composición, sujeta a control homeostático, la cual consiste en biomoléculas y algunos metales y electrolitos. La estructura se automantiene activamente mediante el metabolismo, asegurándose la coordinación de todos los elementos celulares y su perpetuación por replicación a través de un genoma codificado por ácidos nucleicos. La parte de la biología que se ocupa de ella es la citología.
Características
Las células, como sistemas termodinámicos complejos, poseen una serie de elementos estructurales y funcionales comunes que posibilitan su supervivencia; no obstante, los distintos tipos celulares presentan modificaciones de estas características comunes que permiten su especialización funcional y, por ello, la ganancia de complejidad.[15] De este modo, las células permanecen altamente organizadas a costa de incrementar la entropía del entorno, uno de los requisitos de la vida.[16]
Características estructurales

- Individualidad: Todas las células están rodeadas de una envoltura (que puede ser una bicapa lipídica desnuda, en células animales; una pared de polisacárido, en hongos y vegetales; una membrana externa y otros elementos que definen una pared compleja, en bacterias Gram negativas; una pared de peptidoglicano, en bacterias Gram positivas; o una pared de variada composición, en arqueas)[9] que las separa y comunica con el exterior, que controla los movimientos celulares y que mantiene el potencial de membrana.
- Contienen un medio interno acuoso, el citosol, que forma la mayor parte del volumen celular y en el que están inmersos los orgánulos celulares.
- Poseen material genético en forma de ADN, el material hereditario de los genes, que contiene las instrucciones para el funcionamiento celular, así como ARN, a fin de que el primero se exprese.[17]
- Tienen enzimas y otras proteínas, que sustentan, junto con otras biomoléculas, un metabolismo activo.
Características funcionales

Las células vivas son un sistema bioquímico complejo. Las características que permiten diferenciar las células de los sistemas químicos no vivos son:
- Nutrición. Las células toman sustancias del medio, las transforman de una forma a otra, liberan energía y eliminan productos de desecho, mediante el metabolismo.
- Crecimiento y multiplicación. Las células son capaces de dirigir su propia síntesis. A consecuencia de los procesos nutricionales, una célula crece y se divide, formando dos células, en una célula idéntica a la célula original, mediante la división celular.
- Diferenciación. Muchas células pueden sufrir cambios de forma o función en un proceso llamado diferenciación celular. Cuando una célula se diferencia, se forman algunas sustancias o estructuras que no estaban previamente formadas y otras que lo estaban dejan de formarse. La diferenciación es a menudo parte del ciclo celular en que las células forman estructuras especializadas relacionadas con la reproducción, la dispersión o la supervivencia.
- Señalización. Las células responden a estímulos químicos y físicos tanto del medio externo como de su interior y, en el caso de células móviles, hacia determinados estímulos ambientales o en dirección opuesta mediante un proceso que se denomina quimiotaxis. Además, frecuentemente las células pueden interaccionar o comunicar con otras células, generalmente por medio de señales o mensajeros químicos, como hormonas, neurotransmisores, factores de crecimiento... en seres pluricelulares en complicados procesos de comunicación celular y transducción de señales.
- Evolución. A diferencia de las estructuras inanimadas, los organismos unicelulares y pluricelulares evolucionan. Esto significa que hay cambios hereditarios (que ocurren a baja frecuencia en todas las células de modo regular) que pueden influir en la adaptación global de la célula o del organismo superior de modo positivo o negativo. El resultado de la evolución es la selección de aquellos organismos mejor adaptados a vivir en un medio particular.
Las propiedades celulares no tienen por qué ser constantes a lo largo del desarrollo de un organismo: evidentemente, el patrón de expresión de los genes varía en respuesta a estímulos externos, además de factores endógenos.[18] Un aspecto importante a controlar es la pluripotencialidad, característica de algunas células que les permite dirigir su desarrollo hacia un abanico de posibles tipos celulares. En metazoos, la genética subyacente a la determinación del destino de una célula consiste en la expresión de determinados factores de transcripción específicos del linaje celular al cual va a pertenecer, así como a modificaciones epigenéticas. Además, la introducción de otro tipo de factores de transcripción mediante ingeniería genética en células somáticas basta para inducir la mencionada pluripotencialidad, luego este es uno de sus fundamentos moleculares.[19]
Tamaño, forma y función
El tamaño y la forma de las células depende de sus elementos más periféricos (por ejemplo, la pared, si la hubiere) y de su andamiaje interno (es decir, el citoesqueleto). Además, la competencia por el espacio tisular provoca una morfología característica: por ejemplo, las células vegetales, poliédricas in vivo, tienden a ser esféricas in vitro.[20] Incluso pueden existir parámetros químicos sencillos, como los gradientes de concentración de una sal, que determinen la aparición de una forma compleja.[21]
En cuanto al tamaño, la mayoría de las células son microscópicas, es decir, no son observables a simple vista. (un milímetro cúbico de sangre puede contener unos cinco millones de células),[15] A pesar de ser muy pequeñas, el tamaño de las células es extremadamente variable. La célula más pequeña observada, en condiciones normales, corresponde a Mycoplasma genitalium, de 0,2 μm, encontrándose cerca del límite teórico de 0,17 μm.[22] Existen bacterias con 1 y 2 μm de longitud. Las células humanas son muy variables: hematíes de 7 micras, hepatocitos con 20 micras, espermatozoides de 53 μm, óvulos de 150 μm e, incluso, algunas neuronas de en torno a un metro de longitud. En las células vegetales los granos de polen pueden llegar a medir de 200 a 300 μm.
Respecto a las células de mayor tamaño; por ejemplo, los xenofióforos,[23] son foraminíferos unicelulares que han desarrollado un gran tamaño, los cuales alcanzar tamaños macroscópicos (Syringammina fragilissima alcanza los 20 cm de diámetro).[24]
Para la viabilidad de la célula y su correcto funcionamiento siempre se debe tener en cuenta la relación superficie-volumen.[16] Puede aumentar considerablemente el volumen de la célula y no así su superficie de intercambio de membrana, lo que dificultaría el nivel y regulación de los intercambios de sustancias vitales para la célula.
Respecto de su forma, las células presentan una gran variabilidad, e, incluso, algunas no la poseen bien definida o permanente. Pueden ser: fusiformes (forma de huso), estrelladas, prismáticas, aplanadas, elípticas, globosas o redondeadas, etc. Algunas tienen una pared rígida y otras no, lo que les permite deformar la membrana y emitir prolongaciones citoplasmáticas (pseudópodos) para desplazarse o conseguir alimento. Hay células libres que no muestran esas estructuras de desplazamiento, pero poseen cilios o flagelos, que son estructuras derivadas de un orgánulo celular (el centrosoma) que dota a estas células de movimiento.[2] De este modo, existen multitud de tipos celulares, relacionados con la función que desempeñan; por ejemplo:
- Células contráctiles que suelen ser alargadas, como los miocitos esqueléticos.
- Células con finas prolongaciones, como las neuronas que transmiten el impulso nervioso.
- Células con microvellosidades o con pliegues, como las del intestino para ampliar la superficie de contacto y de intercambio de sustancias.
- Células cúbicas, prismáticas o aplanadas como las epiteliales que recubren superficies como las losas de un pavimento.
Estudio de las células
Los biólogos utilizan diversos instrumentos para lograr el conocimiento de las células. Obtienen información de sus formas, tamaños y componentes, que les sirve para comprender además las funciones que en ellas se realizan. Desde las primeras observaciones de células, hace más de 300 años, hasta la época actual, las técnicas y los aparatos se han ido perfeccionando, originándose una rama más de la Biología: la microscopía.
Dado el pequeño tamaño de la gran mayoría de las células, el uso del microscopio es de enorme valor en la investigación biológica. En la actualidad, los biólogos utilizan dos tipos básicos de microscopio: los ópticos y los electrónicos.
La célula procariota
Las células procariotas son pequeñas y menos complejas que las eucariotas. Contienen ribosomas pero carecen de sistemas de endomembranas (esto es, orgánulos delimitados por membranas biológicas, como puede ser el núcleo celular). Por ello poseen el material genético en el citosol. Sin embargo, existen excepciones: algunas bacterias fotosintéticas poseen sistemas de membranas internos.[25] También en el Filo Planctomycetes existen organismos como Pirellula que rodean su material genético mediante una membrana intracitoplasmática y Gemmata obscuriglobus que lo rodea con doble membrana. Esta última posee además otros compartimentos internos de membrana, posiblemente conectados con la membrana externa del nucleoide y con la membrana plasmática, que no está asociada a peptidoglucano.[26][27][28]
Estudios realizados en 2017, demuestran otra particularidad de Gemmata: presenta estructuras similares al poro nuclear, en la membrana que rodea su cuerpo nuclear.[29]
Por lo general podría decirse que los procariotas carecen de citoesqueleto. Sin embargo se ha observado que algunas bacterias, como Bacillus subtilis, poseen proteínas tales como MreB y mbl que actúan de un modo similar a la actina y son importantes en la morfología celular.[30] Fusinita van den Ent, en Nature, va más allá, afirmando que los citoesqueletos de actina y tubulina tienen origen procariótico.[31]
De gran diversidad, los procariotas sustentan un metabolismo extraordinariamente complejo, en algunos casos exclusivo de ciertos taxa, como algunos grupos de bacterias, lo que incide en su versatilidad ecológica.[13] Los procariotas se clasifican, según Carl Woese, en arqueas y bacterias.[32]
Arqueas
Las arqueas poseen un diámetro celular comprendido entre 0,1 y 15 μm, aunque las formas filamentosas pueden ser mayores por agregación de células. Presentan multitud de formas distintas: incluso las hay descritas cuadradas y planas.[33] Algunas arqueas tienen flagelos y son móviles.
Las arqueas, al igual que las bacterias, no tienen membranas internas que delimiten orgánulos. Como todos los organismos presentan ribosomas, pero a diferencia de los encontrados en las bacterias que son sensibles a ciertos agentes antimicrobianos, los de las arqueas, más cercanos a los eucariotas, no lo son. La membrana celular tiene una estructura similar a la de las demás células, pero su composición química es única, con enlaces tipo éter en sus lípidos.[34] Casi todas las arqueas poseen una pared celular (algunos Thermoplasma son la excepción) de composición característica, por ejemplo, no contienen peptidoglicano (mureína), propio de bacterias. No obstante pueden clasificarse bajo la tinción de Gram, de vital importancia en la taxonomía de bacterias; sin embargo, en arqueas, poseedoras de una estructura de pared en absoluto común a la bacteriana, dicha tinción es aplicable pero carece de valor taxonómico. El orden Methanobacteriales tiene una capa de pseudomureína, que provoca que dichas arqueas respondan como positivas a la tinción de Gram.[35][36] [37]
Como en casi todos los procariotas, las células de las arqueas carecen de núcleo, y presentan un solo cromosoma circular. Existen elementos extracromosómicos, tales como plásmidos. Sus genomas son de pequeño tamaño, sobre 2-4 millones de pares de bases. También es característica la presencia de ARN polimerasas de constitución compleja y un gran número de nucleótidos modificados en los ácidos ribonucleicos ribosomales. Por otra parte, su ADN se empaqueta en forma de nucleosomas, como en los eucariotas, gracias a proteínas semejantes a las histonas y algunos genes poseen intrones.[38] Pueden reproducirse por fisión binaria o múltiple, fragmentación o gemación.
Bacterias

Las bacterias son organismos relativamente sencillos, de dimensiones muy reducidas, de apenas unas micras en la mayoría de los casos. Como otros procariotas, carecen de un núcleo delimitado por una membrana, aunque presentan un nucleoide, una estructura elemental que contiene una gran molécula generalmente circular de ADN.[17][39] Carecen de núcleo celular y demás orgánulos delimitados por membranas biológicas.[40] En el citoplasma se pueden apreciar plásmidos, pequeñas moléculas circulares de ADN que coexisten con el nucleoide y que contienen genes: son comúnmente usados por las bacterias en la parasexualidad (reproducción sexual bacteriana). El citoplasma también contiene ribosomas y diversos tipos de gránulos. En algunos casos, puede haber estructuras compuestas por membranas, generalmente relacionadas con la fotosíntesis.[9]
Poseen una membrana celular compuesta de lípidos, en forma de una bicapa y sobre ella se encuentra una cubierta en la que existe un polisacárido complejo denominado peptidoglicano; dependiendo de su estructura y subsecuente su respuesta a la tinción de Gram, se clasifica a las bacterias en Gram positivas y Gram negativas. El espacio comprendido entre la membrana celular y la pared celular (o la membrana externa, si esta existe) se denomina espacio periplásmico. Algunas bacterias presentan una cápsula. Otras son capaces de generar endosporas (estadios latentes capaces de resistir condiciones extremas) en algún momento de su ciclo vital. Entre las formaciones exteriores propias de la célula bacteriana destacan los flagelos (de estructura completamente distinta a la de los flagelos eucariotas) y los pili (estructuras de adherencia y relacionadas con la parasexualidad).[9]
La mayoría de las bacterias disponen de un único cromosoma circular y suelen poseer elementos genéticos adicionales, como distintos tipos de plásmidos. Su reproducción, binaria y muy eficiente en el tiempo, permite la rápida expansión de sus poblaciones, generándose un gran número de células que son virtualmente clones, esto es, idénticas entre sí.[38]
La célula eucariota
Las células eucariotas son el exponente de la complejidad celular actual.[15] Presentan una estructura básica relativamente estable caracterizada por la presencia de distintos tipos de orgánulos intracitoplasmáticos especializados, entre los cuales destaca el núcleo, que alberga el material genético. Especialmente en los organismos pluricelulares, las células pueden alcanzar un alto grado de especialización. Dicha especialización o diferenciación es tal que, en algunos casos, compromete la propia viabilidad del tipo celular en aislamiento. Así, por ejemplo, las neuronas dependen para su supervivencia de las células gliales.[13]
Por otro lado, la estructura de la célula varía dependiendo de la situación taxonómica del ser vivo: de este modo, las células vegetales difieren de las animales, así como de las de los hongos. Por ejemplo, las células animales carecen de pared celular, son muy variables, no tiene plastos, puede tener vacuolas pero no son muy grandes y presentan centríolos (que son agregados de microtúbulos cilíndricos que contribuyen a la formación de los cilios y los flagelos y facilitan la división celular). Las células de los vegetales, por su lado, presentan una pared celular compuesta principalmente de celulosa, disponen de plastos como cloroplastos (orgánulo capaz de realizar la fotosíntesis), cromoplastos (orgánulos que acumulan pigmentos) o leucoplastos (orgánulos que acumulan el almidón fabricado en la fotosíntesis), poseen vacuolas de gran tamaño que acumulan sustancias de reserva o de desecho producidas por la célula y finalmente cuentan también con plasmodesmos, que son conexiones citoplasmáticas que permiten la circulación directa de las sustancias del citoplasma de una célula a otra, con continuidad de sus membranas plasmáticas.[41]


Compartimentos
Las células son entes dinámicos, con un metabolismo celular interno de gran actividad cuya estructura es un flujo entre rutas anastomosadas. Un fenómeno observado en todos los tipos celulares es la compartimentalización, que consiste en una heterogeneidad que da lugar a entornos más o menos definidos (rodeados o no mediante membranas biológicas) en las cuales existe un microentorno que aglutina a los elementos implicados en una ruta biológica.[42] Esta compartimentalización alcanza su máximo exponente en las células eucariotas, las cuales están formadas por diferentes estructuras y orgánulos que desarrollan funciones específicas, lo que supone un método de especialización espacial y temporal.[2] No obstante, células más sencillas, como los procariotas, ya poseen especializaciones semejantes.[43]
Membrana plasmática y superficie celular
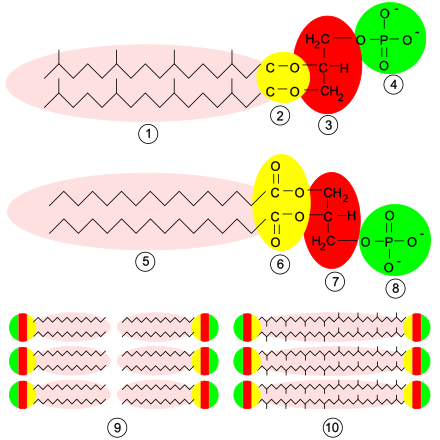
La composición de la membrana plasmática varía entre células dependiendo de la función o del tejido en la que se encuentre, pero posee elementos comunes. Está compuesta por una doble capa de fosfolípidos, por proteínas unidas no covalentemente a esa bicapa, y por glúcidos unidos covalentemente a lípidos o proteínas. Generalmente, las moléculas más numerosas son las de lípidos; sin embargo, las proteínas, debido a su mayor masa molecular, representan aproximadamente el 50 % de la masa de la membrana.[42]
Un modelo que explica el funcionamiento de la membrana plasmática es el modelo del mosaico fluido, de J. S. Singer y Garth Nicolson (1972), que desarrolla un concepto de unidad termodinámica basada en las interacciones hidrófobas entre moléculas y otro tipo de enlaces no covalentes.[44]

Dicha estructura de membrana sustenta un complejo mecanismo de transporte, que posibilita un fluido intercambio de masa y energía entre el entorno intracelular y el externo.[42] Además, la posibilidad de transporte e interacción entre moléculas de células aledañas o de una célula con su entorno faculta a estas poder comunicarse químicamente, esto es, permite la señalización celular. Neurotransmisores, hormonas, mediadores químicos locales afectan a células concretas modificando el patrón de expresión génica mediante mecanismos de transducción de señal.[45]
Sobre la bicapa lipídica, independientemente de la presencia o no de una pared celular, existe una matriz que puede variar, de poco conspicua, como en los epitelios, a muy extensa, como en el tejido conjuntivo. Dicha matriz, denominada glucocalix (glicocáliz), rica en líquido tisular, glucoproteínas, proteoglicanos y fibras, también interviene en la generación de estructuras y funciones emergentes, derivadas de las interacciones célula-célula.[13]
Estructura y expresión génica

Las células eucariotas poseen su material genético en, generalmente, un solo núcleo celular, delimitado por una envoltura consistente en dos bicapas lipídicas atravesadas por numerosos poros nucleares y en continuidad con el retículo endoplasmático. En su interior, se encuentra el material genético, el ADN, observable, en las células en interfase, como cromatina de distribución heterogénea. A esta cromatina se encuentran asociadas multitud de proteínas, entre las cuales destacan las histonas, así como ARN, otro ácido nucleico.[46]
Dicho material genético se encuentra inmerso en una actividad continua de regulación de la expresión génica; las ARN polimerasas transcriben ARN mensajero continuamente, que, exportado al citosol, es traducido a proteína, de acuerdo a las necesidades fisiológicas. Asimismo, dependiendo del momento del ciclo celular, dicho ADN puede entrar en replicación, como paso previo a la mitosis.[38] No obstante, las células eucarióticas poseen material genético extranuclear: concretamente, en mitocondrias y plastos, si los hubiere; estos orgánulos conservan una independencia genética parcial del genoma nuclear.[47][48]
Síntesis y degradación de macromoléculas
Dentro del citosol, esto es, la matriz acuosa que alberga a los orgánulos y demás estructuras celulares, se encuentran inmersos multitud de tipos de maquinaria de metabolismo celular: orgánulos, inclusiones, elementos del citoesqueleto, enzimas... De hecho, estas últimas corresponden al 20 % de las enzimas totales de la célula.[13]


- Ribosoma: Los ribosomas, visibles al microscopio electrónico como partículas esféricas,[49] son complejos supramoleculares encargados de ensamblar proteínas a partir de la información genética que les llega del ADN transcrita en forma de ARN mensajero. Elaborados en el núcleo, desempeñan su función de síntesis de proteínas en el citoplasma. Están formados por ARN ribosómico y por diversos tipos de proteínas. Estructuralmente, tienen dos subunidades. En las células, estos orgánulos aparecen en diferentes estados de disociación. Cuando están completos, pueden estar aislados o formando grupos (polisomas). También pueden aparecer asociados al retículo endoplasmático rugoso o a la envoltura nuclear.[38]
- Retículo endoplasmático: El retículo endoplasmático es orgánulo vesicular interconectado que forma cisternas, tubos aplanados y sáculos comunicados entre sí. Intervienen en funciones relacionadas con la síntesis proteica, glicosilación de proteínas, metabolismo de lípidos y algunos esteroides, detoxificación, así como el tráfico de vesículas. En células especializadas, como las miofibrillas o células musculares, se diferencia en el retículo sarcoplásmico, orgánulo decisivo para que se produzca la contracción muscular.[15]
- Aparato de Golgi: El aparato de Golgi es un orgánulo formado por apilamientos de sáculos denominados dictiosomas, si bien, como ente dinámico, estos pueden interpretarse como estructuras puntuales fruto de la coalescencia de vesículas.[50][51] Recibe las vesículas del retículo endoplasmático rugoso que han de seguir siendo procesadas. Dentro de las funciones que posee el aparato de Golgi se encuentran la glicosilación de proteínas, selección, destinación, glicosilación de lípidos y la síntesis de polisacáridos de la matriz extracelular. Posee tres compartimientos; uno proximal al retículo endoplasmático, denominado «compartimento cis», donde se produce la fosforilación de las manosas de las enzimas que han de dirigirse al lisosoma; el «compartimento intermedio», con abundantes manosidasas y N-acetil-glucosamina transferasas; y el «compartimento o red trans», el más distal, donde se transfieren residuos de galactosa y ácido siálico, y del que emergen las vesículas con los diversos destinos celulares.[13]
- Lisosoma: Los lisosomas son orgánulos que albergan multitud de enzimas hidrolíticas. De morfología muy variable, no se ha demostrado su existencia en células vegetales.[13] Una característica que agrupa a todos los lisosomas es la posesión de hidrolasas ácidas: proteasas, nucleasas, glucosidasas, lisozima, arilsulfatasas, lipasas, fosfolipasas y fosfatasas. Procede de la fusión de vesículas procedentes del aparato de Golgi, que, a su vez, se fusionan en un tipo de orgánulo denominado endosoma temprano, el cual, al acidificarse y ganar en enzimas hidrolíticos, pasa a convertirse en el lisosoma funcional. Sus funciones abarcan desde la degradación de macromoléculas endógenas o procedentes de la fagocitosis a la intervención en procesos de apoptosis.[52]

- Vacuola vegetal: Las vacuolas vegetales, numerosas y pequeñas en células meristemáticas y escasas y grandes en células diferenciadas, son orgánulos exclusivos de los representantes del mundo vegetal. Inmersas en el citosol, están delimitadas por el tonoplasto, una membrana lipídica. Sus funciones son: facilitar el intercambio con el medio externo, mantener la turgencia celular, la digestión celular y la acumulación de sustancias de reserva y subproductos del metabolismo.[41]
- Inclusión citoplasmática: Las inclusiones son acúmulos nunca delimitados por membrana de sustancias de diversa índole, tanto en células vegetales como animales. Típicamente se trata de sustancias de reserva que se conservan como acervo metabólico: almidón, glucógeno, triglicéridos, proteínas... aunque también existen de pigmentos.[13]
Conversión energética
El metabolismo celular está basado en la transformación de unas sustancias químicas, denominadas metabolitos, en otras; dichas reacciones químicas transcurren catalizadas mediante enzimas. Si bien buena parte del metabolismo sucede en el citosol, como la glucólisis, existen procesos específicos de orgánulos.[45]

- Mitocondria: Las mitocondrias son orgánulos de aspecto, número y tamaño variable que intervienen en el ciclo de Krebs, fosforilación oxidativa y en la cadena de transporte de electrones de la respiración. Presentan una doble membrana, externa e interna, que dejan entre ellas un espacio perimitocondrial; la membrana interna, plegada en crestas hacia el interior de la matriz mitocondrial, posee una gran superficie. En su interior posee generalmente una sola molécula de ADN, el genoma mitocondrial, típicamente circular, así como ribosomas más semejantes a los bacterianos que a los eucariotas.[13] Según la teoría endosimbiótica, se asume que la primera protomitocondria era un tipo de proteobacteria.[53]

- Cloroplasto: Los cloroplastos son los orgánulos celulares que en los organismos eucariotas fotosintéticos se ocupan de la fotosíntesis. Están limitados por una envoltura formada por dos membranas concéntricas y contienen vesículas, los tilacoides, donde se encuentran organizados los pigmentos y demás moléculas implicadas en la conversión de la energía lumínica en energía química. Además de esta función, los plastidios intervienen en el metabolismo intermedio, produciendo energía y poder reductor, sintetizando bases púricas y pirimidínicas, algunos aminoácidos y todos los ácidos grasos. Además, en su interior es común la acumulación de sustancias de reserva, como el almidón.[13] Se considera que poseen analogía con las cianobacterias.[54]

- Peroxisoma: Los peroxisomas son orgánulos muy comunes en forma de vesículas que contienen abundantes enzimas de tipo oxidasa y catalasa; de tan abundantes, es común que cristalicen en su interior. Estas enzimas cumplen funciones de detoxificación celular. Otras funciones de los peroxisomas son: las oxidaciones flavínicas generales, el catabolismo de las purinas, la beta-oxidación de los ácidos grasos, el ciclo del glioxilato, el metabolismo del ácido glicólico y la detoxificación en general.[13] Se forman de vesículas procedentes del retículo endoplasmático.[55]
Citoesqueleto
Las células poseen un andamiaje que permite el mantenimiento de su forma y estructura, pero más aún, este es un sistema dinámico que interactúa con el resto de componentes celulares generando un alto grado de orden interno. Dicho andamiaje está formado por una serie de proteínas que se agrupan dando lugar a estructuras filamentosas que, mediante otras proteínas, interactúan entre ellas dando lugar a una especie de retículo. El mencionado andamiaje recibe el nombre de citoesqueleto, y sus elementos mayoritarios son: los microtúbulos, los microfilamentos y los filamentos intermedios.[2][nota 2][56][57]
- Microfilamentos: Los microfilamentos o filamentos de actina están formados por una proteína globular, la actina, que puede polimerizar dando lugar a estructuras filiformes. Dicha actina se expresa en todas las células del cuerpo y especialmente en las musculares ya que está implicada en la contracción muscular, por interacción con la miosina. Además, posee lugares de unión a ATP, lo que dota a sus filamentos de polaridad.[58] Puede encontrarse en forma libre o polimerizarse en microfilamentos, que son esenciales para funciones celulares tan importantes como la movilidad y la contracción de la célula durante la división celular.[50]

- Microtúbulos: Los microtúbulos son estructuras tubulares de 25 nm de diámetro exterior y unos 12 nm de diámetro interior, con longitudes que varían entre unos pocos nanómetros a micrómetros, que se originan en los centros organizadores de microtúbulos y que se extienden a lo largo de todo el citoplasma. Se hallan en las células eucariotas y están formadas por la polimerización de un dímero de dos proteínas globulares, la alfa y la beta tubulina. Las tubulinas poseen capacidad de unir GTP.[2][50] Los microtúbulos intervienen en diversos procesos celulares que involucran desplazamiento de vesículas de secreción, movimiento de orgánulos, transporte intracelular de sustancias, así como en la división celular (mitosis y meiosis) y que, junto con los microfilamentos y los filamentos intermedios, forman el citoesqueleto. Además, constituyen la estructura interna de los cilios y los flagelos.[2][50]
- Filamentos intermedios: Los filamentos intermedios son componentes del citoesqueleto. Formados por agrupaciones de proteínas fibrosas, su nombre deriva de su diámetro, de 10 nm, menor que el de los microtúbulos, de 24 nm, pero mayor que el de los microfilamentos, de 7 nm. Son ubicuos en las células animales, y no existen en plantas ni hongos. Forman un grupo heterogéneo, clasificado en cinco familias: las queratinas, en células epiteliales; los neurofilamentos, en neuronas; los gliofilamentos, en células gliales; la desmina, en músculo liso y estriado; y la vimentina, en células derivadas del mesénquima.[13]

- Centríolos: Los centríolos son una pareja de estructuras que forman parte del citoesqueleto de células animales. Semejantes a cilindros huecos, están rodeados de un material proteico denso llamado material pericentriolar; todos ellos forman el centrosoma o centro organizador de microtúbulos que permiten la polimerización de microtúbulos de dímeros de tubulina que forman parte del citoesqueleto. Los centríolos se posicionan perpendicularmente entre sí. Sus funciones son participar en la mitosis, durante la cual generan el huso acromático, y en la citocinesis,[59] así como, se postula, intervenir en la nucleación de microtúbulos.[60][61]
- Cilios y flagelos: Se trata de especializaciones de la superficie celular con motilidad; con una estructura basada en agrupaciones de microtúbulos, ambos se diferencian en la mayor longitud y menor número de los flagelos, y en la mayor variabilidad de la estructura molecular de estos últimos.[13]
Ciclo vital

El ciclo celular es el proceso ordenado y repetitivo en el tiempo mediante el cual una célula madre crece y se divide en dos células hijas. Las células que no se están dividiendo se encuentran en una fase conocida como G0, paralela al ciclo. La regulación del ciclo celular es esencial para el correcto funcionamiento de las células sanas, está claramente estructurado en fases[50]
- El estado de no división o interfase. La célula realiza sus funciones específicas y, si está destinada a avanzar a la división celular, comienza por realizar la duplicación de su ADN.
- El estado de división, llamado fase M, situación que comprende la mitosis y citocinesis. En algunas células la citocinesis no se produce, obteniéndose como resultado de la división una masa celular plurinucleada denominada plasmodio.[nota 3]
A diferencia de lo que sucede en la mitosis, donde la dotación genética se mantiene, existe una variante de la división celular, propia de las células de la línea germinal, denominada meiosis. En ella, se reduce la dotación genética diploide, común a todas las células somáticas del organismo, a una haploide, esto es, con una sola copia del genoma. De este modo, la fusión, durante la fecundación, de dos gametos haploides procedentes de dos parentales distintos da como resultado un zigoto, un nuevo individuo, diploide, equivalente en dotación genética a sus padres.[62]
- La interfase consta de tres estadios claramente definidos.[2][50]
- Fase G1: es la primera fase del ciclo celular, en la que existe crecimiento celular con síntesis de proteínas y de ARN. Es el período que trascurre entre el fin de una mitosis y el inicio de la síntesis de ADN. En él la célula dobla su tamaño y masa debido a la continua síntesis de todos sus componentes, como resultado de la expresión de los genes que codifican las proteínas responsables de su fenotipo particular.
- Fase S: es la segunda fase del ciclo, en la que se produce la replicación o síntesis del ADN. Como resultado cada cromosoma se duplica y queda formado por dos cromátidas idénticas. Con la duplicación del ADN, el núcleo contiene el doble de proteínas nucleares y de ADN que al principio.
- Fase G2: es la segunda fase de crecimiento del ciclo celular en la que continúa la síntesis de proteínas y ARN. Al final de este período se observa al microscopio cambios en la estructura celular, que indican el principio de la división celular. Termina cuando los cromosomas empiezan a condensarse al inicio de la mitosis.
- La fase M es la fase de la división celular en la cual una célula progenitora se divide en dos células hijas idénticas entre sí y a la madre. Esta fase incluye la mitosis, a su vez dividida en: profase, metafase, anafase, telofase; y la citocinesis, que se inicia ya en la telofase mitótica.
La incorrecta regulación del ciclo celular puede conducir a la aparición de células precancerígenas que, si no son inducidas al suicidio mediante apoptosis, puede dar lugar a la aparición de cáncer. Los fallos conducentes a dicha desregulación están relacionados con la genética celular: lo más común son las alteraciones en oncogenes, genes supresores de tumores y genes de reparación del ADN.[63]
Origen
Origen de la primera célula
La aparición de la vida, y, por ello, de la célula, probablemente se inició gracias a la transformación de moléculas inorgánicas en orgánicas bajo unas condiciones ambientales adecuadas, produciéndose más adelante la interacción de estas biomoléculas generando entes de mayor complejidad. El experimento de Miller y Urey, realizado en 1953, demostró que una mezcla de compuestos orgánicos sencillos puede transformarse en algunos aminoácidos, glúcidos y lípidos (componentes todos ellos de la materia viva) bajo unas condiciones ambientales que simulan las presentes hipotéticamente en la Tierra primigenia (en torno al eón Arcaico).[64]
Se postula que dichos componentes orgánicos se agruparon generando estructuras complejas, los coacervados de Oparin, aún acelulares que, en cuanto alcanzaron la capacidad de autoorganizarse y perpetuarse, dieron lugar a un tipo de célula primitiva, el progenote de Carl Woese, antecesor de los tipos celulares actuales.[32] Una vez se diversificó este grupo celular, dando lugar a las variantes procariotas, arqueas y bacterias, pudieron aparecer nuevos tipos de células, más complejos, por endosimbiosis, esto es, captación permanente de unos tipos celulares en otros sin una pérdida total de autonomía de aquellos.[65] De este modo, algunos autores describen un modelo en el cual la primera célula eucariota surgió por introducción de una arquea en el interior de una bacteria, dando lugar esta primera a un primitivo núcleo celular.[66] No obstante, la imposibilidad de que una bacteria pueda efectuar una fagocitosis y, por ello, captar a otro tipo de célula, dio lugar a otra hipótesis, que sugiere que fue una célula denominada cronocito la que fagocitó a una bacteria y a una arquea, dando lugar al primer organismo eucariota. De este modo, y mediante un análisis de secuencias a nivel genómico de organismos modelo eucariotas, se ha conseguido describir a este cronocito original como un organismo con citoesqueleto y membrana plasmática, lo cual sustenta su capacidad fagocítica, y cuyo material genético era el ARN, lo que puede explicar, si la arquea fagocitada lo poseía en el ADN, la separación espacial en los eucariotas actuales entre la transcripción (nuclear), y la traducción (citoplasmática).[67]
Una dificultad adicional es el hecho de que no se han encontrado organismos eucariotas primitivamente amitocondriados como exige la hipótesis endosimbionte. Además, el equipo de María Rivera, de la Universidad de California, comparando genomas completos de todos los dominios de la vida ha encontrado evidencias de que los eucariotas contienen dos genomas diferentes, uno más semejante a bacterias y otro a arqueas, apuntando en este último caso semejanzas a los metanógenos, en particular en el caso de las histonas.[68][69] Esto llevó a Bill Martin y Miklós Müller a plantear la hipótesis de que la célula eucariota surgiera no por endosimbiosis, sino por fusión quimérica y acoplamiento metabólico de un metanógeno y una α-proteobacteria simbiontes a través del hidrógeno (hipótesis del hidrógeno).[70] Esta hipótesis atrae hoy en día posiciones muy encontradas, con detractores como Christian de Duve.[71]
Harold Morowitz, un físico de la Universidad Yale, ha calculado que las probabilidades de obtener la bacteria viva más sencilla mediante cambios al azar es de 1 sobre 1 seguido por 100 000 000 000 ceros. «Este número es tan grande —dijo Robert Shapiro— que para escribirlo en forma convencional necesitaríamos varios centenares de miles de libros en blanco». Presenta la acusación de que los científicos que han abrazado la evolución química de la vida pasan por alto la evidencia aumentante y «han optado por aceptarla como verdad que no puede ser cuestionada, consagrándola así como mitología».[72]
Origen de la célula eucariota

Las células eucariotas se crearon hace unos 2.200 millones de años en un proceso llamado eucariogénesis.[74] Se acepta ampliamente que esto implicó una simbiogénesis, en la que arqueas y bacterias se unieron para crear el primer ancestro común eucariota.[75] Esta célula tenía un nuevo nivel de complejidad y capacidad, con un núcleo[76][77] y mitocondrias facultativamente aeróbicas.[73] Evolucionó hace unos 2 mil millones de años hasta convertirse en una población de organismos unicelulares que incluía al último ancestro común eucariota, acumulando capacidades a lo largo del camino, aunque la secuencia de los pasos involucrados ha sido cuestionada y es posible que no haya comenzado con la simbiogénesis. Presentaba al menos un centriolo y cilio, sexo (meiosis y singamia), peroxisomas y un quiste latente con una pared celular de quitina y/o celulosa.[78][79] A su vez, el último ancestro común eucariota dio origen al grupo terminal de los eucariotas, que contiene los ancestros de animales, hongos, plantas y una amplia gama de organismos unicelulares.[80][81][82] Las células vegetales se crearon hace unos 1.600 millones de años con un segundo episodio de simbiogénesis al que se añadieron cloroplastos, derivados de cianobacterias.[83][84][85][73]
Véase también
Notas
- ↑ Algunos autores consideran que la cifra propuesta por Schopf es un desacierto. Por ejemplo, destacan que los presuntos microfósiles encontrados en rocas de más de 2,7 Ga de antigüedad como estromatoloides, ondulaciones, dendritas, efectos de «cercos de café», filoides, rebordes de cristales poligonales y esferulitas podrían ser en realidad estructuras auto-organizadas que tuvieron lugar en un momento en que los macrociclos geoquímicos globales tenían mucha más importancia, la corteza continental era menor y la actividad magmática e hidrotermal tenía una importancia capital. Según este estudio no se puede atribuir estas estructuras a la actividad biológica (endolitos) con toda seguridad.
- ↑ Cabe destacar que el citoesqueleto no es un elemento exclusivo del tipo celular eucariota: hay homólogos bacterianos para sus proteínas de mayor relevancia. De este modo, en procariotas el citoesqueleto también contribuye a la división celular, determinación de la forma y polaridad, etc.
- ↑ A veces se denomina incorrectamente sincitio a la mencionada masa pluricelular, si bien el término solo debe emplearse para describir a las células que proceden de la fusión de células mononucleadas y no a aquellas producto de la ausencia de citocinesis.
Referencias
- ↑ Entrada "célula" en el DRAE
- ↑ a b c d e f g Alberts et al (2004). Biología molecular de la célula. Barcelona: Omega. ISBN 54-282-1351-8.
- ↑ Aréchiga, H. (1996). Siglo XXI, ed. Los fenómenos fundamentales de la vida. p. 178. ISBN 9789682320194.
- ↑ Maton, Anthea; Hopkins, Jean Johnson, Susan LaHart, David Quon Warner, Maryanna Wright, Jill D (1997). Cells Building Blocks of Life. New Jersey: Prentice Hall. ISBN 0-13-423476-6.
- ↑ J William Schopf. New evidence of the antiquity of life. Origins of Life and Evolution of Biospheres. Springer Netherlands. ISSN 0169-6149
- ↑ M Brasier, N McLoughlin, O Green, D Wacey. A fresh look at the fossil evidence for early Archaean cellular life Philosophical Transactions of the Royal Society B, 2006 - The Royal Society
- ↑ Wacey, David; Matt R. Kilburn, Martin Saunders, John Cliff, Martin D. Brasier (2011-08). «Microfossils of sulphur-metabolizing cells in 3.4-billion-year-old rocks of Western Australia». Nature Geoscience. ISSN 1752-0894. doi:10.1038/ngeo1238. Consultado el 23 de agosto de 2011.
- ↑ Bechtel, William (2005). Discovering Cell Mechanisms: The Creation of Modern Cell. Cambridge University Press. ISBN 052181247X.
- ↑ a b c d Prescott, L.M. (1999). Microbiología. McGraw-Hill Interamericana de España, S.A.U. ISBN 84-486-0261-7.
- ↑ Janssen's Microscope Optical microscopy primer: museum of microscopy.
- ↑ Extracto de la descripción por Hooke (Universidad de Berkeley)
[...]I could exceedingly plainly perceive it to be all perforated and porous, much like a Honey-comb, but that the pores of it were not regular [..] these pores, or cells, [..] were indeed the first microscopical pores I ever saw, and perhaps, that were ever seen, for I had not met with any Writer or Person, that had made any mention of them before this. [...]Hooke
- ↑ Lynn Margulis (1981): Symbiosis in Cell Evolution. WH Freeman & Company.
- ↑ a b c d e f g h i j k l m Paniagua, R.; Nistal, M.; Sesma, P.; Álvarez-Uría, M.; Fraile, B.; Anadón, R. y José Sáez, F. (2002). Citología e histología vegetal y animal. McGraw-Hill Interamericana de España, S.A.U. ISBN 84-486-0436-9.
- ↑ Tavassoli (1980). The cell theory: a foundation to the edifice of biology. American Journal of Patholology January; 98(1): 44.[1]
- ↑ a b c d Randall, D.; Burggren, W. et French, K. (1998). Eckert Fisiología animal (4.ª edición). ISBN 84-486-0200-5.
- ↑ a b Cromer, A.H. (1996). Física para ciencias de la vida. Reverté ediciones. ISBN para España 84-291-1808-X.
- ↑ a b Griffiths, J. F. A. et al. (2002). Genética. McGraw-Hill Interamericana. ISBN 84-486-0368-0.
- ↑ Sally A. Moody, ed. (2007). Principles of Developmental Genetics (1 edición). Burlington, USA: Elsevier. ISBN 978-0-12-369548-2.
- ↑ Welstead, GG, Schorderet, P and Boyer, LA.The reprogramming language of pluripotency. Curr Opin Genet Dev. 2008 Apr;18(2):123-9
- ↑ Azcón-Bieto,J y Talón, M. (2000). Fundamentos de Fisiología Vegetal. Mc Graw Hill Interamericana de España SAU. ISBN 84-486-0258-7.
- ↑ Brian Goodwin. The Cytoskeleton of the algae, 1989
- ↑ Mike Conrad. «What is the smallest living thing?». Consultado el 19 de junio de 2008.
- ↑ Zhuravlev, AY 1993, Were Ediacaran Vendobionta multicellulars? Neues Jahrb. Geol. Palëontol. 190ː 299-314.
- ↑ Marshall M (3 de febrero de 2010). «Zoologger: 'Living beach ball' is giant single cell». New Scientist.
- ↑ J. Oelze and G. Drews Membranes of photosynthetic bacteri1 Archivado el 26 de septiembre de 2008 en Wayback Machine. Biochimica et Biophysica Acta (BBA) - Reviews on Biomembranes Volume 265, Issue 2, 18 April 1972, Pages 209-239
- ↑ Prescott, LM; Harley, JP y Klein, DA: (1999). Microbiología. McGraw Hill-interamericana. ISBN 084-486-0261-7.
- ↑ «Gemmata». Consultado el 19-6 de 2008.
- ↑ «Pirellula». Consultado el 19-6.
- ↑ Sagulenko E, Nouwens A, Webb RI, Green K, Yee B, Morgan G, et al. (2017). «uclear Pore-Like Structures in a Compartmentalized Bacterium». PLoS ONE 12 (2): e0169432. ISSN 1932-6203. doi:10.1371/journal.pone.0169432.
- ↑ Jones LJ, Carballido-López R, Errington J (2001). «Control of cell shape in bacteria: helical, actin-like filaments in Bacillus subtilis». Cell 104 (6). PMID 11290328.
- ↑ van den Ent F, Amos LA, Löwe J (2001). «Prokaryotic origin of the actin cytoskeleton». Nature 413 (6851). PMID 11544518.
- ↑ a b Woese C, Kandler O, Wheelis M (1990). «Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya.». Proc Natl Acad Sci U S A 87 (12): 4576-9. PMID 2112744.
- ↑ Burns DG, Camakaris HM, Janssen PH, Dyall-Smith ML. (2004). «Cultivation of Walsby's square haloarchaeon.». FEMS Microbiol Lett. 238 (2): 469-73. PMID 15358434.
- ↑ Yosuke Koga et Hiroyuki Morii. Recent Advances in Structural Research on Ether Lipids from Archaea Including Comparative and Physiological Aspects. Bioscience, Biotechnology, and Biochemistry Vol. 69 (2005), No. 11 pp.2019-2034
- ↑ TJ Beveridge et S Schutze-Lam (2002). «The structure of bacterial surfaces and its influence on stainability». Journal of histotechnology 25 (1): 55-60.
- ↑ TJ Beveridge y S Schultze-Lam (1996). «The response of selected members of the archaea to the gram stain». Microbiology 142: 2887-2895.
- ↑ «Curso de mirobiología general». Archivado desde el original el 8 de julio de 2007.
- ↑ a b c d Watson, J, D.; Baker, T. A.; Bell, S. P.; Gann, A.; Levine, M. et Losick, R (2004). Molecular Biology of the Gene (Fifth edition edición). San Francisco: Benjamin Cummings. ISBN 0-321-22368-3.
- ↑ Thanbichler M, Wang S, Shapiro L (2005). «The bacterial nucleoid: a highly organized and dynamic structure». J Cell Biochem 96 (3): 506-21. PMID 15988757.
- ↑ Berg J., Tymoczko J. and Stryer L. (2002) Biochemistry. W. H. Freeman and Company ISBN 0-7167-4955-6
- ↑ a b Taiz, Lincoln; Zeiger, Eduardo (2006). Plant Physiology (4.ª edición). Sunderland, USA: Sinauer Associates, Inc. ISBN 978-0-87893-856-8.
- ↑ a b c Mathews, C. K.; Van Holde, K.E et Ahern, K.G (2003). «6». Bioquímica (3 edición). pp. 204 y ss. ISBN 84-7892-053-2.
- ↑ Germaine Cohen-Bazire, Norbert Pfennig and Riyo Kunisawa The fine structure of green bacteria The Journal of Cell Biology, Vol 22, 207-225, 1964
- ↑ The fluid mosaic model of the structure of cell membranes by S. J. Singer and G. L. Nicolson in Science (1972) Volume 175, pages 720-731.
- ↑ a b Lehninger, Albert (1993). Principles of Biochemistry, 2nd Ed. Worth Publishers. ISBN 0-87901-711-2.
- ↑ D L Spector. Macromolecular Domains within the Cell Nucleus. Annual Review of Cell Biology. Vol. 9: 265-315 (doi:10.1146/annurev.cb.09.110193.001405)
- ↑ Robert N. Lightowlers, Patrick F. Chinnery, Douglass M. Turnbulland Neil Howell. Mammalian mitochondrial genetics: heredity, heteroplasmy and disease. Trends in Genetics. Volume 13, Issue 11, November 1997, Pages 450-455
- ↑ Shusei Sato, Yasukazu Nakamura, Takakazu Kaneko, Erika Asamizu and Satoshi Tabata. Complete Structure of the Chloroplast Genome of Arabidopsis thaliana. DNA Research 1999 6(5):283-290; doi:10.1093/dnares/6.5.283
- ↑ G.E. Palade. (1955) «A small particulate component of the cytoplasm». J Biophys Biochem Cytol. Jan;1(1): pp. 59-68. PMID 14381428
- ↑ a b c d e f Lodish et al. (2005). Biología celular y molecular. Buenos Aires: Médica Panamericana. ISBN 950-06-1974-3.
- ↑ Glick, B.S. and Malhotra, V. (1998). «The curious status of the Golgi apparatus». Cell 95: 883-889.
- ↑ Mader, Sylvia. (2007). Biology 9th ed. McGraw Hill. New York. ISBN 978-0-07-246463-4
- ↑ Futuyma DJ (2005). «On Darwin's Shoulders». Natural History 114 (9): 64-68.
- ↑ Mereschkowsky C (1905). «Über Natur und Ursprung der Chromatophoren im Pflanzenreiche». Biol Centralbl 25: 593-604.
- ↑ Hoepfner D, Schildknegt D, Braakman I, Philippsen P, Tabak HF (2005). «Contribution of the endoplasmic reticulum to peroxisome formation». Cell 122 (1): 85-95. PMID 16009135. doi:10.1016/j.cell.2005.04.025.
- ↑ Shih YL, Rothfield L (2006). «The bacterial cytoskeleton». Microbiol. Mol. Biol. Rev. 70 (3): 729-54. PMID 16959967. doi:10.1128/MMBR.00017-06.
- ↑ Michie KA, Löwe J (2006). «Dynamic filaments of the bacterial cytoskeleton». Annu. Rev. Biochem. 75: 467-92. PMID 16756499. doi:10.1146/annurev.biochem.75.103004.142452. Archivado desde el original el 26 de marzo de 2009. Consultado el 17 de mayo de 2013.
- ↑ Straub, F.B. and Feuer, G. (1950) Adenosinetriphosphate the functional group of actin. Biochim. Biophys. Acta. 4, 455-470 Entrez PubMed 2673365
- ↑ Jeffrey L. Salisbury, Kelly M. Suino, Robert Busby, Margaret Springett; Centrin-2 Is Required for Centriole Duplication in Mammalian Cells Archivado el 18 de mayo de 2008 en Wayback Machine.; Current Biology, Volume 12, Issue 15, 6 August 2002, Pages 1287-1292; doi:10.1016/S0960-9822(02)01019-9
- ↑ Jessica L. Feldman, Stefan Geimer, Wallace F. Marshall; The Mother Centriole Plays an Instructive Role in Defining Cell Geometry; PLoS Biol 5(6): e149 doi:10.1371/journal.pbio.0050149 (Creative Commons Attribution License)
- ↑ Beisson, J. and Wright M. (2003).Basal body/centriole assembly and continuity Archivado el 17 de febrero de 2012 en Wayback Machine.. Current Opinion in Cell Biology 15, 96-104.
- ↑ Kardong, K. V (1999). Vertebrados. Anatomía comparada, función, evolución. McGraw-Hill Interamericana de España, S.A.U. ISBN 84-486-0261-7.
- ↑ Vogelstein, Bert; Kenneth W. Kinzler (2002). The Genetic Basis of Human Cancer. McGraw-Hill Professional. ISBN 0-07-137050-1.
- ↑ Miller S. L. (1953). «Production of Amino Acids Under Possible Primitive Earth Conditions». Science 117: 528. doi:10.1126/science.117.3046.528. Archivado desde el original el 28 de febrero de 2008.
- ↑ Lynn Sagan (1967). «On the origin of mitosing cells». J Theor Bio. 14 (3): 255-274. PMID 11541392. doi:10.1016/0022-5193(67)90079-3.
- ↑ Mereschowsky, C. (1910). Biol. Zentralbl 30 (3): 278 -367.
- ↑ Hyman Hartman y Alexei Fedorov (2001). «The origin of the eukaryotic cell: A genomic investigation». PNAS. Archivado desde el original el 15 de mayo de 2008. Consultado el 12 de abril de 2008.
- ↑ Rivera MC, Jain R, Moore JE, Lake JA (1997). «Genomic evidence for two functionally distinct gene classes». PNAS 95 (11). PMID 9600949.
- ↑ Rivera, MC y Lake, JA (2004). «The ring of life provides evidence for a genome fusion origin of eukaryotes». Nature. doi 10.1038/nature02848.
- ↑ Martin W y Müller M: (1998). «The hydrogen hypothesis for the first eukaryote». Nature 392 (6671). PMID 9510246.
- ↑ Poole AM, Penny D (2007). «Evaluating hypotheses for the origin of eukaryotes». Nature Reviews Genetics 8 (5). PMID 17429433.
- ↑ Origins: A Skeptic’s Guide, págs. 32, 49, 128.
- ↑ a b c Latorre, A.; Durban, A; Moya, A.; Pereto, J. (2011). «The role of symbiosis in eukaryotic evolution». En Gargaud, Muriel; López-Garcìa, Purificacion; Martin, H., eds. Origins and Evolution of Life: An astrobiological perspective. Cambridge: Cambridge University Press. pp. 326-339. ISBN 978-0-521-76131-4. Archivado desde el original
|urlarchivo=requiere|url=(ayuda) el 24 de marzo de 2019. Consultado el 27 de agosto de 2017. - ↑ Sampedro, Javier (2002). Deconstruyendo a Darwin. Barcelona: Crítica S.L. p. 47. ISBN 978-84-08-04135-1.
- ↑ Margulis, Lynn; Sagan, Dorion (2009). «5. Fusiones permanentes». ¿Qué es la vida? (3ª edición). Barcelona: Tusquets Editores S.A. pp. 90-117. ISBN 978-84-7223-799-5.
- ↑ McGrath, Casey (31 de mayo de 2022). «Highlight: Unraveling the Origins of LUCA and LECA on the Tree of Life». Genome Biology and Evolution 14 (6). PMC 9168435. doi:10.1093/gbe/evac072.
- ↑ Weiss, Madeline C.; Sousa, F. L.; Mrnjavac, N. et al. (2016). «The physiology and habitat of the last universal common ancestor». Nature Microbiology 1 (9): 16116. PMID 27562259. S2CID 2997255. doi:10.1038/nmicrobiol.2016.116.
- ↑ Leander, B. S. (Mayo 2020). «Predatory protists». Current Biology 30 (10): R510-R516. PMID 32428491. S2CID 218710816. doi:10.1016/j.cub.2020.03.052.
- ↑ Strassert, Jürgen F. H.; Irisarri, Iker; Williams, Tom A.; Burki, Fabien (25 de marzo de 2021). «A molecular timescale for eukaryote evolution with implications for the origin of red algal-derived plastids». Nature Communications 12 (1): 1879. Bibcode:2021NatCo..12.1879S. PMC 7994803. PMID 33767194. doi:10.1038/s41467-021-22044-z.
- ↑ Margulis, Lynn; Sagan, Dorion (2003). Captando genomas (1ª edición). Barcelona: Kairós. p. 103. ISBN 84-7245-551-3.
- ↑ Gabaldón, T. (October 2021). «Origin and Early Evolution of the Eukaryotic Cell». Annual Review of Microbiology 75 (1): 631-647. PMID 34343017. S2CID 236916203. doi:10.1146/annurev-micro-090817-062213.
- ↑ Woese, C.R.; Kandler, Otto; Wheelis, Mark L. (Junio 1990). «Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya». Proceedings of the National Academy of Sciences of the United States of America 87 (12): 4576-4579. Bibcode:1990PNAS...87.4576W. PMC 54159. PMID 2112744. doi:10.1073/pnas.87.12.4576.
- ↑ Alberts et al., Raff, p. 25-7, 805-7.
- ↑ Margulis, Lynn; Sagan, Dorion (2009). «4. Fusiones permanentes». ¿Qué es la vida? (3ª edición). Barcelona: Tusquets Editores S.A. pp. 107-110. ISBN 978-84-7223-799-5.
- ↑ Margulis, Lynn; Sagan, Dorion (2003). Captando genomas (1ª edición). Barcelona: Kairós. pp. 30, 132, 247. ISBN 84-7245-551-3.
Bibliografía
- Alberts et al (2004). Biología molecular de la célula. Barcelona: Omega. ISBN 54-282-1351-8.
- Alberts, Bruce; Johnson, Alexander; Lewis, Julian; Morgan, David; Raff, Martin; Roberts, K.; Walter, P. (2016). Biología molecular de la célula (6ª edición). Barcelona: Omega S.A. ISBN 978-84-282-1638-8.
- Lane, Nick (2005). Power, Sex, Suicide. Mitochondria and the Meaning of Life. Oxford University Press. ISBN 0-19-280481-2.
- Lodish et al. (2005). Biología celular y molecular. Buenos Aires: Médica Panamericana. ISBN 950-06-1974-3.
- Paniagua, R.; Nistal, M.; Sesma, P.; Álvarez-Uría, M.; Fraile, B.; Anadón, R. y José Sáez, F. (2002). Citología e histología vegetal y animal. McGraw-Hill Interamericana de España, S.A.U. ISBN 84-486-0436-9.
Enlaces externos
Wikiquote alberga frases célebres de o sobre Célula.
Wikimedia Commons alberga una categoría multimedia sobre Célula.
Wikcionario tiene definiciones y otra información sobre célula.
Wikilibros alberga un libro o manual sobre Biología celular.
- El Diccionario de la Real Academia Española tiene una definición para célula.
- Estructura celular.
- Libro en línea sobre biología celular (en inglés)