Cellevæg

En cellevæg er en struktur, der ligger uden på cellemembranen i celler hos bakterier, svampe, alger og planter. Hos planter består cellevæggen hovedsagelig af kulhydratet cellulose. Cellevæggen er med til at give organismerne struktur og form. Den beskytter cellens indre. Den er hård ved planteceller. Dyreceller har ingen cellevæg, de har kun en cellemembran.
Bakterier kan inddeles i gram-negative og gram-positive, hvor der ved de gram-positive er et tykt lag peptidoglykan.
Historie
[redigér | rediger kildetekst]En plantecellevæg blev først observeret og navngivet (simpelthen som en "væg") af Robert Hooke i 1665.[1] Imidlertid blev "det døde ekstruderingsprodukt fra den levende protoplast" glemt i næsten tre århundreder, idet det primært var genstand for videnskabelig interesse som en ressource til industriel forarbejdning eller i relation til dyrs eller menneskers sundhed.
I 1804 beviste Karl Rudolphi og Heinrich Friedrich Link, at celler havde uafhængige cellevægge.[2] Før havde man troet, at celler delte vægge, og at væske passerede mellem dem på denne måde.
Cellevæggens dannelsesmåde var kontroversiel i det 19. århundrede. Hugo von Mohl (1853, 1858) talte for ideen om, at cellevæggen vokser ved apposition. Carl Wilhelm von Nägeli (1858, 1862, 1863) mente, at væggens vækst i tykkelse og i areal skyldtes en proces kaldt intussusception. Hver teori blev forbedret i de følgende årtier: appositions- (eller laminerings-)teorien af Eduard Strasburger (1882, 1889) og teorien om intussusception af Julius Wiesner (1886).[3]
I 1930 opfandt Ernst Münch udtrykket apoplast for at adskille den "levende" symplast fra den "døde" planteregion, hvoraf sidstnævnte omfattede cellevæggen.[4]
I 1980'erne foreslog nogle forfattere at erstatte udtrykket "cellevæg", især brugt om planter, med det mere præcise udtryk "ekstracellulær matrix", som bruges om dyreceller, men andre foretrak det ældre udtryk.[5]

Egenskaber
[redigér | rediger kildetekst]Cellevægge kan have lignende egenskaber som de organisme, som de findes i. De kan give cellerne stivhed og styrke og dermed give beskyttelse mod mekanisk stres. Den kemiske komposition og de mekaniske egenskaber for cellevæggen er forbundet med plantens cellevækst og morfogenese.[6] I flercellede organisme giver cellevæggene organismen mulighed for at bygge og holde en bestemt form. Cellevægge kan også begrænse større molekyler, der kan være giftige for cellen, i at komme ind i organismen. De kan yderligere tillade skabelsen af et stabilt osmotisk miljø ved at forhindre cytolyse, og hjælpe med at holde på vand. Cellevæggenes komposition, egenskaber og form kan ændre sig under cellens cyklus, og de afhænger af vækstbetingelserne.[6]
Planters cellevæg
[redigér | rediger kildetekst]
Plantecellevæggen er en netværksstruktur, der primært består af polysakkarider såsom cellulose, hemicellulose og pektin, men som ofte også omfatter glykoproteiner og lignin. Denne struktur omslutter planteceller og giver dem forskellige former, størrelser og distinkte fysisk-kemiske egenskaber, der gør det muligt for dem at udfylde deres roller i plantens forskellige organer og i dens forskellige udviklingsstadier.[7] Plantecellevæggen spiller mange grundlæggende roller, herunder bestemmelse af plantemorfogenese og -arkitektur, yde mekanisk støtte til plantekroppen, lede vand og næringsstoffer, udgøre et forsvare mod biotiske og abiotiske udfordringer med mere.[8][9][10]. Derudover er plantecellevæggen den mest udbredte fornybare ressource på Jorden, som har stor praktisk værdi for mennesker, herunder som brug til dyrefoder og varmeproduktion, som byggemateriale og som kilde til naturlige fibre til tekstil- og papirproduktion.[11]
Plantecellevæggens opbygning og biosyntese
[redigér | rediger kildetekst]Cellevæggens opbygning bestemmer cellevæggens struktur og funktion. Efterhånden som planter tilpassede sig livet på jorden, udviklede og diversificerede cellevæggens sammensætning sig, hvilket i sidste ende resulterede i at forskellige plantearter har cellevægge med varierende kemi, struktur og funktion.[10][12] Identifikation af de proteiner, der kræves til biosyntesen af vigtige cellevægskomponenter, herunder cellulose, hemicellulose, pektin og lignin, er et væsentligt trin i afdækningen af plantecellevæggens struktur og -funktion.[7]
Cellulose
[redigér | rediger kildetekst]
Celluloserige cellevægge har stor betydning for terrestriske karplanter. Cellulose består af lineære fibriller samlet af glucan og er hovedpolymeren i de fleste plantecellevægge. Glucankæderne er uforgrenede og β-(1,4)-bundne. Selvom den kemiske natur af cellulose er meget enkel, samles kæderne på en kompleks måde til store ordnede cellulosefibre. Krystallinske cellulosefibre dannes ud fra glucankæder, som derefter bundtes sammen til nanofibre og aggregeres til en ledningslignende strukturer med forskellige diametre, som afsløret ved højopløsnings elektronmikroskopi og atomar kraftmikroskopi.[13][14] Disse multiskala cellulosefibre forbinder sig et fibrillært netværk, hvilket resulterer i en sammenhængende nanostruktur.[7]
Den strukturelle heterogenitet af cellulose afspejler kompleksiteten af polymerens biosyntese, som kræver aktiviteten af flere forskellige proteiner. Et af disse proteiner er cellulosesyntase, som også kaldes CESA:[15] CES (for cellulosesyntase) efterfulgt af A (for at angive at det er en homolog til bakterielle katalytiske subunits). CESA katalyserer en forlængelse af glucankæden der foregår direkte på cellemembranen. Genetiske undersøgelser afslørede, at cellulosesyntasekomplekset (CSC) i almindelig gåsemad (Arabidopsis thaliana) indeholder tre CESA-proteiner (CESA1, CESA3 og enten CESA2, CESA5, CESA6 eller CESA9). Protein-protein-interaktioner fører til at komplekset dannes, og det skal samles for at kunne producere cellulose i den primære cellevæg.[16][17] CSC bestående af CESA4, CESA7 og CESA8 er uundværlig for cellulosesyntese i den sekundære cellevæg.[18] Dette syntetiske maskineri er meget konserveret i plantearter, selv i den enkimbladede plante ris. Defekter i disse CESA'er resulterer i misvækst.[16][19][10][20] CESA-proteiner i forskellige plantearter deler høj sekvenslighed. CESA'er er afgørende proteiner, der direkte katalyserer celluloseforlængelse og er i centrum for forståelsen af hvordan cellen producerer cellulose.[7]
Opdagelsen af den tredimensionelle struktur af plante-CSC'er repræsenterer endnu et gennembrud. Elektronmikroskopi udført i 1980 afslørede, at plante-CSC'er er samlede i en større sekskantet struktur, der minder om en ”roset”,[21] hvilket gør CSC'er til et af planters mest komplekse membranproteinkomplekser.
Hemicellulose
[redigér | rediger kildetekst]Hemicellulose er et polysakkarid, der består af β-glycan-rygrad med forskellige substitutioner. De fleste cellevægspolymerer opnået ved alkalisk ekstraktion er hemicelluloseholdige polysakkarider, hvorimod pektinpolymerer let ekstraheres ved hjælp af varm syre eller chelatorer.[22] Hemicellulosepolysaccharider, herunder xylaner, xyloglucaner, mannaner, glucomannaner og β-(1,3;1,4)-glucan, har alle β-(1,4)-glycosyl-bundne rygrader.[22] De fleste hemicellulose-rygrader syntetiseres af cellulosesyntaselignende proteiner (CSL'er) fra GT2-familien af proteiner,[23][22] selvom glycosylresterne varierer. Undtagelsen er xylaner, hvis rygrad produceres af Type II-membranproteiner fra GT47- og GT43-familierne.[24][25][26] Hemicellulosernes rygrad er ofte erstattet af forskellige glycosylrester med visse mønstre, som bestemmer variationerne i deres fysisk-kemiske egenskaber og strukturer.[7]
Xylaner er de mest udbredte hemicellulosepolymerer i karplanter. Den samme β-(1,4)-koblede xylosyl-rygrad er til stede i forskellige plantearter, hvilket peger på et konserveret biosyntetisk maskineri i planter. Fordi den pentoseholdige rygrad er forskellig fra den hexoseholdige rygrad, syntetiseres xylan på sin egen måde.[27] Xylan er et stærkt acetyleret polysakkarid, som inkorporerer størstedelen af acetylestere i cellevæggen.[28][29]
Mannaner er lineære glycankæder af β-(1,4)-bundne mannoserester, mens glukomannan refererer til mannan, der indeholder indsat β-(1,4)-bundet glukose i sin rygrad. Galaktomannan og galaktoglukomannan har delvist galaktosylerede mannosylrester på O-6-positionen. Mannan-niveauer er normalt lave i blomsterplanter, men galaktoglucomannan er hovedbestanddelen i den sekundære cellevæg i gymnospermer. Mannaner er også fungerer også som opbevaring for polysakkarider i nogle plantearter.[7]
Pektiner
[redigér | rediger kildetekst]
Pektiner er strukturelle polysakkarider beriget med galakturonsyrer. Pektiner er således en gruppe af cellevægspolysakkarider, der har α-(1,4)-bundne galakturonsyrer (GalA) i deres rygrad og er klassificeret i tre typer eller domæner: uforgrenet homogalakturonan (HG), rhamnose-alterneret rhamnogalakturonan I (RG- I), og det komplekse forgrenede rhamnogalakturonan II (RG-II).[30][31] Homogalacturonan rummer en ikke-substitueret lineær kæde af α-(1,4)-bundet GalA. RG-II er den mest komplekse pektinpolymer, da den har mere end 20 bindinger på 12 forskellige sukkerarter for at danne den HG-lignende rygrad og fire oligosakkarid-sidekæder (A-D). Mens sidekæderne C og D er disakkarider, er sidekæde A et invariant octasakkarid med varierede methyleringsmodifikationer, der har udskiftelige L-fucose- eller L-galaktose-substitutioner. Sidekæde B besidder seks til ni sukkerarter med forskellige terminale rester og acetyleringsmodifikationer.[32]
Lignin
[redigér | rediger kildetekst]
Til forskel fra andre cellevægsbestanddele er ligniner ikke polysakkarider, men polyfenoler. Ligniner dannes således af polymeriserede aromatiske stoffer fra fenylpropanoid-vejen. Disse helt uordnede polymerer kan blive meget store og er sammensat af fenylalanin-afledte (og tyrosin-afledte i græs) aromatiske monomere forbindelser, der udgør en karakteristisk cellevægskomponent i karplanter.[33][34] Selvom lignin er amorft, forstærker det cellevægsstrukturen og fungerer ofte som en fysisk barriere mod patogenangreb. Lignin er normalt sammensat af guaiacyl, syringyl og nogle gange p-hydroxyphenyl monomerer afledt af komplekse fenylpropanoid metaboliske veje.[35] Til dato er mindst 35 naturlige ligninmonomerer blevet opdaget, hvilket fremhæver den metaboliske kompleksitet af lignin.[7]
Svampes cellevæg
[redigér | rediger kildetekst]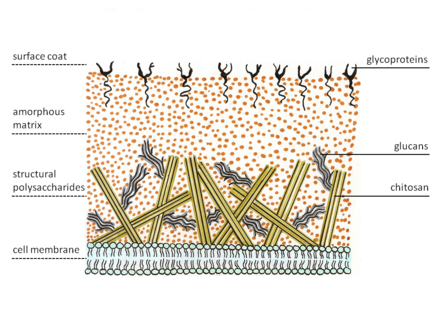
Svampes cellevæg er en essentiel struktur med stor plasticitet.[36] Cellevæggen spiller en vigtig rolle i forskellige biologiske funktioner såsom kontrol af cellulær permeabilitet og beskyttelse af cellen mod osmotisk og mekanisk stress.[37][38] Ud over disse vigtige funktioner medierer cellevæggen interaktioner med det ydre miljø gennem adhæsiner og et stort antal receptorer, der efter deres aktivering vil udløse en kompleks kaskade af signaler inde i cellen. Cellevæggen er unikt sammensat af polysakkarider og proteiner samt lipider og pigmenter.[37] Desuden er nogle vægkomponenter meget immunogene og stimulerer cellulære og humorale responser under infektion.[39] β-glucaner og mannaner, såvel som antistoffer rettet mod dem, er meget nyttige diagnostiske værktøjer, da de kan påvises hos patienter med invasiv svampeinfektion.[40] Eftersom cellevæggen er en essentiel struktur, kan defekter i den resultere i alvorlige virkninger på cellevækst og morfologi, og føre til celledød. Derfor betragtes den som et godt mål for svampemidler.[41][42][36]
Svampecellevæggens opbygning
[redigér | rediger kildetekst]Cellevæggen er en specifik og kompleks cellulær organel sammensat af glucaner, kitin, chitosan og glykosylerede proteiner. Proteiner er generelt forbundet med polysaccharider, hvilket resulterer i glycoproteiner. Sammen bidrager disse komponenter til cellevæggens stivhed. Syntese og vedligeholdelse af cellevæg involverer et stort antal biosyntese- og signalveje.
Cellevæggen er opbygget i forskellige lag, hvor det inderste lag er en mere konserveret struktur, hvorpå de resterende lag er aflejret og kan variere mellem forskellige svampearter.[36]
Glucaner
[redigér | rediger kildetekst]Glucan er det vigtigste strukturelle polysakkarid i svampecellevæggen og repræsenterer 50-60% af væggens tørvægt.[36] I glucan er D-glukose-enheder bundet sammen af glykosidbindinger. α- og β- og tal angiver hvilken type af O-glykosidbinding, der er tale om. De fleste polymerer af glucan er bundet sammen af β-1,3 bindinger (65-90%), selvom der også er glucaner med β-1,6 (i Candida men ikke i Aspergillus), β-1,4, α-1,3 og α-1,4 bindinger. β-1,3-D-glucanen er den vigtigste strukturelle komponent af svampens cellevæg, hvortil andre komponenter er bundet kovalent. β-1,3-D-glucan syntetiseres af et kompleks af enzymer placeret i cellemembranen kaldet glucansyntaser. Generne, der koder for β-1,3-D-glucaner, FKS1 og FKS2, blev oprindeligt identificeret i gærsvampen Saccharomyces cerevisiae.[43][44] Analoger af disse gener er kendt i flere arter af Candida, Aspergillus, Cryptococcus og Pneumocystis og flere andre svampe. Tab af funktion af et af disse gener påvirker cellevækst negativt,[43][45] medens eliminering af begge gener resulterer i celledød.[45][46] α-1,3-glucanen er også en grundlæggende komponent i svampens cellevæg og syntetiseres af α-glucansyntase (AGS1).
Kitin
[redigér | rediger kildetekst]Kitin er ligesom glucan et polysakkarid, der er opbygget i lange kæder af aminosukkerelementer (N-acetylglukosamin). Kitin findes ikke i planter, men indgår også i leddyrs ydre skelet. Som ved cellulose, der er det dominerende polysakkarid i planters cellevæg, bindes sukkerenhederne i kitin sammen med β-1,4-bindinger. Kitinindholdet i svampevæggen varierer afhængigt af svampens morfologiske fase. Det repræsenterer 1-2% af tørvægten af gærcellevæggen, mens det i filamentøse svampe kan nå op til 10-20%. Kitin syntetiseres ud fra n-acetylglucosamin af enzymet kitinsyntase, som afsætter kitinpolymerer i det ekstracellulære rum udenfor cellemembranen. Indholdet af kitin i cellevæggen kan vairere og er i C. albicans’ hyfe-væg fx tre gange højere end det er i gær.[47]
Glykoproteiner
[redigér | rediger kildetekst]Proteiner udgør 30-50% af tørvægten af svampevæggen i gær og 20-30% af tørvægten af væggen af de filamentøse svampe. De fleste proteiner er forbundet med kulhydrater ved O- eller N-bindinger, hvilket resulterer i glycoproteiner. Cellevægsproteiner har forskellige funktioner, herunder deltagelse i opretholdelsen af den cellulære form, adhæsionsprocesser, cellulær beskyttelse mod forskellige stoffer, absorption af molekyler, signaltransmission og syntese og reorganisering af vægkomponenter.[46]
Melanin
[redigér | rediger kildetekst]Melanin er et pigment med høj molekylvægt, der er negativt ladet, hydrofobt og uopløseligt i vandige opløsninger og beskytter svampe mod stressfaktorer i omgivelserne.[48] Svampene producerer melanin ad to veje, fra 1,8-dihydroxynaphthalen (DHN) mellemprodukt og fra L-3,4-dihydroxyphenylalanin (L-dopa).[49] Melaninproduktion bidrager til svampevirulens,[50] forbedrer modstanden mod miljøskader såsom ekstrem temperatur,[51][52] UV-lys og toksiner,[49] og er vigtig for invasion og formidling. For eksempel er C. neoformans melanin blevet forbundet med spredning af gærceller fra lungerne til andre organer,[53] er kendt for at påvirke værtens immunrespons. ,[49]
Prokaryotes cellevægge
[redigér | rediger kildetekst]Egentlige bakteriers cellevægge
[redigér | rediger kildetekst]

Egentlige bakteriers cellevæg befinder sig på ydersiden af cellemembranen. Bakteriecellevægge er lavet af peptidoglycan (også kaldet murein), som er lavet af polysakkarid-kæder bundet sammen på tværs af usædvanlige peptider indeholdende D-aminosyre-rester.[54] Bakteriecellevægge er forskellige fra cellevæggene hos planter og svampe, som henholdsvis primært er opbygget af cellulose og kitin.[55] Bakteriers cellevæg adskiller sig også fra Archaeas cellevæg, som ikke indeholder peptidoglycan. Cellevæggen er afgørende for overlevelsen af mange bakterier, selvom bakterier, der mangler en cellevæg, kan produceres og overleve i laboratoriet.[56] Antibiotikummet penicillin er i stand til at dræbe bakterier ved at forhindre tværbinding af peptidoglycan og dette får cellevæggen til at svækkes, svulme op og sprænges.[55] Enzymet lysozym kan også beskadige bakteriers cellevægge.
Der er overordnet to forskellige typer cellevæg i bakterier, kaldet gram-positive og gram-negative bakterier. Navnene stammer fra cellernes reaktion på Gramfarvning, en test, der længe har været anvendt til klassificering af bakteriearter.[57]
Gram-positive bakterier har en tyk cellevæg, der indeholder mange lag af peptidoglycan og teikoidsyrer. I modsætning hertil har gramnegative bakterier en relativt tynd cellevæg bestående af et par lag peptidoglycan omgivet af en anden lipidmembran indeholdende lipopolysakkariderer og lipoproteiner. De fleste bakterier har den gram-negative cellevæg, og kun Bacillota og Actinomycetota (tidligere kendt som henholdsvis lav G+C og høj G+C gram-positive bakterier) har det alternative gram-positive arrangement.[58] Disse forskelle i struktur kan forårsage forskelle i antibiotikafølsomhed, f.eks. kan vancomycin kun dræbe gram-positive bakterier og er ineffektiv over for gram-negative patogener, såsom Haemophilus influenzae eller Pseudomonas aeruginosa.[59]
Arkæers cellevægge
[redigér | rediger kildetekst]Cellevæggene i Arkæer adskiller sig på et afgørende punkt fra dem i egentlige bakterier. Mens peptidoglycan er indgår i cellevæggene hos egentlige bakterier, mangler alle arkæale cellevægge peptidoglycan,[60] selvom nogle methanogener har en cellevæg lavet af en lignende polymer kaldet pseudopeptidoglycan.[61] Man finder fire typer cellevægge blandt Arkæer.
En type archaeal cellevæg er den, der består af pseudopeptidoglycan (også kaldet pseudomurein). Denne type cellevæg findes i nogle methanogener, såsom Methanobacterium og Metanothermus.[62] Mens den overordnede struktur af arkæal pseudopeptidoglycan overfladisk ligner den for bakteriel peptidoglycan, er der en række væsentlige kemiske forskelle. Ligesom peptidoglycanen, der findes i bakterielle cellevægge, består pseudopeptidoglycan af polymer-kæder af glycan tværbundet af korte peptid-forbindelser. Men i modsætning til peptidoglycan er sukkeret N-acetylmuraminsyre erstattet af N-acetyltalosaminuronsyre,[60] og de to sukre er bundet med en β,1-3 glykosidbinding i stedet for β,1-4. Derudover er de tværbindende peptider L-aminosyrer snarere end D-aminosyrer, som det er tilfældet hos bakterier.[62]
En anden type archaeal cellevæg findes i Methanosarcina og Halococcus. Denne type cellevæg består udelukkende af et tykt lag af polysakkarider, som kan være sulfaterede som det ses hos Halococcus.[62] Strukturen i denne type væg er kompleks og ikke fuldt ud undersøgt.
En tredje type væg blandt Arkæa består af glykoprotein, og forekommer i den hypertermofile arkæ Halobacterium og nogle methanogener. I Halobacterium har proteinerne i væggen et højt indhold af aminosyrerne aspartat og glutamat, hvilket giver væggen en samlet negativ ladning. Resultatet er en ustabil struktur, der stabiliseres af tilstedeværelsen af store mængder positive natriumioner, der neutraliserer de negative ladninger.[62] På grund af sin evne til at binde Na+ trives Halobacterium kun under forhold med høj saltholdighed.
I andre Archæa, såsom Methanomicrobium og Desulfurococcus, er væggen kun sammensat af overfladelag af proteiner,[61] kendt som S-laget. S-laget er almindelige hos bakterier, hvor de tjener som enten den eneste cellevægskomponent eller et ydre lag i forbindelse med polysakkarider. De fleste Arkæa er Gram-negative, selvom der kendes mindst ét Gram-positivt medlem.[61]
Referencer
[redigér | rediger kildetekst]- ^ Micrographia: or, Some physiological descriptions of minute bodies made by magnifying glasses. London. 1665.
- ^ Grundlehren der anatomie und physiologie der pflanzen. Danckwerts. 1807.
- ^ Introduction To Cytology. New York: McGraw Hill. 1921. s. 25.
- ^ Die Stoffbewegungen in der Pflanze. Jena: Verlag von Gustav Fischer. 1930.
- ^ Esau's Plant Anatomy: Meristems, Cells, and Tissues of the Plant Body: Their Structure, Function, and Development (3rd udgave). Hoboken, New Jersey: John Wiley & Sons, Inc. 2006. s. 65-66. ISBN 978-0-470-04737-8.
- ^ a b Bidhendi AJ, Geitmann A (januar 2016). "Relating the mechanics of the primary plant cell wall to morphogenesis". Journal of Experimental Botany. 67 (2): 449-61. doi:10.1093/jxb/erv535. PMID 26689854.
- ^ a b c d e f g Zhang B, Gao Y, Zhang L, Zhou Y (2021) The plant cell wall: Biosynthesis, construction, and functions. Journal of Integrative Plant Biology. 63(1):251-272. doi: 10.1111/jipb.13055. PMID: 33325153.
- ^ Bacic, A., Harris, P. and Stone, B. (1988). Structure and function of plant cell walls. In The Biochemistry of Plants, J. Priess, ed (New York/London/San Francisco: Academic Press), pp. 297−371.
- ^ Carpita NC, Gibeaut DM (1993) Structural models of primary cell walls in flowering plants: consistency of molecular structure with the physical properties of the walls during growth. Plant Journal. 3(1):1-30. doi: 10.1111/j.1365-313x.1993.tb00007.x. PMID: 8401598.
- ^ a b c Somerville C, Bauer S, Brininstool G, Facette M, Hamann T, Milne J, Osborne E, Paredez A, Persson S, Raab T, Vorwerk S, Youngs H (2004) Toward a systems approach to understanding plant cell walls. Science. 306(5705):2206-11. doi: 10.1126/science.1102765. PMID: 15618507.
- ^ Pauly M, Keegstra K (2010) Plant cell wall polymers as precursors for biofuels. Current Opininion in Plant Biology. 13(3):305-12. doi: 10.1016/j.pbi.2009.12.009. PMID: 20097119.
- ^ Burton RA, Gidley MJ, Fincher GB. Heterogeneity in the chemistry, structure and function of plant cell walls. Nature Chemical Biology. 6(10):724-32. doi: 10.1038/nchembio.439. Epub 2010 Sep 17. PMID: 20852610.
- ^ Ding SY, Liu YS, Zeng Y, Himmel ME, Baker JO, Bayer EA (2012) How does plant cell wall nanoscale architecture correlate with enzymatic digestibility? Science. 338(6110):1055-60. doi: 10.1126/science.1227491. PMID: 23180856.
- ^ Zhang L, Gao C, Mentink-Vigier F, Tang L, Zhang D, Wang S, Cao S, Xu Z, Liu X, Wang T, Zhou Y, Zhang B (2019) Arabinosyl Deacetylase Modulates the Arabinoxylan Acetylation Profile and Secondary Wall Formation. Plant Cell. 31(5):1113-1126. doi: 10.1105/tpc.18.00894. PMID: 30886126; PMCID: PMC6533017.
- ^ Delmer DP (1999) CELLULOSE BIOSYNTHESIS: Exciting Times for A Difficult Field of Study. Annual Review of Plant Physiology and Plant Molecular Biology. 50:245-276. doi: 10.1146/annurev.arplant.50.1.245. PMID: 15012210.
- ^ a b Arioli T, Peng L, Betzner AS, Burn J, Wittke W, Herth W, Camilleri C, Höfte H, Plazinski J, Birch R, Cork A, Glover J, Redmond J, Williamson RE (1998) Molecular analysis of cellulose biosynthesis in Arabidopsis. Science. 79(5351):717-20. doi: 10.1126/science.279.5351.717. PMID: 9445479.
- ^ Scheible WR, Eshed R, Richmond T, Delmer D, Somerville C. Modifications of cellulose synthase confer resistance to isoxaben and thiazolidinone herbicides in Arabidopsis Ixr1 mutants. Proceedings of the National Academy of Sciences U S A. 98(18):10079-84. doi: 10.1073/pnas.191361598. PMID: 11517344; PMCID: PMC56918.
- ^ Taylor NG, Howells RM, Huttly AK, Vickers K, Turner SR (2003) Interactions among three distinct CesA proteins essential for cellulose synthesis. Proceedings of the National Academy of Sciences U S A. 100(3):1450-5. doi: 10.1073/pnas.0337628100. PMID: 12538856; PMCID: PMC298793.
- ^ Tanaka K, Murata K, Yamazaki M, Onosato K, Miyao A, Hirochika H. Three distinct rice cellulose synthase catalytic subunit genes required for cellulose synthesis in the secondary wall. Plant Physiology. 133(1):73-83. doi: 10.1104/pp.103.022442. PMID: 12970476; PMCID: PMC196581.
- ^ Zhang B, Deng L, Qian Q, Xiong G, Zeng D, Li R, Guo L, Li J, Zhou Y (2009) A missense mutation in the transmembrane domain of CESA4 affects protein abundance in the plasma membrane and results in abnormal cell wall biosynthesis in rice. Plant Molecular Biology. 71(4-5):509-24. doi: 10.1007/s11103-009-9536-4. Epub 2009 Aug 21. PMID: 19697141.
- ^ Mueller SC, Brown RM Jr (1980) Evidence for an intramembrane component associated with a cellulose microfibril-synthesizing complex in higher plants. Journal of Cell Biology. 84(2):315-26. doi: 10.1083/jcb.84.2.315. PMID: 7189755; PMCID: PMC2110546.
- ^ a b c Scheller HV, Ulvskov P. Hemicelluloses. Annual Review of Plant Biology. 61:263-89. doi: 10.1146/annurev-arplant-042809-112315. PMID: 20192742.
- ^ Richmond TA, Somerville CR (2000) The cellulose synthase superfamily. Plant Physiology. 124(2):495-8. doi: 10.1104/pp.124.2.495. PMID: 11027699; PMCID: PMC1539280.
- ^ Brown DM, Goubet F, Wong VW, Goodacre R, Stephens E, Dupree P, Turner SR (2007) Comparison of five xylan synthesis mutants reveals new insight into the mechanisms of xylan synthesis. Plant Journal. 52(6):1154-68. doi: 10.1111/j.1365-313X.2007.03307.x. PMID: 17944810.
- ^ Brown DM, Zhang Z, Stephens E, Dupree P, Turner SR. Characterization of IRX10 and IRX10-like reveals an essential role in glucuronoxylan biosynthesis in Arabidopsis. Plant Journal. 57(4):732-46. doi: 10.1111/j.1365-313X.2008.03729.x. PMID: 18980662.
- ^ Wu AM, Hörnblad E, Voxeur A, Gerber L, Rihouey C, Lerouge P, Marchant A (2010) Analysis of the Arabidopsis IRX9/IRX9-L and IRX14/IRX14-L pairs of glycosyltransferase genes reveals critical contributions to biosynthesis of the hemicellulose glucuronoxylan. Plant Physiology. 153(2):542-54. doi: 10.1104/pp.110.154971. PMID: 20424005; PMCID: PMC2879767.
- ^ Smith PJ, Wang HT, York WS, Peña MJ, Urbanowicz BR (2017) Designer biomass for next-generation biorefineries: leveraging recent insights into xylan structure and biosynthesis. Biotechnology for Biofuels. 10:286. doi: 10.1186/s13068-017-0973-z. PMID: 29213325; PMCID: PMC5708106.
- ^ Gille S, Pauly M (2012) O-acetylation of plant cell wall polysaccharides. Frontiers in Plant Science. 3:12. doi: 10.3389/fpls.2012.00012. PMID: 22639638; PMCID: PMC3355586.
- ^ Xiong G, Cheng K, Pauly M (2013) Xylan O-acetylation impacts xylem development and enzymatic recalcitrance as indicated by the Arabidopsis mutant tbl29. Molecular Plant. 6(4):1373-5. doi: 10.1093/mp/sst014. PMID: 23340742.
- ^ Mohnen D (2008) Pectin structure and biosynthesis. Current Opinion in Plant Biology. 11(3):266-77. doi: 10.1016/j.pbi.2008.03.006. PMID: 18486536.
- ^ Atmodjo MA, Hao Z, Mohnen D (2013) Evolving views of pectin biosynthesis. Annual Review of Plant Biology. 64:747-79. doi: 10.1146/annurev-arplant-042811-105534. PMID: 23451775.
- ^ Pabst M, Fischl RM, Brecker L, Morelle W, Fauland A, Köfeler H, Altmann F, Léonard R (2013) Rhamnogalacturonan II structure shows variation in the side chains monosaccharide composition and methylation status within and across different plant species. Plant Journal. 76(1):61-72. doi: 10.1111/tpj.12271. PMID: 23802881.
- ^ Barros J, Serrani-Yarce JC, Chen F, Baxter D, Venables BJ, Dixon RA. Role of bifunctional ammonia-lyase in grass cell wall biosynthesis. Nature Plants. 2(6):16050. doi: 10.1038/nplants.2016.50. PMID: 27255834.
- ^ Vanholme R, Cesarino I, Rataj K, Xiao Y, Sundin L, Goeminne G, Kim H, Cross J, Morreel K, Araujo P, Welsh L, Haustraete J, McClellan C, Vanholme B, Ralph J, Simpson GG, Halpin C, Boerjan W (2013) Caffeoyl shikimate esterase (CSE) is an enzyme in the lignin biosynthetic pathway in Arabidopsis. Science. 341(6150):1103-6. doi: 10.1126/science.1241602. PMID: 23950498.
- ^ Vanholme R, De Meester B, Ralph J, Boerjan W (2019) Lignin biosynthesis and its integration into metabolism. Current Opinion in Biotechnology. 56:230-239. doi: 10.1016/j.copbio.2019.02.018. PMID: 30913460.
- ^ a b c d Garcia-Rubio R, de Oliveira HC, Rivera J, Trevijano-Contador N (2020) The Fungal Cell Wall: Candida, Cryptococcus, and Aspergillus Species. Frontiers in Microbiology 10:2993. doi: 10.3389/fmicb.2019.02993. PMID: 31993032; PMCID: PMC6962315.
- ^ a b Gow NAR, Latge JP, Munro CA (2017) The Fungal Cell Wall: Structure, Biosynthesis, and Function. Microbiology Spectrum 5(3). doi: 10.1128/microbiolspec.FUNK-0035-2016. PMID: 28513415.
- ^ Agustinho DP, Miller LC, Li LX, Doering TL (2018) Peeling the onion: the outer layers of Cryptococcus neoformans. Memórias do Instituto Oswaldo Cruz. 113:e180040. doi: 10.1590/0074-02760180040. PMID: 29742198; PMCID: PMC5951675.
- ^ Erwig LP, Gow NA (2016) Interactions of fungal pathogens with phagocytes. Nature Reviews Microbiology. 14(3):163-76. doi: 10.1038/nrmicro.2015.21. PMID: 26853116.
- ^ Pazos C, Moragues MD, Quindós G, Pontón J, del Palacio A (2006) Diagnostic potential of (1,3)-beta-D-glucan and anti-Candida albicans germ tube antibodies for the diagnosis and therapeutic monitoring of invasive candidiasis in neutropenic adult patients. Rev Iberoam Micol. 23(4):209-15. doi: 10.1016/s1130-1406(06)70046-8. PMID: 17388644.
- ^ Heitman J (2005) Cell biology. A fungal Achilles' heel. Science 309(5744):2175-6. doi: 10.1126/science.1119321. PMID: 16195450
- ^ Cortés JCG, Curto MÁ, Carvalho VSD, Pérez P, Ribas JC (2019) The fungal cell wall as a target for the development of new antifungal therapies. Biotechnological Advances 37(6):107352. doi: 10.1016/j.biotechadv.2019.02.008. Epub 2019 Feb 21. PMID: 30797093.
- ^ a b Douglas CM, Foor F, Marrinan JA, Morin N, Nielsen JB, Dahl AM, Mazur P, Baginsky W, Li W, el-Sherbeini M (1994) The Saccharomyces cerevisiae FKS1 (ETG1) gene encodes an integral membrane protein which is a subunit of 1,3-beta-D-glucan synthase. Proceedings of the National Academy of Sciences U S A 91(26):12907-11. doi: 10.1073/pnas.91.26.12907. PMID: 7528927; PMCID: PMC45549.
- ^ Qadota H, Python CP, Inoue SB, Arisawa M, Anraku Y, Zheng Y, Watanabe T, Levin DE, Ohya Y (1996) Identification of yeast Rho1p GTPase as a regulatory subunit of 1,3-beta-glucan synthase. Science. 272(5259):279-81. doi: 10.1126/science.272.5259.279. PMID: 8602515.
- ^ a b Mazur P, Morin N, Baginsky W, el-Sherbeini M, Clemas JA, Nielsen JB, Foor F (1995) Differential expression and function of two homologous subunits of yeast 1,3-beta-D-glucan synthase. Molecular and Cellular Biology. 15(10):5671-81. doi: 10.1128/MCB.15.10.5671. PMID: 7565718; PMCID: PMC230817.
- ^ a b Bowman SM, Free SJ (2006) The structure and synthesis of the fungal cell wall. Bioessays. 28(8):799-808. doi: 10.1002/bies.20441. PMID: 16927300.
- ^ Chattaway FW, Holmes MR, Barlow AJ (1968) Cell wall composition of the mycelial and blastospore forms of Candida albicans. J Gen Microbiol. 51(3):367-76. doi: 10.1099/00221287-51-3-367. PMID: 5657261.
- ^ Nosanchuk JD, Casadevall A. Impact of melanin on microbial virulence and clinical resistance to antimicrobial compounds. Antimicrobial Agents and Chemotherapy. 50(11):3519-28. doi: 10.1128/AAC.00545-06. PMID: 17065617; PMCID: PMC1635213.
- ^ a b c Eisenman HC, Casadevall A (2012) Synthesis and assembly of fungal melanin. Appl Microbiol Biotechnol. 93(3):931-40. doi: 10.1007/s00253-011-3777-2. PMID: 22173481; PMCID: PMC4318813.
- ^ Nosanchuk JD, Stark RE, Casadevall A (2015) Fungal Melanin: What do We Know About Structure? Frontiers in Microbiology 6:1463. doi: 10.3389/fmicb.2015.01463. PMID: 26733993; PMCID: PMC4687393.
- ^ Rosa LH, Almeida Vieira Mde L, Santiago IF, Rosa CA (2010) Endophytic fungi community associated with the dicotyledonous plant Colobanthus quitensis (Kunth) Bartl. (Caryophyllaceae) in Antarctica. FEMS Microbiol Ecol. 73(1):178-89. doi: 10.1111/j.1574-6941.2010.00872.x. PMID: 20455944
- ^ Zalar P, Novak M, de Hoog GS, Gunde-Cimerman N (2011) Dishwashers - a man-made ecological niche accommodating human opportunistic fungal pathogens. Fungal Biology. 115(10):997-1007. doi: 10.1016/j.funbio.2011.04.007. PMID: 21944212.
- ^ Noverr MC, Williamson PR, Fajardo RS, Huffnagle GB (2004) CNLAC1 is required for extrapulmonary dissemination of Cryptococcus neoformans but not pulmonary persistence. Infect Immun. 72(3):1693-9. doi: 10.1128/IAI.72.3.1693-1699.2004. PMID: 14977977; PMCID: PMC356011.
- ^ van Heijenoort J (2001). "Formation of the glycan chains in the synthesis of bacterial peptidoglycan". Glycobiology. 11 (3): 25R-36R. doi:10.1093/glycob/11.3.25R. PMID 11320055.
- ^ a b Koch AL (oktober 2003). "Bacterial wall as target for attack: past, present, and future research". Clinical Microbiology Reviews. 16 (4): 673-87. doi:10.1128/CMR.16.4.673-687.2003. PMC 207114. PMID 14557293.
- ^ Joseleau-Petit D, Liébart JC, Ayala JA, D'Ari R (september 2007). "Unstable Escherichia coli L forms revisited: growth requires peptidoglycan synthesis". Journal of Bacteriology. 189 (18): 6512-20. doi:10.1128/JB.00273-07. PMC 2045188. PMID 17586646.
- ^ Gram, HC (1884). "Über die isolierte Färbung der Schizomyceten in Schnitt- und Trockenpräparaten". Fortschr. Med. 2: 185-189.
- ^ Hugenholtz P (2002). "Exploring prokaryotic diversity in the genomic era". Genome Biology. 3 (2): REVIEWS0003. doi:10.1186/gb-2002-3-2-reviews0003. PMC 139013. PMID 11864374.
- ^ Walsh F, Amyes S (2004). "Microbiology and drug resistance mechanisms of fully resistant pathogens" (PDF). Curr Opin Microbiol. 7 (5): 439-44. doi:10.1016/j.mib.2004.08.007. PMID 15451497.
- ^ a b White D (1995). The Physiology and Biochemistry of Prokaryotes. Oxford: Oxford University Press. s. 6, 12-21. ISBN 978-0-19-508439-9.
- ^ a b c Howland JL (2000). The Surprising Archaea: Discovering Another Domain of Life. Oxford: Oxford University Press. s. 69–71. ISBN 978-0-19-511183-5.
- ^ a b c d Brock TD, Madigan MT, Martinko JM, Parker J (1994). Biology of Microorganisms (7th udgave). Englewood Cliffs, NJ: Prentice Hall. s. 818–819, 824. ISBN 978-0-13-042169-2.
Eksterne henvisninger
[redigér | rediger kildetekst]
| Botanikkens underemner |  | |
|---|---|---|
| Planter | ||
| Planteorganer | ||
| Planteceller | ||
| Planters livscyklus | ||
| Planters taxonomi | ||
| Spire Denne artikel om biologi er en spire som bør udbygges. Du er velkommen til at hjælpe Wikipedia ved at udvide den. |