BRCA1
BRCA1






유방암 제1형 감수성 단백질은 BRCA1(/ (brékˈʌw)n/) [5]유전자에 의해 인간에서 코드되는 단백질이다.척추동물의 유전체는 다른 척추동물 종에서 흔히 볼 수 있는 반면 무척추동물의 게놈은 더 먼 친척의 [6]유전자를 부호화할 수 있다.BRCA1은 인간 종양 억제 유전자[7][8](관리 유전자라고도 함)로 DNA를 [9]수리하는 역할을 한다.
BRCA1과 BRCA2는 관련이 없는 [10]단백질이지만, 둘 다 보통 유방과 다른 조직의 세포에서 발현되며, 손상된 DNA를 복구하거나 DNA가 복구될 수 없다면 세포를 파괴하는 것을 돕는다.이들은 염색체 손상 복구에 관여하며 DNA 이중 가닥 절단의 [11][12]오류 없는 복구에 중요한 역할을 한다.BRCA1이나 BRCA2 자체가 BRCA 돌연변이에 의해 손상되면 손상된 DNA가 제대로 복구되지 않아 유방암에 [13][14]걸릴 위험이 높아진다.BRCA1과 BRCA2는 "유방암 감수성 유전자"와 "유방암 감수성 단백질"로 설명되었다.지배적인 대립 유전자는 정상적이고 종양 억제 기능을 가지고 있는 반면, 이러한 유전자의 높은 투과성 돌연변이는 유방암의 [15]증가 위험과 관련이 있는 종양 억제 기능의 상실을 야기합니다.
BRCA1은 다른 종양 억제제, DNA 손상 센서 및 신호 변환기와 결합하여 BRCA1-관련 게놈 감시 복합체(BASC)[16]로 알려진 대규모 다중 서브유닛 단백질 복합체를 형성합니다.BRCA1 단백질은 RNA 중합효소 II 및 C 말단 도메인을 통해 히스톤 탈아세틸화효소 복합체와도 상호작용한다.따라서, 이 단백질은 전사에 역할을 하며, 이중 가닥 DNA의[14] DNA 복구는 유비쿼티네이션, 전사 조절 및 다른 [17]기능을 파괴한다.
BRCA1과 BRCA2에 돌연변이가 있는 환자의 암 발생 가능성을 검사하는 방법은 Miriad [18][19]Genetics가 소유하거나 통제하는 특허에 의해 다루어졌다.1994년에 진단 시험 독점적으로 Myriad하는을 제공하 Myriad의 사업 모델을 시작 1200직원과달러 500M 연간 매출액은 2012년에, 상장 회사;[20]그것은 또한 높은 물가로 인해 논란과 이것이 이번에는으로 이어졌다 다른 진단 실험실,에서 두번째 의견을을 가져오는 것을 이끌었다.l그리고 마크 분자병리학회 대 미리어드 유전학 소송.[21]
검출
유방암 감수성과 관련된 DNA 복구 효소를 코드하는 유전자의 존재에 대한 첫 번째 증거는 1990년 [22]UC 버클리 소재 메리-클레어 킹 연구소에 의해 제공되었다.4년 후,[23] 그것을 찾기 위한 국제적인 경쟁 끝에, 이 유전자는 유타 대학, 국립환경보건과학연구소, 그리고 미리어드 [18][24]유전학 연구자들에 의해 1994년에 복제되었다.
유전자 위치
인간 BRCA1 유전자는 염기쌍 41,196,312에서 염기쌍 41,277,500(구축 GRCh37/hg19)(맵)[25]까지 영역 2 밴드 1에서 염색체 17의 긴 팔(q)에 위치한다.BRCA1 정형어는 완전한 게놈 데이터를 이용할 [6]수 있는 대부분의 척추동물에서 확인되었다.
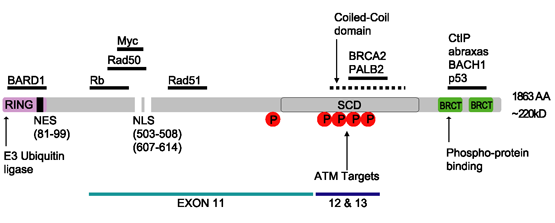
단백질 구조
BRCA1 단백질에는 다음 [26]도메인이 포함됩니다.
이 단백질은 핵 국재 신호와 핵 수출 신호 [27]모티브도 포함하고 있다.
인간 BRCA1 단백질은 Znf C3HC4-RING 도메인, BRCA1 세린 도메인 및 2개의 BRCT 도메인 등 4개의 주요 단백질 도메인으로 구성된다.이러한 도메인은 BRCA1 단백질의 약 27%를 인코딩합니다.BRCA1에는 [28]6개의 알려진 이소형식이 있으며, 이소형식 1과 2는 [citation needed]각각 1863개의 아미노산으로 구성됩니다.
BRCA1은 BRCA2와 관련이 없다. 즉, 호몰로그나 [10]패럴로그가 아니다.
아연 약지 도메인
진핵생물 펩타이드에서 발견되는 Zn 핑거인 RING 모티브는 40~60개의 아미노산 길이로 보존된 8개의 금속 결합 잔류물, 두 개의 아연 [30]원자를 조정하는 시스테인 또는 히스티딘 잔류물 2개 쿼텟으로 구성된다.이 모티브는 짧은 반평행 베타 시트, 두 개의 아연 결합 루프 및 작은 영역의 중앙 알파 나선을 포함한다.이 RING 도메인은 또한 RING 모티브를 포함하는 BARD1을 포함한 관련 단백질과 상호작용하여 헤테로디머를 형성한다.BRCA1 RING 모티브는 BRCA1 단백질의 잔류물 8~22와 81~96에 의해 형성된 알파 헬리크로 측면 형성된다.또한 잔류물 36–48과 101–116에서 형성된 두 개의 알파-헬리크로 측면의 RING 핑거로 구성된 BARD1의 상동 영역과 상호작용한다.이들 4개의 나선은 결합되어 헤테로다이머화 계면을 형성하고 BRCA1-BAR1 헤테로다이머 복합체를 안정화한다.측면 영역의 인접 잔류물과 소수성 상호작용 간의 상호작용에 의해 추가적인 안정화가 달성된다.BARD1/BRCA1 상호작용은 BRCA1의 종양유전성 아미노산 치환에 의해 파괴되며, 이는 이들 단백질 사이의 안정적인 복합체 형성이 BRCA1 종양 억제의 [30]필수적인 측면일 수 있음을 암시한다.
고리 도메인은 단백질 유비퀴티네이션을 촉매하는 유비퀴틴 E3 연결효소의 중요한 요소입니다.유비퀴틴은 모든 조직에서 발견되는 작은 조절 단백질로 단백질을 세포 내의 구획으로 유도한다.특히 BRCA1 폴리펩타이드, Lys-48 연결 폴리유비퀴틴 사슬은 휴지 세포핵 전체에 분산되어 있지만 DNA 복제 개시 시에는 BRCA2 및 BARD1을 포함한 억제된 그룹으로 모인다.BARD1은 유비퀴타이션을 [31]위한 단백질 타깃의 인식 및 결합에 관여하는 것으로 생각된다.단백질에 달라붙어 파괴의 꼬리표를 붙인다.유비퀴티네이션은 BRCA1 융합 단백질을 통해 발생하며 아연 킬레이트화에 의해 [30]폐지된다.융합 단백질의 효소 활성은 고리 [citation needed]영역의 적절한 접힘에 따라 달라집니다.
Serine 클러스터 도메인
BRCA1 세린 클러스터 도메인(SCD)은 아미노산 1280–1524에 걸쳐 있다.도메인의 일부는 exon 11 ~13에 있습니다.높은 돌연변이율은 엑손 11-13에서 발생한다.보고된 BRCA1의 인산화 부위는 시험관내 및 생체내 ATM/ATR 키나제에 의해 인산화된다.ATM/ATR은 DNA 손상에 의해 활성화된 키나아제입니다.세린 잔기의 돌연변이는 DNA 손상 부위 및 DNA 손상 반응 [29]기능에 대한 BRCA1의 국재화에 영향을 미칠 수 있다.
BRCT 도메인
BRCA1 단백질의 이중 반복 BRCT 도메인은 길이가 약 70Ω, [32]폭이 30~35Ω인 가늘고 긴 구조이다.BRCT의 85~95개의 아미노산 도메인은 단일 모듈 또는 [33]두 개의 도메인을 포함하는 다중 탠덤 반복으로 확인할 수 있다.이 두 가지 가능성 모두 다양한 형태의 [32]단일 단백질에서 발생할 수 있습니다.BRCA1 단백질의 C 말단 BRCT 영역은 DNA의 복구, 전사 조절 및 종양 억제 기능을 [34]위해 필수적이다.BRCA1에서 이중 탠덤 반복 BRCT 도메인은 3차원 구조에서 머리부터 꼬리까지 배치되어 계면에 소수성 용매 접근 가능한 표면적 1600Ω을 묻는다.이 모든 것이 인터페이스를 구성하는 꽉 막힌 노브인홀 구조의 원인이 됩니다.이러한 상동 도메인은 DNA 손상에 대한 세포 반응을 제어하기 위해 상호작용합니다.이 두 단백질의 계면에서의 미스센스 돌연변이는 세포주기를 교란시켜 암에 [citation needed]걸릴 위험이 더 크다.
기능 및 메커니즘
BRCA1은 DNA의 이중 가닥 파손을 복구하는 복합체의 일부이다.DNA 이중나선의 가닥은 손상되면서 지속적으로 끊어지고 있다.한 가닥만 끊어질 때도 있고 두 가닥이 동시에 끊어질 때도 있습니다.DNA 가교제는 염색체/DNA 손상의 중요한 원천이다.이중사슬 절단은 가교 제거 후 중간체로서 발생하며, 실제로 BRCA1의 바이알렐 돌연변이는 DNA 가교제에 대한 과민증과 관련된 유전병인 [35]판코니 빈혈의 원인이 되는 것으로 확인되었다.BRCA1은 두 가닥이 끊어졌을 때 DNA를 복구하는 단백질 복합체의 일부이다.이 경우 복구 메커니즘이 올바른 DNA 염기서열을 대체하는 방법을 아는 것은 어렵고 복구를 시도하는 방법은 여러 가지가 있습니다.BRCA1이 참여하는 이중 가닥 복구 메커니즘은 호몰로지 방향 복구이며, 여기서 복구 단백질은 온전한 [36]자매 염색체로부터 동일한 염기서열을 복제한다.
많은 유형의 정상 세포의 핵에서, BRCA1 단백질은 DNA 이중 가닥 [37]절단 수복 중에 RAD51과 상호작용합니다.이러한 파손은 자연 방사선이나 다른 피폭에 의해 발생할 수 있지만, 염색체가 유전 물질을 교환할 때(예: 감수 분열 중 "교차") 발생한다.BRCA1과 유사한 기능을 가진 BRCA2 단백질도 RAD51 단백질과 상호작용한다.DNA 손상 복구에 영향을 줌으로써, 이 세 가지 단백질은 인간 [citation needed]게놈의 안정성을 유지하는 역할을 한다.
BRCA1은 불일치 복구라고 불리는 또 다른 유형의 DNA 복구에도 관여합니다.BRCA1은 DNA 미스매치 복구 단백질 MSH2와 [38]상호작용한다.MSH2, MSH6, PARP 및 단사슬 복구에 관여하는 다른 단백질은 BRCA1 결손 유선종양에서 [39]상승하는 것으로 보고되고 있다.
발로신 함유 단백질이라고 불리는 단백질은 손상된 DNA 부위에 BRCA1을 모집하는 역할을 한다.전리방사선 후 VCP를 DNA 병변으로 끌어들여 유비퀴틴 리가아제 RNF8과 협력하여 효율적인 DSB [40]수복을 위한 시그널링 복합체 조립을 조정한다.BRCA1은 [41]VCP와 상호작용합니다.BRCA1은 또한 c-Myc 및 게놈 [42]안정성을 유지하기 위해 중요한 다른 단백질과도 상호작용합니다.
BRCA1은 분지 DNA 구조에 대해 높은 친화력을 가지고 DNA에 직접 결합한다.DNA에 결합하는 이러한 능력은 Mre11만의 [43]핵산가수분해효소 활성뿐만 아니라 MRN 복합체의 핵산가수분해효소 활성도 억제하는 능력에 기여한다.이는 BRCA1이 비호몰로지 엔드 결합(NHEJ)[44]에 의한 저충실도 DNA 수복을 촉진하는 역할을 설명할 수 있다.BRCA1은 또한 DNA 이중사슬 단절 복구 포치에서 γ-H2AX(세린-139에서 인산화되는 히스톤 H2AX)와 함께 열원소화되며, [17][45]이는 복구 인자를 모집하는 데 역할을 할 수 있음을 나타낸다.
포름알데히드와 아세트알데히드는 [46]흔히 경로를 포함하는 BRCA1에 의해 매개되는 DNA 교차 연결의 일반적인 환경 소스입니다.
이 DNA 복구 기능은 필수적이다. 두 BRCA1 대립 유전자에 기능 돌연변이가 있는 생쥐는 생존할 수 없으며, 2015년 현재 두 대립 유전자에 기능 돌연변이가 있는 성인은 두 명뿐이며, 선천적 또는 발달적 문제가 있으며, 둘 다 암에 걸렸다.하나는 BRCA1 돌연변이 중 하나가 [47]하이포메이션이었기 때문에 성인기까지 생존한 것으로 추정되었다.
문자 변환
BRCA1은 Hela 추출물에서 인간 RNA 중합효소 II 홀로엔자임과 함께 정제되는 것으로 나타나 홀로엔자임의 [48]성분임을 시사했다.그러나 이후 연구는 이 가정을 반박하고 대신 HeLa 세포에서 BRCA1을 포함한 주요 복합체는 SWI/[49]SNF를 포함하는 2메가달톤 복합체이며, SWI/SNF는 크로마틴 리모델링 복합체라는 것을 보여주었다.염색질에 대한 BRCA1의 인위적 테더링은 [45]이 역할에 SWI/SNF 상호작용 도메인이 필요하지 않았지만 헤테로크로마틴을 축합하는 것으로 나타났다.BRCA1은 NELF [45]복합체의 NELF-B(COBRA1) 서브유닛과 상호작용한다.
돌연변이 및 암 위험

BRCA1 유전자의 특정 변이는 유전성 유방암-이온성 암 증후군의 일부로서 유방암 위험을 증가시킨다.연구자들은 BRCA1 유전자에서 수백 개의 돌연변이를 확인했는데, 그 중 다수는 암의 위험 증가와 관련이 있다.비정상적인 BRCA1 또는 BRCA2 유전자를 가진 여성은 90세까지 유방암에 걸릴 위험이 최대 80%에 이른다. 난소암 발병 위험은 BRCA1 돌연변이를 가진 여성의 경우 약 55%, BRCA2 [51]돌연변이를 가진 여성의 경우 약 25%이다.
이러한 돌연변이는 하나 또는 소수의 DNA 염기쌍(DNA의 구성 요소)의 변화일 수 있으며 PCR 및 DNA 염기서열 [citation needed]분석으로 식별될 수 있다.
경우에 따라서는 DNA의 큰 부분이 재배치됩니다.큰 재배치라고도 불리는 이러한 큰 부분들은 유전자의 하나 또는 여러 엑손의 결실 또는 복제일 수 있다.돌연변이 검출(시퀀싱)을 위한 고전적인 방법으로는 이러한 유형의 [52]돌연변이를 밝힐 수 없다.기존의 정량적 PCR,[53] Multiplex Lection-Dependent Probe Amplification(MLPA;[54] 다중화 프로브 증폭) 및 Quantitative Multiplex PCR of Short Fluorects(QMPSF;[55] 짧은 형광 조각의 정량적 다중화 PCR) 등 다른 방법이 제안되었습니다.새로운 방법: 다모양 전기영동에 의한 헤테로듀플렉스 분석(HDA) 또는 비교 유전체 혼성(array-CGH)[56]에 기초한 전용 올리고뉴클레오티드 어레이도 최근 제안되었다.
일부 결과에 따르면 일부 암에서 보고된 BRCA1 프로모터의 과메틸화는 BRCA1 [57]발현 불활성화 메커니즘으로 간주될 수 있다.
변이된 BRCA1 유전자는 보통 제대로 기능하지 않는 단백질을 만든다.연구자들은 결함이 있는 BRCA1 단백질이 다른 유전자의 돌연변이를 일으키는 DNA 손상을 고치는 데 도움을 줄 수 없다고 믿고 있다.이러한 돌연변이는 축적될 수 있고 세포들이 통제할 수 없이 자라고 분열하여 종양을 형성하게 할 수 있다.따라서, BRCA1 불활성화 돌연변이는 [citation needed]암에 대한 소인으로 이어진다.
BRCA1 mRNA 3' UTR은 miRNA, Mir-17 마이크로RNA와 결합할 수 있다.Mir-30 마이크로RNA와 함께 이 miRNA의 변이가 유방암에 [58]걸리기 쉽다고 제안되었다.
유방암 외에도 BRCA1 유전자의 돌연변이는 난소암과 전립선암의 위험도 높인다.또한 팔로피아 튜브 내의 전암성 병변(이형성증)은 BRCA1 유전자 돌연변이와 관련이 있다.BRCA1과 BRCA2를 포함하는 모델 경로에서 병원성 돌연변이는 백혈병과 림프종의 [14]하위 집합에 대한 위험을 크게 증가시킨다.
결함이 있는 BRCA1 또는 BRCA2 유전자를 물려받은 여성들은 유방암과 난소암에 걸릴 위험이 매우 높다.유방암이나 난소암에 걸릴 위험이 매우 높고 암에만 특이해 많은 돌연변이 보균자들이 예방 수술을 받기를 선택한다.그렇게 눈에 띄는 조직의 특이성을 설명하는 데는 많은 추측이 있었다.BRCA 1/2 유전성 암이 발생하는 주요 결정요인은 만성 염증을 일으키는 물질인 암 병원체의 조직 특이성과 관련이 있다.대상 조직은 병원체에 대한 수용체를 가질 수 있으며 염증 과정 또는 발암 물질에 선택적으로 노출될 수 있습니다.종양억제유전자의 선천적인 유전체결핍은 정상반응을 저해하고 장기표적에서 질병에 대한 감수성을 악화시킨다.이 이론은 또한 BRCA1 또는 BRCA2를 초과하는 여러 종양 억제제에 대한 데이터에도 적합하다.이 모델의 주요 장점은 예방 [59]수술 외에 몇 가지 옵션이 있을 수 있다는 것입니다.
유방암과 난소암에서 BRCA1의 낮은 발현
BRCA1 발현은 대부분의 고악도 유관성 [60]유방암에서 감소되거나 검출되지 않는다.BRCA1 활성의 상실은 생식계 돌연변이 또는 유전자 발현의 하향 조절에 의해 특정 표적 조직에서 종양 형성으로 이어진다는 것은 오랫동안 알려져 왔다.특히 BRCA1 발현 감소는 산발성 및 유전성 유방종양의 [61]진행에 모두 기여한다.BRCA1의 감소된 발현은 상동 재조합의 [62]잠재적 무오류 경로에 의한 DNA 손상, 특히 이중 가닥 파괴의 복구에 중요한 역할을 하기 때문에 종양 유발이다.BRCA1 단백질이 부족한 세포는 보다 오류가 발생하기 쉬운 대체 메커니즘에 의해 DNA 손상을 복구하는 경향이 있기 때문에, 이 단백질의 감소 또는 침묵은 유방암으로 [62]진행될 수 있는 돌연변이와 총염색체 재배열을 생성한다.
마찬가지로, EOC가 난소암의 가장 흔한 유형으로 난소암의 약 90%를 차지하는 산발성 상피 난소암([63]EOCs)의 과반수(55%)에서 BRC1 발현이 낮다.EOCs의 약 3분의 2를 구성하는 하위 범주인 장액성 난소암종에서 낮은 BRCA1 발현이 50%[64] 이상에서 발생한다.Bowtell은[65] BRCA1 결핍으로 인한 상동 재조합 수리가 부족하다는 것을 나타내는 문헌을 검토했다.특히 이 결핍은 고농도의 장액성 난소암의 진화를 조각하고 치료에 대한 반응을 지시하는 일련의 분자적 사건을 일으킨다.특히 BRCA1 결핍은 BRCA1 돌연변이 또는 BRCA1 발현 결핍을 유발하는 다른 사건으로 인해 종양 발생의 원인이 될 수 있다는 점에 주목했다.
유방암과 난소암의 BRCA1 돌연변이
유방암에 걸린 모든 여성의 약 3%-8%만이 BRCA1 또는 BRCA2에 [66]돌연변이를 가지고 있다.마찬가지로, BRCA1 돌연변이는 난소암의 약 18%에서만 볼 수 있다(배아세포 돌연변이 13%, 체세포 [67]돌연변이 5%).
따라서 이러한 암의 대부분에서 BRCA1 발현이 낮은 반면, BRCA1 돌연변이는 발현 감소의 주요 원인이 아니다.유방암 종양에서 자주 발견되는 특정 잠복 바이러스는 BRCA1 유전자의 발현을 감소시켜 유방암의 [68]발생을 유발할 수 있다.
유방암과 난소암의 BRCA1 프로모터 과메틸화
BRCA1 프로모터 과메틸화는 선택되지 않은 1차 유방암의 [69]13%에서만 나타났다.마찬가지로, BRC1 프로모터 과메틸화는 EOC [63]사례의 5% ~ 15%에서만 존재했다.
따라서 이러한 암에서 BRCA1 발현이 낮은 반면, BRCA1 프로모터 메틸화는 발현 감소의 사소한 원인일 뿐이다.
유방암에서 BRCA1의 MicroRNA 억제
과도하게 발현되면 특정 DNA 복구 단백질의 발현을 직접적으로 감소시키는 많은 특정 마이크로RNA가 있다(MicroRNA 섹션 DNA 복구 및 암 참조). 유방암의 경우, 마이크로RNA-182(miR-182)가 특히 BRCA1을 [70]대상으로 한다.유방암은 수용체 상태 또는 조직학에 따라 분류할 수 있으며, 삼중 음성 유방암(유방암의 15%–25%), HER2+(유방암의 15%–30%), ER+/PR+(유방암의 약 70%), 침습성 소엽암(침습성 유방암의 약 5%–10%)이 있다.4종류의 유방암 모두 정상 유방조직에 비해 [71]miR-182가 평균 약 100배 증가한 것으로 나타났다.유방암 세포주에서는 BRCA1 단백질 [70]수치와 miR-182 발현과의 역상관관계가 있다.따라서 고악도관성 유방암에서 BRCA1의 감소 또는 부재는 과다 발현된 miR-182에 기인할 수 있다.
miR-182 외에 miR-146a 및 miR-146b-5p 한 쌍의 거의 동일한 마이크로RNA도 BRCA1 발현을 억제한다.이 두 개의 마이크로RNA는 삼중 음성 종양에서 과잉 발현되어 BRCA1 [72]불활성화를 초래한다.따라서 miR-146a 및/또는 miR-146b-5p도 이러한 3중 음성 유방암에서 BRCA1의 발현 감소에 기여할 수 있다.
난소암에서 BRCA1의 마이크로RNA 억제
장액세관내암(고급장액난소암(HG-SOC)의 전구병변)과 HG-SOC 자체에서 약 70%의 사례에서 [73]miR-182가 과압된다.miR-182가 과다 발현된 세포에서 BRCA1은 이온화 방사선에 피폭된 후에도 낮은 수준을 유지했다(일반적으로 BRCA1 [73]발현을 증가시킨다).따라서 HG-SOC에서의 BRCA1의 감소 또는 부재는 과잉 발현 miR-182에 의한 것일 수 있습니다.
난소암세포에서 BRCA1의 발현을 감소시키는 것으로 알려진 또 다른 마이크로RNA는 miR-9이다.[63]2기 IIIC 또는 4기 장액 난소암(HG-SOG) 환자의 종양 58개 중 miR-9 발현과 BRCA1 [63]발현 사이에 역상관관계가 발견되었으므로 miR-9 증가가 이들 난소암에서 BRCA1 발현 감소에도 기여할 수 있다.
BRCA1 발현 결핍은 종양유전성일 가능성이 있습니다
DNA 손상은 [74]암의 주요 근본 원인인 것으로 보이며, DNA 수복의 결핍은 많은 형태의 [75]암의 근저에 있는 것으로 보인다.DNA 복구가 부족하면 DNA 손상이 축적되는 경향이 있다.이러한 과도한 DNA 손상은 오류가 발생하기 쉬운 전이 합성으로 인해 DNA 복제 중 돌연변이 오류를 증가시킬 수 있습니다.과도한 DNA 손상은 또한 DNA [76][77]복구 중 오류로 인한 후생유전학적 변화를 증가시킬 수 있다.그러한 돌연변이와 후생유전적 변화는 암을 유발할 수 있다.유방암과 난소암에서 빈번한 마이크로RNA 유도 BRCA1 결핍은 이러한 암의 진행에 기여할 수 있다.
생식계 돌연변이와 창시자 효과
현재까지 확인된 모든 배아줄 BRCA1 돌연변이는 유전되었고, 이는 특정 돌연변이가 잘 정의된 모집단 그룹에 공통적이고 이론적으로 공통의 조상으로 거슬러 올라갈 수 있는 큰 "창시자" 효과의 가능성을 시사한다.BRCA1에 대한 돌연변이 검사의 복잡성을 고려할 때, 이러한 일반적인 돌연변이는 특정 모집단에서 돌연변이 검사에 필요한 방법을 단순화할 수 있다.높은 빈도로 발생하는 돌연변이를 분석하면 임상 발현에 대한 [78]연구도 가능하다.창시자 효과의 징후는 아슈케나지 유대인들 사이에서 볼 수 있다.BRCA1의 세 가지 돌연변이는 유전 BRCA1 관련 유방 및/또는 난소암을 가진 아슈케나지 유대인 환자의 대다수를 설명하는 것으로 보고되었다: BRCA1 [79][80]유전자의 185delAG, 188del11 및 5382insC.실제로 유대인 여성이 BRCA1 185delAG, BRCA1 5382insC 설립자 돌연변이를 갖고 있지 않으면 다른 BRCA1 돌연변이가 [81]발견될 가능성은 매우 낮은 것으로 나타났다.BRCA1의 설립자 돌연변이의 추가 예는 표 1(주로 에서 도출[78])에 제시되어 있다.
| 모집단 또는 부분군 | BRCA1 [82]변환 | 레퍼런스 |
|---|---|---|
| 흑인 | 943ins10, M1775R | [83] |
| 아프리카인 | E881X, 1374delC | [84][85] |
| 아슈케나지 유대인 | 185delAG, 188del11, 5382inc | [79][80] |
| 오스트리아인 | 2795delA, C61G, 5382insC, Q1806 스톱 | [86] |
| 벨기에인 | 2804delAA, IVS5+3A>g | [87][88] |
| 네덜란드 | Exon 2 삭제, Exon 13 삭제, 2804delAA | [87][89][90] |
| 핀 | 3745delT, IVS11-2A>g | [91][92] |
| 프랑스 | 3600del11, G1710X | [93] |
| 프랑스계 캐나다인 | C4446T | [94] |
| 독일인들. | 5382inC, 4184del4 | [95][96] |
| 그리스인 | 5382inC | [97] |
| 헝가리인 | 300T>G, 5382inC, 185delAG | [98] |
| 이탈리아인 | 5083del19 | [99] |
| 일본인입니다 | L63X, Q934X | [100] |
| 북미 원주민 | 1510insG, 1506A>g | [101] |
| 북아일랜드어 | 2800delAA | [102] |
| 노르웨이인 | 816delGT, 1135insA, 1675delA, 3347delAG | [103] |