롱기스콰마
Longisquama| 롱기스콰마 | |
|---|---|
 | |
| 모식 시료의 주조 | |
| 과학적 분류 | |
| 왕국: | 애니멀리아 |
| 문: | 챠다타 |
| 클래스: | 파충류 |
| Clade: | 네오디압시다 |
| 속: | †롱기스콰마 샤로프, 1970년 |
| 종류: | ② L. 휘장 |
| 이항명 | |
| †롱기스콰마 휘장 샤로프, 1970년 | |
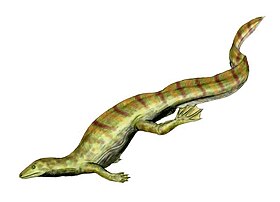
롱기스콰마는 멸종된 파충류의 한 속이다.키르기스스탄의 트라이아스기 마디겐 층에서 나온 불완전한 화석 흔적과 잘 보존되지 않은 골격으로 알려진 롱기스콰마 휘장은 단 한 종뿐입니다.화석 표본, 슬래브 및 카운터랩 유형(PIN 2548/4 및 PIN 2584/5)과 가능한 부속물의 참조 표본 5개(PIN 2584/7 ~ 9)를 통해 알려져 있다.모든 표본은 모스크바에 있는 러시아 과학 아카데미 고생물학 연구소에 소장되어 있다.
롱기스콰마는 "긴 비늘"을 의미하며, 구체적인 이름 휘장은 그것의 작은 크기를 나타냅니다.롱기스콰마 완모형은 피부에서 자라는 것으로 보이는 많은 긴 구조물들로 유명하다.현재 롱기스콰마는 애매모호하고 새의 기원과는 무관하다는 의견이 있다.
역사
해석

Haubold와 Vuffetaut는 그 구조들이 한 쌍의 [1]활공막처럼 신체의 측면 벽에 쌍으로 붙어 있는 길고 변형된 비늘이라고 믿었다.그들은 드라코종이나 쿠에네오사우루스 라투스와 같은 활공 도마뱀과 비슷한 패턴의 깃털을 가진 롱기스콰마를 재탄생시켜 활공하거나 최소한 낙하산을 탈 수 있게 한 것을 발표했다.비록 현재 그 재건이 부정확했다고 생각되지만, 그것의 버전은 여전히 현대의 [citation needed]고미술품에서 종종 묘사된다.
다른 연구자들은 척도를 다르게 배치합니다.Unwin과 Benton은 그것들을 등쪽 [2]중간선을 따라 이어지는, 짝을 이루지 않은 하나의 변형된 비늘줄로 해석했다.존스 외 연구진은 그것들을 해부학적으로 깃털과 매우 흡사한 두 쌍의 구조물로 해석했고, 그것들은 새의 척추 깃털 [3]관로의 그것들과 비슷한 위치에 있었다.깃털 발달 전문가인 리차드 프룸(그리고 레이즈와 수에스도)은 이 구조물을 해부학적으로 깃털과 매우 다르다고 보고, 길고 리본 같은 [4][5]비늘이라고 생각합니다.
다른 관찰자들(2006년 프레이저 등)은 이 구조물이 롱기스콰마의 일부가 아니라 파충류와 함께 보존된 식물 잎일 뿐이며 잘못 [6]해석된 것이라고 믿고 있다.Buchwitz & Voigt(2012)는 롱기스콰마의 구조물은 식물 유적이 아니라고 주장한다. 왜냐하면 완모식자 PIN 2584/4의 마지막을 제외한 모든 구조물은 규칙적으로 배열되어 있고 마디겐 [7]층의 식물 보존의 일반적인 방식인 탄소막으로 보존되어 있지 않기 때문이다.마디겐에서 롱기스콰마 구조와 유사한 유일한 식물은 메세테리오필룸 코츠네비이지만, 그 잎은 롱기스콰마에서 [7]기인한 구조의 뚜렷한 하키 스틱 모양을 가지고 있지 않다.
묘사
피복 구조

롱기스콰마는 등을 따라 독특한 피막 구조를 가진 것이 특징이다.완모식표본(specimen PIN 2584/4)은 연관된 골격의 뒤쪽에서 돌출된 이러한 부속물을 보존하는 유일한 화석입니다.7개의 부속물이 부채꼴 모양으로 뻗어 있지만 끝부분은 보존되어 있지 않습니다.PIN 2584/9는 5개의 부속품을 서로 간격을 두고 보관합니다.PIN 2584/6은 두 개의 긴 곡선의 부속품을 나란히 이어 유지합니다.PIN 2585/7 및 FG 596/V/1과 같은 다른 검체는 부속품을 하나만 보존합니다.이 구조물은 대부분 길이에 걸쳐 길고 좁으며, 하키 스틱처럼 보이기 위해 끝 부근에서 뒤로 기울어져 있다.근위부 직선 섹션은 세 개의 세로 로브(양쪽의 매끄러운 로브와 그 사이에 가로로 갈라진 로브)로 나뉩니다.가운데 주름이 있는 잎은 솟아오른 "루게"와 깊은 "간격"으로 이루어져 있는데, 샤로프는 이것을 묵주 구슬과 비교합니다.원위부 부분은 근위부 부분의 중앙 및 전엽의 연장선상에 있는 것으로 생각된다.전엽은 원위부에서 넓어지는 반면 근위부의 후엽은 원위부의 기부에서 끝날 때까지 좁아진다.또한 "외측 플랜지"는 근위부 섹션의 약 3분의 2까지 나타나 원위부 섹션의 선단까지 계속된다.원위부의 양로브는 홈이 있는 축에 의해 돌출되어 분리된다.일부 표본의 경우 원위부 엽의 융단이 서로 일렬로 정렬되는 반면, 다른 표본의 경우 그렇지 않습니다.축에 수직으로 돌출된 직선 러그가 있는 시료와 S자 모양으로 곡선을 이루는 러그가 있는 시료도 있다.Longisquama의 표본 중 하나인 PIN 2584/5는 원위부 [7]축에서 돌출된 작은 가시를 가지고 있습니다.
완모형 골격은 척추에 붙어있는 각각의 구조를 보여준다.이러한 고정점은 상승 노브로 볼 수 있습니다.각 부속물의 밑부분은 구조물의 평평한 모양과는 달리 약간 볼록하다.볼록한 모양은 포유류의 털이나 새의 깃털과 같은 다른 부속 구조물처럼 모낭에 고정되면서 각 구조물의 기초가 생활 속에서 관상이었다는 증거일 수 있다.또한 각 구조가 대응하는 척추에 근접해 있다는 것은 모낭을 포함한 연조직의 두꺼운 층이 [7]각 베이스를 둘러싸고 있다는 것을 시사한다.
분류
롱기스콰마의 골격적 특징도 '롱기스콰마'와 마찬가지로 진단하기 어렵다.그 결과, 롱기스콰마는 과학자들에 의해 많은 다른 용각류 집단들과 연관되어 왔다.
샤로프는 하악골 회향과 안와골 회향의 [8]두 가지 특징에 기초하여 그것이 "의도수키안" ("원시적인" 아치공룡이지만 상대적으로 파생된 파충류)이라고 결론지었다.샤로프의 원래 묘사에는 긴 견갑골도 포함되어 있다.존스 등은 롱기스콰마를 대룡으로 보고 샤로프의 두 캐릭터에 퍼큘라를 [3]추가했다.올셰프스키는 롱기스콰마가 대룡이고, 게다가 초기 [9]공룡이며, 만약 사실이라면 새의 [clarification needed]기원에 대한 거의 모든 논쟁을 없앨 수 있을 것이라고 믿고 있다.
언윈과 벤튼은 중요한 페네스트래를 진단하는 것이 불가능하다고 생각했다; 그 구멍들은 단순히 [2]화석에 대한 손상일 수 있다.그들은 롱기스콰마가 악로돈트 이빨과 쇄골을 가지고 있다는 샤로프의 의견에 동의했지만, 털갈이 대신, 그들은 쌍으로 된 쇄골을 보았다.이러한 특징들은 Lepidosauromorpha의 일원에 더 전형적인 것으로, Longisquama는 대룡이 아니기 때문에 조류와 밀접한 관계가 없다는 것을 의미한다.2004년 필 센터의 분지학적 연구에 따르면, 롱기스콰마는 훨씬 더 기초적인 디아피드이며 실루로사우라부스와 [10]더 가까운 Avicephala의 일원이다.
2012년 이 화석을 재조사한 결과, 대룡으로 분류하는 데 중요한 두개골의 존재는 확인되지 않았다.사실, 이전에 안와전향의 존재를 정당화하기 위해 사용되었던 화석 석판 중 하나의 두개골 부분에는 실제 뼈가 포함되어 있지 않다.이 연구는 롱기스콰마의 제안된 분류 중 어떤 것도 이용 가능한 증거를 사용하여 확인하거나 반박할 수 없다고 결론지었다.연구 저자들은 깃털, 새 깃털, 악어 비늘, 익룡 피노피버 [7]사이의 발달적인 "깊은 호몰로지" 가설의 결과로 롱기스콰마를 잠정적으로 아르코사우루스 형상에 포함시켰다.
새의 기원에 대한 논쟁


롱기스콰마의 파충류 분류와 '긴 비늘'의 정확한 기능에 관한 질문은 새의 기원과 공룡의 후손인지에 대한 광범위한 논쟁과 관련이 있다.
배경
고생물학자들의 일치된 의견은 새들이 수각류 공룡에서 진화했다는 것에 동의한다.이 가설의 시나리오는 초기 수각류 공룡이 흡열성이었고 단열성을 위해 단순한 필라멘트 모양의 깃털을 진화시켰다는 것이다.이 깃털들은 나중에 크기와 복잡성이 증가하여 공기역학적 용도에 적응하였다.이 가설에 대한 충분한 증거가 화석 기록에서 발견되었는데, 특히 쿨린다드롬우스, 시노사우롭테릭스, 카우디프테릭스, 미크로랍토르 그리고 많은 다른 공룡들에 대한 것이다.따라서 롱기스콰마는 이상한 비늘, 애매한 골격의 특징, 그리고 조류 진화에 실질적인 의미가 없는 반딧불이로 여겨진다.
소수의 과학자들은 새들이 롱기스콰마와 같은 작은 나무 위에 사는 아치오사우루스로부터 진화했다는 가설을 선호한다.그들은 이들을 길쭉한 비늘과 페니지 깃털을 발달시켜 활공에 적응한 외온성 동물로 본다.그러나 이 가설은 클래디스트 [11]분석에 의해 뒷받침되지 않는다.
레퍼런스
- ^ Haubold, H. & Buffetaut, E. (1987). "Une novelle interprétation de Longisquama insignis, reptile énigmatique du Trias supérieur d'Asie centrale [A new interpretation of Longisquama insignis, an enigmatic reptile from the Upper Triassic of Central Asia]". Comptes Rendus de l'Académie des Sciences de Paris. 305 (serie II): 65–70.
- ^ a b Prum, R. O./Unwin, D.M., Benton, M.J./Response; Jones, T.D., Ruben, J.A., Maderson, P.F.A., Martin, L.D. (9 March 2001). "Longisquama Fossil and Feather Morphology". Science. 291 (5510): 1899–1902. doi:10.1126/science.291.5510.1899c. PMID 11245191. S2CID 34144897.
{{cite journal}}: CS1 maint: 여러 이름: 작성자 목록(링크) - ^ a b Jones, T.D.; Ruben, J.A.; Martin, L.D.; Kurochkin, E.; Feduccia, A.; Maderson, P.F.A.; Hillenius, W.J.; Geist, N.R.; Alifanov, V. (23 June 2000). "Nonavian Feathers in a Late Triassic Archosaur". Science. 288 (5474): 2202–2205. Bibcode:2000Sci...288.2202J. doi:10.1126/science.288.5474.2202. PMID 10864867.
- ^ Prum, R.O. (2002). "Are current critiques of the theropod origin of birds science? Rebuttal to Feduccia". The Auk. 120 (2): 550–561. doi:10.1642/0004-8038(2003)120[0550:ACCOTT]2.0.CO;2. ISSN 0004-8038.
- ^ Reisz, R.R.; Sues, H.-D. (23 November 2000). "The "Feathers" of Longisquama". Nature. 408 (6811): 428. doi:10.1038/35044204. PMID 11100716. S2CID 4425631.
- ^ Fraser, N. (2006). Dawn of The Dinosaurs: Life in the Triassic. Bloomington: Indiana University Press. ISBN 978-025-334-3.
- ^ a b c d e Buchwitz, M.; Voigt, S. (2012). "The dorsal appendages of the Triassic reptile Longisquama insignis: reconsideration of a controversial integument type". Paläontologische Zeitschrift. 86 (3): 313–331. doi:10.1007/s12542-012-0135-3. S2CID 84633512.
- ^ Sharov, A.G. (1970). "A peculiar reptile from the lower Triassic of Fergana". Paleontologicheskii Zhurnal (1): 127–130.
- ^ 올셰프스키, G. 19911869년 고등 크로코딜리아를 제외한 파랭프라클래스 아크오사우루스 코프 개정판.중생대 미안더링스, 2:196pp.
- ^ Senter, P. (2004). "Phylogeny of the Drepanosauridae (Reptilia: Diapsida)". Journal of Systematic Palaeontology. 2 (3): 257–268. doi:10.1017/S1477201904001427. S2CID 83840423.
- ^ Padian, Kevin (2004). "Basal Avialae". In Weishampel, David B.; Dodson, Peter; Osmólska Halszka (eds.). The Dinosauria (Second ed.). Berkeley: University of California Press. pp. 210–231. ISBN 0-520-24209-2.
추가 정보
- Feduccia, A.; Lingham-Soliar, T. & Hinchliffe, J.R. (2005). "Do feathered dinosaurs exist? Testing the hypothesis on neontological and paleontological evidence". Journal of Morphology. 266 (2): 125–66. doi:10.1002/jmor.10382. PMID 16217748. S2CID 15079072.
- Martin, L. D. (2008). "Origin of avian flight- a new perspective". Oryctos. 7: 45–54.
- Peters, D. (2000). "A Redescription of Four Prolacertiform Genera and Implications for Pterosaur Phylogenesis". Rivista Italiana di Paleontologia e Stratigrafia. 106 (3): 293–336.
- Peters, D. (2002). "A New Model for the Evolution of the Pterosaur Wing – with a twist". Historical Biology. 15 (4): 277–301. doi:10.1080/08912960127805. S2CID 84295762.
- Stokstad E. (23 June 2000). "Feathers, or flight of fancy?". Science. 288 (5474): 2124–2125. doi:10.1126/science.288.5474.2124. PMID 10896578. S2CID 82060725.
- Sebastian Voigt; Michael Buchwitz; Jan Fischer; Daniel Krause & Robert Georgi (2009). "Feather-like development of Triassic diapsid skin appendages". Naturwissenschaften. 96 (1): 81–86. Bibcode:2009NW.....96...81V. doi:10.1007/s00114-008-0453-1. PMID 18836696. S2CID 10104426.
외부 링크
- 미키 모티머의 마틴 2004 비평
- 미키 모티머의 페두치아 외 2005년 비평
- 쿠로치킨과 마틴이 발견한 롱기스콰마 플룸에 관한 신문 기사
- Jones 등으로부터 받은 모식 표본의2000
- Nicolay Gnezdilov의 트위터 계정에서 모식표본 사진
- Christian Kamerer의 트위터 계정에서 모식표본 사진
사우롭시다(렙틸리아센스라토) | |||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||