ABC proteini
| ABC Transporter | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Vitamin B12 transporter, BtuCD PDB 1l7v | |||||||||
| Identifikatori | |||||||||
| Simbol | ABC_tran | ||||||||
| Pfam | PF00005 | ||||||||
| InterPro | IPR003439 | ||||||||
| PROSITE | PDOC00185 | ||||||||
| SCOP | 1b0u | ||||||||
| SUPERFAMILY | 1b0u | ||||||||
| TCDB | 3.A.1 | ||||||||
| OPM superfamilija | 17 | ||||||||
| OPM protein | 3g5u | ||||||||
| |||||||||


ABC proteini (ATP-vezujući kasetni transporteri, ABC transporteri) članovi su proteinske superfamilije, koja je jedna od najvećih i najstarijih familija sa predstavnicima u svim postojećim taksonomskim razdelima, od prokariota do ljudi.[1][2] ABC transporteri se obično sastoje od višestrukih podjedinica, jedna ili dve od kojih su transmembranski proteini i jedna ili dve su za membranu vezane ATPaze. ATPazne podjedinice koriste energiju vezivanja i hidrolize adenozin trifosfata (ATP) za translokaciju raznih supstrata kroz membrane, bilo za preuzimanje ili eksport supstrata. Većina, mada ne svi sistemi takođe imaju ekstracitoplazmični receptor, protein za koji se vezuje rastvorak. Neke homologne ATPaze funkcionišu u procesima koji nisu vezani za transport, kao što je translacija RNK i popravka DNK.[3][4]
Smatra se da su ABC transporteri deo ABC superfamilije. To gledište je bazirano na njihovim proteinskim sekvencama i na organizaciji njihovih ATP-vezujućih kasetnih (ABC) domena, iako su integralni membranski proteini verovatno nezavisno evoluirali neko puta, i stoga postoji nekoliko različitih proteinskih familija. Postoje indikacije da su integralni membranski proteini ABC eksportera nezavisno evoluirali bar tri puta.[5] ABC1 eksporteri su nastali intragenskom triplikacijom 2 TMS prekurzora čime je formirano 6 TMS proteina. ABC2 eksporteri su nastali intragenskom duplikacijom 3 TMS prekurzora, i ABC3 eksporteri su nastali iz 4 TMS prekurzora koji je dupliran bilo ekstragenski što daje dva 4 TMS proteina, oba od kojih su neophodna za transportnu funkciju, ili intragenski što daje 8 ili 10 TMS proteina. Tih 10 TMS proteina imaju dva dodatna TMS domena između dve 4 TMS ponavljajuće jedinice.[6] Slično tome, moguće je da su integralni membranski proteini ABC sistema preuzimanja nezavisno nastali bar tri puta, sudeći po njihovim trodimenzionalnim strukturama visoke rezolucije.[7] ABC unosni prenosioci transportuju mnoštvo različitih nutrijenata, biosintetičkih prekurzora, retkih metala i vitamina, dok eksporteri transportuju lipide, sterole, lekove i mnoštvo različitih primarnih i sekundarnih metabolita. Neki od tih eksportera kod ljudi doprinose otpornosti tumora, cističnoj fibrozi i opsegu drugih ljudskih naslednih bolesti. Visok nivo izražavanja gena koji kodiraju pojedine eksportere kod prokariotskih i eukariotskih organizama (uključujući ljude) dovodi do razvića otpornosti na višestruke lekove, kao što su antibiotici i anti-kancerni agenti.
Poznate su stotine ABC transportera kod prokariota i eukariota.[8] ABC geni su esencijalni za mnoge procese u ćeliji, i mutacije ljudskih gena uzrokuju ili doprinose nizu ozbiljnih genetičkih bolesti.[9] Četrdeset osam ABC gena je poznato kod ljudi. Mnogi od njih su okarakterisani i pokazano je da su uzročno povezani sa bolestima kao što je cistična fibroza, adrenoleukodistrofija, Stargardtova bolest, tumori otporni na lekove, Dubin-Džonsonov sindrom, Bilerova bolest, progresivna familijarna intrahepatička holestaza, X-vezana sideroblastna anemija, ataksija, i perzistentna i hiperinsulimenska hipoglicemija.[8] ABC transporteri isto tako učestvuju u otpornosti na višestruke lekove i iz tog razloga su neki od njih prvobitno identifikovani. Kad su ABC transportni proteini prekomerno izraženi u ćelijama raka, oni mogu da eksportuju antikancerne lekove, što čini tumore rezistentnim.[10]
Funkcija
[уреди | уреди извор]ABC transporteri koriste energiju ATP vezivanja i hidrolize za transport raznih substrata kroz ćelijske membrane. Oni se dele u tri glavne funkcionalne kategorije. Kod prokariota, importeri posreduju unos nutrijenata u ćeliju. Supstrati koji mogu biti transportovani su joni, aminokiseline, peptidi, šećeri, i drugi molekuli, koji su uglavnom hidrofilni. Region ABC proteina koji premošćava membranu štiti hidrofilne supstrate od lipida iz membranskog dvosloja čime otvara put kroz ćelijsku membranu. Eukariote ne poseduju importere. Eksporteri ili eflukseri, koji su prisutni kod prokariota i eukariota, funkcionišu kao pumpe koje istiskuju toksine i lekove iz ćelije. Kod gram-negativnih bakterija, eksporteri transportuju lipide i pojedine polisaharide iz citoplazme do periplazme. Treća podgrupa ABC proteina nisu transporteri, nego proteini koji učestvuju u translaciji i procesima popravke DNK.[3]
Prokariotski ABC proteini
[уреди | уреди извор]Bakterijski ABC transporteri su esencijalni za ćelijsku vitalnost, virulenciju, i patogenost.[3] Na primer, ABC sistemi za preuzimanje gvožđa su važni efektori virulencije.[11] Patogeni koriste siderofore, kao što je enterobaktin, da dođu do gvožđa koje je u kompleksu sa proteinima koji ga vezuju sa visokim afinitetom ili eritrocitima. Oni su gvožđe-helatni molekuli visokog afiniteta koje izlučuju bakterije, i koji se zatim reapsorbuju u obliku gvožđe-siderofor kompleksa. ChvE-gguAB gen u Agrobacterium tumefaciens kodira importere glukoze i galaktoze koji takođe doprinose virulenciji.[12][13] Transporteri su od vitalne važnosti za opstanak ćelija. Oni funkcionišu kao proteinski sistemi koji se suprostavljaju svim neželjenim promenama u ćeliji. Na primer, potencijalno letalno povišenje osmotske jačine se uravnotežava aktivacijom osmosenzitivnog ABC transportera koji posreduje preuzimanje rastvoraka.[14] Osim učešća u transportu, deo bakterijskih ABC proteina takođe učestvuje u regulaciji nekoliko fizioloških procesa.[3]
Opseg materija koje bakterijski efluksni sistemi iznose iz ćelije obuhvata površinske komponente bakterijskog ćelijskog zida (npr. kapsularne polisaharide, lipopolisaharide, i teihoinsku kiselinu), proteine koji učestvuju u bakterijskoj patogenezi (e.g. hemolizi: Hem-vezujući protein, i alkalne proteaze), hem, hidrolitičke enzime, proteine S-sloja, faktore kompetencije, toksine, antibiotike, bakteriocine, peptidne antibiotike, lekove i siderofore.[15] Oni takođe imaju važnu ulogu u biosintetičkim putevima, uključujući ekstracelularnu polisaharidnu biosintezu[16] i citohromne biogeneze.[17]
Eukariotski ABC proteini
[уреди | уреди извор]Mada su eukariotski ABC transporteri uglavnom efluksori, neki od njih ne učestvuju direktno u transportu supstrata. U transmembranskom regulatoru cistične fibroze (CFTR) i u receptoru sulfonilureje (SUR), ATP hidroliza je povezana sa regulacijom otvaranja i zatvaranja jonskih kanala koju izvode sami ABC proteini ili drugi proteini.[4]
Ljudski ABC transporteri učestvuju u nekoliko bolesti koje nastaju usled polimorfizama ABC gena, a retko usled potpunog gubitka funkcije pojedinačnih ABC proteina.[18] Takve bolesti obuhvataju Mendelove bolesti i kompleksne genetičke poremećaje kao što su cistična fibroza, adrenoleukodistrofija, Štargardtova bolest, Tangerova bolest, imunska deficijencija, progresivna porodična intraheptična holestaza, Dabin-Džonsonov sindrom, Pseudoksantoma elastikum, perzistentna hiperinsulinemska hipoglicemija u detinjstvu usled fokalne adenomatozne hiperplazije, X-povezana sideroblastoza i anemija, starostna makularna degeneracija, familijalna hipoapoproteinemija, Retinitis pigmentosum, distrofija konusnih štapića, i druge.[4] Ljudska ABCB (MDR/TAP) familija je odgovorna za otpornost na višestruke lekove (MDR) za mnoštvo strukturno nevezanih lekova. ABCB1 ili MDR1 P-glikoprotein takođe učestvuju u drugim biološkim procesima u kojima im je lipidni transport glavna funkcija. Utvrđeno je da posreduju sekreciju steroida aldosterona u nadbubrežnim žlezdama, i njegovu inhibiciju blokiranu migracijom dendritskih imunskih ćelija, što je verovatno povezano sa izlučivanjem PAF lipida. Poznato je da ABCB1 posreduje transport kortizola i deksametazona, ali ne i progesterona i ABCB1 transfektovanim ćelijama. MDR1 takođe može da transportuje holesterol, kratkolančane i dugolančane analoge fosfatidilholina (PC), fosfatidiletanolamina (PE), fosfatidilserina (PS), sfingomijelina (SM), i glukozilceramida (GlcCer). Multispecifični transport raznovrsnih endogenih lipida putem MDR1 transportera može da utiče na transdvoslojnu distribuciju lipida, posebno u vrstama koje su normalno predominantne na unutrašnjim membranama.[18]
Nedavno je pokazano da ABC-transporteri postoje unutar posteljice gde je njihova uloga da štite razvijajući fetus od ksenobiotika.[19]
Struktura
[уреди | уреди извор]

Zajednička karakteristika svih ABC transportera je da se sastoje od dva zasebna domena, transmembranskog domena (TMD) i za nukleotid vezujućeg domena (NBD). TMD, takođe poznat kao domen koji premošćava membranu (MSD), ili integralni membranski (IM) domen, se sastoji od alfa heliksa ugrađenih u membranski dvosloj. On prepoznaje mnoštvo substrata i podleže konformacionim promenama radi transporta supstrate kroz membranu. Sekvenca i arhitektura TMD domena je promenljiva, što odražava hemijsku raznovrsnost supstrata koji bivaju translocirani. NBD ili ATP-vezujući kasetni (ABC) domen, za razliku od njega, je lociran u citoplazmi i ima visoko konzerviranu sekvencu. NBD sadrži mesto vezivanja ATP.[20] Kod većine eksportera, N-terminalni transmembranski domen i C-terminalni ABC domain su spojeni u jedan polipeptidni lanac, organizovan u redosledu TMD-NBD-TMD-NBD, kao na primer u E. coli hemolizinskom eksporteru HlyB. Importeri imaju obrnutu organizaciju, NBD-TMD-NBD-TMD, pri čemu je ABC domen N-terminal, dok je TMD C-terminal, kao na primer kod E. coli MacB proteina odgovornog za makrolidnu otpornost.[3][4]
Strukturalna arhitetura ABC transportera se sastoji od minimalno dva TMD i dva NBD domena. Četiri individualna polipeptidna lanca uključujući dve TMD i dve NBD podjedinice se mogu kombinovati da formiraju pun transporter, kao u E. coli BtuCD[21][22] importeru koji učestvuje u preuzimanju vitamina B12. Većina eksportera, kao što je eksporter višestrukih lekova Sav1866[23] iz Staphylococcus aureus, su formirani od homodimera koji se sastoji od dva polutransportera ili monomera sa TMD spojenim sa domenom vezivanja nukleotida (NBD). Pun transporter je često neophodan da bi protein bio funkcionalan. Pojedini ABC transporteri imaju dodatne elemente koji doprinose regulatornoj funkciji ove klase proteina. Specifično, importeri imaju vezujući protein (BP) visokog afiniteta koji se specifično asocira sa supstratom u periplazmi radi isporuke odgovarajućem ABC transporteru. Eksporteri nemaju vezujući protein, ali imaju intracelularni domen (ICD) koji se pridružuje heliksima koji premošćavaju membranu i ABC domenu. Smatra se da je ICD odgovoran za komunikaciju između TMD i NBD.[20]
Transmembranski domen (TMD)
[уреди | уреди извор]Većina transportera ima transmembranske domene koji se sastoje od ukupno 12 α-heliksa, sa 6 α-heliksa po monomeru. Pošto su TMD domeni strukturno raznovrsni, pojedini transporteri imaju različiti broj heliksa (između šest i jedanaest). TM domeni se kategorišu u tri zasebna seta nabora: tip I ABC importer, tip II ABC importer i ABC eksporter. Klasifikacija importerskih nabora je bazirana na detaljnoj karakterizaciji sekvenci.[20] Nabor tipa I ABC importera je originalno primećen u ModB TM podjedinici molibdatnog transportera.[24] Taj dijagnostički nabor je takođe prisutan u MalF i MalG TM podjedinicama MalFGK2[25] i Met transporteru MetI.[26] U MetI transporteru, minimalni set od 5 transmembranskih heliksa sačinjava ovaj motiv, dok je jedan dodatni heliks prusutan u ModB i MalG. Opšta organizacija ovog motiva je „gore-dole” topologija TM2-5 heliksa koji opasuju translokacinu stazu i TM1 heliks obavijen oko spoljašnje membranske površine i u kontaktu sa drugim TM heliksima. Tip II ABC importerski motiv je uočen u dvadeset TM heliksnih domena BtuCD[21] i u Hi1471,[27] homolognom transporteru iz Haemophilus influenzae. U BtuCD, pakovanje heliksa je kompleksno. Primetan obrazac je da je TM2 heliks zauzima poziciju kroz centar podjedinice gde je blisko opasan drugim heliksima. TM5 i TM10 heliksi su locirani na TMD interfejsu. Region koji premošćava membranu ABC eksportera je organizovan u dva „krila” koja se sastoje od heliksa TM1 i TM2 iz jedne podjedinice i TM3-6 iz druge, u aranžmanu domenske razmene. Prominentni obrazac je da su heliksi TM1-3 srodni sa TM4-6 u smislu približne dvostruke rotacije oko ose u ravni membrane.[20]
Domen vezivanja nukleotida (NBD)
[уреди | уреди извор]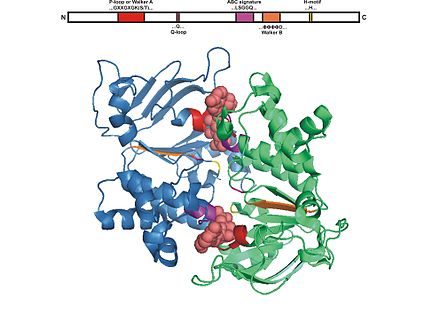
ABC domen se sastoji od dva domena, kataličkog sržnog domena poput RecA sličnih ATPaznih motora i manjeg, strukturno raznovrsnog α-heliksnog potdomena koji je jedinstven za ABC transportere. Veći domen se tipično sastoji od dve β-ravni i šest α heliksa, gde je smešten katalitički Vokerov A motif (GXXGXGKS/T pri čemu je X bilo koja aminokiselina) ili P-petlje i Vokerovog B motiva (ΦΦΦΦD, gde je Φ hidrofobni ostatak). Heliksni domen se sastoji od tri ili četiri heliksa i ABC specifičnog motiva, takođe poznatog kao LSGGQ motif, poveznog peptida ili C motiva. ABC domen isto tako ima glutaminski ostatak u fleksiblnoj petlji poznatoj pod nazivom Q petlja, poklopac ili γ-fosfatni prekidač, koji povezuje TMD i ABC. Smatra se da Q petlja učestvuje u interakciji NBD i TMD, posebno u sprezi nukleotidne hidrolize i konformacionih promena TMD tokom translokacije supstrata. H motiv ili prekidački region sadrži visoko konzervirani histidinski ostatak koji je takođe važan u interakciji ABC domena sa ATP. Ime ATP vezujuće kasete je izvedeno iz dijagnostičkog aranžmana savijanja ili motiva ove klase proteina nakon formiranja ATP sendviča i ATP hidrolize.[3][15][20]
ATP vezivanje i hidroliza
[уреди | уреди извор]ATP vezivanje je neophodno za formiranje dimera dva ABC domena transportera.[28] Generalno je primećeno da je ATP vezano stanje asocirano sa najekstenzivnijim interfejsom između ABC domena, dok strukture transportera bez nukleotida manifestuju konformacije sa većim razdvajanjem između ABC domena.[20] Strukture ATP vezanog stanja izolovanih NBD domena su poznate kod importera uključujući HisP,[29] GlcV,[30] MJ1267,[31] E. coli MalK (E.c.MalK),[32] T. litoralis MalK (TlMalK),[33] i eksportera poput TAP,[34] HlyB,[35] MJ0796,[36][37] Sav1866,[23] i MsbA.[38] U tim transporterima, ATP je vezan za ABC domen. Dva molekula ATP su pozicionirana u interfejsu dimera, između Vokerovog A motiva jedne podjedinice i LSGGQ motiva druge.[20] To je prvo uočeno kod Rad50[39] i u strukturama MJ0796, NBD podjedinici LolD transportera iz Methanococcus jannaschii[37] i E.c.MalK maltoznog transportera.[32] Te strukture su takođe konzistentne sa rezultima biohemijskih studija koji su pokazale da je ATP u bližem kontaktu sa ostacima P-petlje i LSGGQ motiva tokom katalize.[40]
Nukleotidno vezivanje je neophodno da bi se osigurao elektrostatički i/ili strukturni integritet aktivnog mesta i pomoglo formiranje aktivnog NBD dimera.[41] ATP vezivanje je stabilizovano sledećim interakcijama: (1) interakcija slaganja prstena konserviranih aromatičnih ostataka koji prethode Vokerovom A motivu i adenozinskog prstena ATP,[42][43] (2) vodoničnom vezama između konzerviranog lizinskog ostatka u Vokerovom A motivu i atoma kiseonika β- i γ-fosfata ATP i koordinacijom tih fosfata i pojedinih ostataka Vokerovog A motiva sa Mg2+ jonom,[30][34] i (3) γ-fosfatnom koordinacijom sa bočnim lancom serina i amidnim grupama proteinskog lanca glicinskih ostataka u LSGGQ motivu.[44] Osim toga, otatak koji sugeriše blisku spregu ATP vezivanja i dimerizacije, je konzervirani histidin u H-petlji. Taj histidin ostvaruje kontakte sa ostacima druge strane dimerskog interfejsa u Vokerovom A motivu i D petlji, konzerviranoj sekvenci koja sledi Vokerov B motiv.[32][37][39][45]
Pravilno vezivanje fosfata i pozicioniranje γ-fosfata prema napadajućem molekulu vode je neophodno za enzimatsku hidrolizu.[20] U nukleotidnom mestu vezivanja, atomi kiseonika β- i γ-fosfata ATP su stabilizovani ostacima u Vokerovom A motivu[46][47] i koordiniraju se sa Mg2+.[20] Mg2+ jon se takođe koordinira sa terminalnim aspartatnim ostatkom u Vokerovom B motivu kroz napaduajući H2O molekul .[30][31][36] Generalna baza, koja može da bude ostatak glutamata koji je susedan Vokerovom B motivu,[28][37][43] glutamin u Q-petlji,[27][33][37] ili histidin u prekidačkom regionu koji formira vodoničnu vezu sa γ-fosfatom ATP, katalizuju brzinu ATP hidrolize promovisanjem napadajuće H2O.[32][33][37][45] Precizni molekulski mehanizam ATP hidrolize je još uvek kontroverzan.[3]
Mehanizam transporta
[уреди | уреди извор]ABC transporteri su aktivni transporteri. Drugim rečima, da bi oni transportovali supstrate kroz ćelijske membrane njima je neophodna energija u obliku adenozin trifosfata (ATP). Ovi proteini koriste energiju ATP vezivanja i/ili hidrolize za izvršavanje konformacionih promena u transmembranskom domenu (TMD) i konsekventno za transport molekula.[48] ABC importeri i eksporteri imaju zajednički mehanizam transporta supstrata, što je posledica sličnosti njihovih struktura. Mehanizam koji opisuje konformacione promene usled vezivanja supstrata je model naizmeničnog pristupa. U tom modelu, mesto vezivanja supstrata naizmenično prelazi između konformacija ka spoljašnjosti i unutrašnjosti. Relativni afiniteti vezivanja supstrata te dve konformacije u znatnoj meri određuju smer transporta. Za importere, koji translociraju supstrat iz periplazme u citoplazmu, konformacija ka spoljašnjosti ima veći afinitet vezivanja supstrata. U kontrastu s tim, afinitet vezivanja supstrata eksportera je veći u konformaciji orijentisanoj prema unutrašnjosti.[20] Model koji opisuje konformacione promene u domenu vezivanja nukleotida (NBD) usled vezivanja ATP i hidrolize je model ATP-prekidača. Taj model se bazira na dve glavne NBD konformacije: formiranje zatvorenog dimera nakon vezivanja dva molekula ATP i disocijaciju u otvoreni dimer omogućenu hidrolizom ATP i oslobađanjem neorganskog fosfata (Pi) i adenozin difosfata (ADP). Prelaz između konformacija otvorenog i zatvorenog dimera indukuje konformacione promene u TMD, što dovodi do translokacije supstrata.[49]
Opšti mehanizam transportnog ciklusa ABC transportera nije u potpunosti razjašnjen mada postoji znatna količina strukturnih i biohemijskih podata koji podržavaju model u kome su ATP vezivanje i hidroliza spregnuti sa konformacionim promenama transportera. Odmarajuće stanje svih ABC transportera ima NBD domene u konfiguraciji otvorenog dimera, sa niskim afinitetom za ATP. Ta otvorena konformacija poseduje komoru koja je dostupna sa unutrašnjosti transportera. Transportni ciklus se inicira vezivanjem supstrata za mesto vezivanja visokog afiniteta na TMD, što indukuje konformacione promene u NBD domenima i pospešuje vezivanje ATP. Dva molekula ATP se kooperativno vezuju, i time se formira konfiguracija zatvorenog dimera. Zatvoreni NBD dimer indukuje konformacionu promenu u TMD domenima tako da dolazi do TMD otvaranja, i formiranja komore sa otvorom na suprotnoj strani od inicijalnog stanja. Afinitet suptrata za TMD se redukuje, te se supstrat oslobađa. Tome sledi hidroliza ATP i zatim sekvencijalno otpuštanje Pi i zatim ADP, čime se transporter vraća u prvobitnu konfiguraciju. Mada je predložen zajednički mehanizam, redosled vezivanja supstrata, nukleotida, hidrolize, i konformacionih promena, kao i interakcija između domena su još uvek predmet aktivne debate.[3][15][18][20][38][41][48][49][50][51][52][53]
Nekoliko grupa koje studiraju ABC transportere imaju različite postavke u pogledu mehanizma funkcionisanja transportera. Generalno se podrazumeva da ATP hidroliza pruža glavni energetski input ili podsticaj za transport i da NBD domeni deluju naizmenično, kao i da je moguće da učestvuju u različitim koracima transportnog ciklusa.[54] Međutim, nedavni strukturni i biohemijski podaci pokazuju da ATP vezivanje, umesto ATP hidrolize, pruža podsticaj. Pošto ATP vezivanje inicira NBD dimerizaciju, isto tako je moguće da formiranje dimera predstavlja podsticaj. Pojedini transporteri imaju NBD domene koji nemaju slične sposobnosti vezivanja i hidrolize ATP, i njihov interfejs NBD dimera se sastoji od dva mesta vezivanja ATP, što sugeriše konkarentnu funkciju dva NBD domena u transportnom ciklusu.[49]
Postoji izvesna evidencija da je ATP vezivanje zaista podsticaj transportnog ciklusa.[49] Pokazano je da vezivanje ATP indukuje promene svojstava vezivanja supstrata TM domena. Teško je direktno meriti afinitet ABC transportera za substrate, a indirektna merenja, na primer putem stimulacije aktivnosti ATPaze, često odražavaju druge korake koji uslovljavaju brzinu reakcije. Nedavno je pokazano, direktnim merenjem vezivanja vinblastina za permeazni-glikoprotein (P-glikoprotein) u prisustvu nehidrolizabilnih ATP analoga, e.g. 5’-adenilil-β-γ-imidodifosfata (AMP-PNP), da je ATP vezivanje u odsustvu hidrolize dovoljno da se redukuje afinitet vezivanja supstrata.[55] Isto tako, ATP vezivanje indukuje znatne konformacione promene u dva TM domena. Spektroskopske studije proteazne pristupnosti i unakrsnog vezivanja su pokazala da ATP vezivanje za NB domene indukuje konformacione promene u proteinu-1 koji je povezan sa otpornošću na višestruke lekove (MRP1),[56] HisPMQ,[57] LmrA,[58] i Pgp.[59] Dvodimenzione kristalne strukture AMP-PNP-vezanog Pgp su pokazale da se glavna konformaciona promena tokom transportnog ciklusa javlja nakon ATP vezivanja i subsekventna hidroliza ATP dovodi do ograničenih promena.[60] Rotacija i naginjanje transmembranskih α-heliksa mogu da doprinesu tim konformacionim promenama. Druge studije su bile usresređene na nalaženju dokaza da ATP vezivanje indukuje formiranje zatvorenog dimera NBD. Biohemijske studije netaknutih transportnih kompleksa sugerišu da su konformacione promene NB domena relativnom male. U odsustvu ATP, NB domeni mogu da budu relativno fleksibilni, mada ne dolazi do većih reorijentacija NB domena u odnosu na druge domene. ATP vezivanje indukuje rotacije krutih tela dva ABC potdomena jedan u odnosu na drugi, što omogućava odgovarajuće poravnavanje nukleotida u aktivnom mestu i interakciju sa namenskim motivima. Postoji jaka biohemijska evidencija da vezivanje dva ATP molekula može da bude kooperativno, drugim rečima, ATP se mora vezati na dva aktivna mesta da bi došlo do dimerizacije NB domena i formiranja zatvorene forme, koja je katalitički aktivna konformacija.[49]
ABC importeri
[уреди | уреди извор]Većina ABC transportera koji posreduju unos nutrijenata i drugih molekula u bakterije se oslanjaju na proteine vezivanja rastvorka visokog afiniteta (BP). Oni su rastvorni proteini koji su locirani u periplazmičnom prostoru između unutrašnje i spoljašnje membrane gram negativnih bakterija. Gram-pozitivni mikroorganizmi nemaju periplazmu tako da je njihov vezujući protein često lipoprotein vezan za spoljašnje lice ćelijske membrane. Neke gram-pozitivne bakterije imaju BP proteine vezane za transmembranski domen samog transportera.[3] Prva uspešna kristalna struktura netaknutog ABC importera je molibdenski transporter (ModBC-A) iz Archaeoglobus fulgidus.[24] Strukture atomske-rezolucije tri druga bakterijska importera, E. coli BtuCD,[21] E. coli maltoznog transportera (MalFGK2-E),[25] i mogućeg metal-helatnog transportera Haemophilus influenza, HI1470/1,[27] su takođe određene. Te strukture pružaju detaljne slike interakcije transmembrankih i ABC domena. One takođe otkrivaju dve različite konformacije sa otvorom u dva različita smera. Još jedno zajedničko svojstvo importera je da je svaki NBD vezan za jedan TMD prvenstveno putem kratkog citoplazmatičnog heliksa TM domena, „heliksa sprezanja”. Ta porcija EAA petlje je smeštena na površini otvora formiranog između RecA-sličnog i heliksnog ABC potdomena i leže približno paralelno sa membranskim dvoslojom.[51]
Veliki ABC importeri
[уреди | уреди извор]BtuCD i HI1470/1 su klasifikovani kao veliki ABC importeri. Transmembranska podjedinica importera vitamina B12, BtuCD, sadrži 10 TM heliksa, a funkcionalna jedinica se sastoji od dve kopije, svaka od kojih ima domen vezivanja nukleotida (NBD) i transmembranski domen (TMD). TMD i NBD formiraju interakcije jedan s drugim putem citoplazmatične petlje između dva TM heliksa i Q petlje u ABC. U odsustvu nukleotida, postoje dva ABC domena i interfejs dimera je otvoren. Poređenje struktura vezujućeg proteina sa (BtuCDF) i bez (BtuCD) otkriva da BtuCD ima otvor ka periplazmi, dok je u BtuCDF, konformacija ka spoljašnjosti zatvorena na obe strane membrane. Strukture BtuCD i BtuCD homologa, HI1470/1, predstavljaju dva različita konformaciona stanja ABC transportera. Predviđeni translokacioni put u BtuCD je otvoren ka periplazmi i zatvoren ka citoplazmi membrane, dok je kod HI1470/1 suprotan slučaj i otvor je strani citoplazme. Razlika u strukturama je u zaokretu za 9° jedne TM podjedinice relativno na drugu.[3][20][51]
Mali ABC importeri
[уреди | уреди извор]Strukture ModBC-A i MalFGK2-E, koje su u kompleksu sa njihovim vezujućim proteinom, odgovaraju malim ABC importerima. TM domeni ModBC-A i MalFGK2-E imaju samo šest heliksa po podjedinici. Homodimer ModBC-A je u konformaciji u kojoj su TM podjedinice (ModB) orijentisane u invertovanom V-obliku sa otvorom dostupnom iz citoplazme. ABC podjedinice (ModC), s druge strane, su uređene u otvorenoj konformaciji bez nukleotida, u kojoj je P-petlja jedne podjedinice naspramna, mada odvojena od LSGGQ motiva druge. Vezujući protein ModA je u zatvorenoj konformaciji sa supstratom vezanim u otvor između dva domena i vezan za ekstracelularne petlje ModB, gde supstrat sedi direktno iznad zatvorenog ulaza transportera. MalFGK2-E struktura podseća na katalitičko prelazno stanje ATP hidrolize. Ona je u zatvorenoj konformaciji u kojoj se sastoji od dva ATP molekula, u sendviču između Voker A i B motiva jedne podjedinice i LSGGQ motiva druge podjedinice. Protein vezivanja maltoze (MBP ili MalE) je smešten na periplazmatičnoj strani TM podjedinica (MalF i MalG) i velika, zaklonjena šupljina se može naći na interfejsu MalF i MalG. Aranžman TM heliksa je u konformaciji koja je zatvorena ka citoplazmi, i ima otvor ka spoljašnjoj strani. Ta struktura sugeše da je moguće da MBP stimuliše ATPaznu aktivnost transportera nakon vezivanja.[3][20][51]
Mehanizam transportnog importera
[уреди | уреди извор]
Mehanizam transporta importera podržava model naizmeničnog pristupa. Odmarajuće stanje importera je usmereno ka unutrašnjoj strani, gde je dimerni interfejs domena vezivanja nukleotida (NBD) otvoren TM domenima, mada je zaklonjen od citoplazme. Nakon dokiranja zatvorenog vezujućeg proteina sa substratom na periplazmičnoj strani transmembranskih domena, ATP se vezuje i NBD dimer se zatvara. Time se odmarajuće stanje transportera stavlja u konfomraciju prema spoljašnjoj strani u kojoj su TM domeni reorijentisani tako da mogu da prime supstrat iz vezujućeg proteina. Nakon hidrolize ATP, NBD dimer se otvara i supstrat se oslobađa u citoplazmu. Otpuštanje ADP i Pi vraća transporter u njegovo odmarajuće stanje. Jedina inkonzistentnost ovog mehanizma u odnosu na model ATP-prekidača je da je konformacija u njegovom odmarajućem stanju bez nukleotida različita od očekivane konformacije usmerene ka spoljašnjoj strani. Mada je to slučaj, ključna tačka je da se NBD ne dimerizuje ukoliko ATP i vezujući protein nisu vezani za transporter.[3][15][20][49][51]
ABC eksporteri
[уреди | уреди извор]Prokariotski ABC eksporteri su izobilni. Oni imaju bliske homologe kod eukariotima. Ova klasa transportera se klasifikuje po tipu supstrata koji transportuju. Jedna grupa učestvuje u eksportu proteina (e.g. toksina, hidrolitičkih enzima, proteina S-sloja, lantibiotika, bakteriocina, i faktora kompetencije), a drugi u efluksu lekova. ABC transporteri su privukli ekstenzivnu pažnju, zato što oni doprinose otpornosti ćelija na antibiotike i antikancerne agense putem ispumpavanja lekova iz ćelija.[3]
U gram-negativnim organizmima, ABC transporteri posreduju simultanu sekreciju proteinskih supstrata kroz unutrašnje i spoljašnje membranes bez direktnog prolaza kroz periplazmu. Ovaj tip sekrecije se naziva sekrecijom tipa I, koja obuhvata tri komponente koje usklađeno funkcionišu: ABC eksporter, membranski fuzioni protein (MFP), i faktor spoljašnje membrane (OMF). Primer ovakve sekrecije je izlučivanje hemolizina (HlyA) iz E. coli, gde ABC transporter unutrašnje membrane HlyB formira interakcije sa fuzionim proteinom unutrašnje membrane HlyD i faktorom spoljašnje membrane, TolC. TolC omogućava hemolizinu da bude transportovan kroz dve membrane, bez kontakta sa periplazmom.[15]
Bakterijska otpornost na lekove je sve veći zdravstveni problem. Jedan od mehanizama otpornosti na lekove je povezan sa povećanim efluksom antibiotika iz bakterijskih ćelija. Otpornost na lekove usled efluksa posredovanog P-glikoproteinom je originalno zapažena u ćelijama sisara. Kod bakterija, Levi i njegovi saradnici su prvi objavili dokaze da je otpornost na antibiotike uzrokovana efluksom lekova.[61] P-glikoprotein je najbolje izučena efluksna pumpa i kao takva pruža značajan uvid u mehanizam bakterijskih pumpi.[3] Mada pojedini transporteri transportuju specifični tip supstrata, većina transportera iznosi raznovrsnu klase lekova sa znatno različitim hemijskim strukturama.[18] Ti transporteri se zajednički nazivaju ABC transporterima otpornosti na višestruke lekove (MDR), a ponekad se nazivaju i „hidrofobnim usisivačima”.[52]
Ljudski ABCB1/MDR1 P-glikoprotein
[уреди | уреди извор]P-glikoprotein je dobro izučen protein koji doprinosi otpornosti na višestruke lekove. On pripada ljudskoj ABCB (MDR/TAP) familiji. Isto tako je poznat kao ABCB1 ili MDR1 Pgp. MDR1 se sastoji od funkcionalnog monomera sa dva transmembranska domena (TMD) i dva domena vezivanja nukleotida (NBD). Ovaj protein uglavnom može da transportuje katjonske ili električno neutralne supstrate, kao i širok spektar amfifilnih supstrata. Struktura celokupnog ABCB1 monomera je dobijena u prisustvu i odsustvu nukleotida koristeći elektronsku kriokristalografiju. Bez nukleotida, TMD su približno paralelni i formiraju snop koji okružuje centralnu poru, sa otvorom ka ekstracelularnoj strani membrane. U prisustvu nehidrolizibilnog ATP analoga, AMP-PNP, TM domeni su u znatnoj meri reorganizovani sa tri jasno segregirana domena. Centralna pora, koja je opasana TM domenima, je delimično otvorena ka intracelularnoj strani sa razmakom između dva domena koji omogućava pristup supstratu iz lipidne faze. Znatna promena pakovanja i moguća rotacija TM heliksa nakon vezivanja nukleotida sugerišu model heliksne rotacije kao transportni mehanizam.[18]
Biljni ABCB transporteri
[уреди | уреди извор]Genom modelne biljke Arabidopsis thaliana ima sposobnost kodiranja 120 ABC proteina za razliku od 50-70 ABC proteina kodiranih u ljudskom genomu i vinkim mušicama (Drosophila melanogaster). Biljni ABC proteini se kategorišu u 13 potfamilija na osnovu veličine (pun, polu ili četvrt), orijentacije, i sveukupne sličnosti aminokiselinskih sekvenci.[62] Homolozi otpornosti na višestruke lekove (MDR), takođe poznati kao P-glikoproteini, predstavljaju najveću potfamiliju kod biljaka sa 22 člana i drugu sveukupno najveću ABC potfamiliju. B potfamilija biljnih ABC transportera (ABCB) je karakteristična po njihovoj lokaciji u ćelijskoj membrani.[63] Biljni ABCB transporteri su okarakterisani putem heterolognog izražavanja u Escherichia coli, Saccharomyces cerevisiae, Schizosaccharomyces pombe i HeLa ćelija radi određivanja specifičnosti supstrata.
Biljni ABCB transporteri transportuju fitohormon indol-3-sirćetnu kiselinu (IAA),[64] takođe poznatu kao auksin, koja je esencijalni regulator biljnog rasta i razvića.[65][66] Usmereni polarni transport auksina posreduje biljne odgovore na stimuluse iz okruženja putem procesa kao što su fototropizam i gravitropizam.[67] Dva najbolje izučena auksinska transportera, ABCB1 and ABCB19, su okarakterisana kao primarni auksinski eksporteri.[65] Drugi ABCB transporteri kao što je ABCB4 učestvuju u eksportu i importu auksina.[65] Pri niskim intracelularnim koncentracijama auksina ABCB4 unosi auksin dok se ne dostigne određeni nivo, nakon čega se menja smer transporta.[65][68]
Sav1866
[уреди | уреди извор]Prva objavljena struktura visoke rezolucije ABC eksportera je bila za Sav1866 iz Staphylococcus aureus.[18][69] Sav1866 je homolog ABC transportera za višestruke lekove. On ima znatno sličnu sekvencu sa ljudskim ABC transporterima potfamilije B koja obuhvata MDR1 i TAP1/TAP2. Aktivnost ATPaze Sav1866 transportera je stimulisana lekovima za kancer kao što su doksorubicin, vinblastin i drugi,[70] što sugeriše sličnu specifičnost za supstrat sa P-glikoproteinom i stoga mogući zajednički mehanizam substratne translokacije. Sav1866 je homodimer polu transportera, svaka podjedinica sadrži N-terminal TMD sa šest heliksa i C-terminalni NBD. NB domeni imaju slične strukture sa domenima drugih ABC transportera, kod kojih su dva mesta ATP vezivanja formirana na interfejsu dimera između Voker A motiva jednog NBD i LSGGQ motiva drugog. ADP-vezana struktura Sav1866 pokazuje da su NBD u zatvorenom dimeru i da su TM heliksi razdvojeni u dva „krila” orijentisana ka periplazmi, čime formiraju konfiguraciju otvorenu ka spoljašnjoj strani. Svako krilo se sastoji od heliksa TM1-2 iz jedne podjedinice i TM3-6 iz druge podjedinice. Transporter sadrži dugačke intracelularne petlje (ICL ili ICD) koje povezuju TM domene i koje se protežu izvan lipidnog dvosloja u citoplazmu i interaguju sa 8=D. Dok importeri sadrže kratak sprezuči heliks koji je u kontaktu sa jednim NBD, Sav1866 ima dva intracelularna sprežuća heliksa, jedan (ICL1) je u kontaktu sa NB domenima obe podjedinice, a drugi (ICL2) formira interakcije samo sa suprotnom NBD podjedinicom.[20][23][51]
MsbA
[уреди | уреди извор]MsbA je ABC transporter otpornosti na višestruke lekove (MDR) i moguće je da je lipidna flipaza. On je ATPaza koja transportuje lipid A, hidrofobni deo lipopolisaharida (LPS), saharolipid baziran na glukozaminu koji formira spoljašnji monosloj spoljašnjih membrana većine gram-negativnih bakterija. Lipid A je endotoksin i stoga gubitak MsbA iz ćelijske membrane ili mutacije koje poremećuju transport dovode do akumulacije lipida A u unutrašnjoj ćelijskoj membrani, što dovodi do smrti ćelije. On je blisko homologan sa P-glikoproteinom (Pgp) u pogledu proteinskih sekvenci i ima srodnu supstratnu specifičnost sa MDR-ABC transporterom LmrA iz Lactococcus lactis.[71] MsbA iz E. coli je 36% identičan sa NH2-terminalnom polovinom ljudskog MDR1, što sugeriše zajednički mehanizam transporta amfifatičnih i hidrofobnih supstrata. MsbA gen kodira polu transporter koji se sastoji od transmembranskog domena (TMD) stopljenog sa domenim vezivanja nukleotida (NBD). On je konstruisan kao homodimer sa totalnom molekulskom masom od 129,2 kD. MsbA sadrži 6 TM domena na periplazmičnoj strani, jedan NB domen lociran na citoplazmičnoj strani ćelijske membrane, i intracelularni domain (ICD), koji premošćava TMD i NBD. Taj konzervirani heliks koji se proteže od TMD segmenta u ili blizo aktivnog mesta NBD je u znatnoj meri odgovoran za prenos informacija između TMD i NBD. Specifično, ICD1 služi kao konzervirana stacionarna tačka oko koje NBD može da rotira, što omogućava NBD disasocijaciju i dimerizaciju tokom vezivanja ATP i hidrolize.[3][15][18][20][41][51][52][72]

Svojevremeno objavljene (i sad retraktovane) kristalne strukture MsbA su bile inkonzistentne sa bakterijskim homologom Sav1866.[73][74] Strukture su bile preispitane i utvrđeno je da sadrže grešku i da su stoga rezultirajući modeli MsbA inkorektni. Nedavno su greške bile korigovane i nove strukture su objavljene.[38] Odmarajuće stanje E. coli MsbA manifestuje invertovani „V” oblik sa komorom koja je dostupna ka unutrašnjosti transportera. Kontakti dimera su koncentrisani između ekstracelularnih petlji, i dok su NB domeni na rastojanju od ~50Å, podjedinice su jedna naspram druge. Rastojanje između ostataka na mestu dimernog interfejsa je potvrđeno putem eksperimenata unakrsnog vezivanja[75] i EPR spektroskopskim studijama.[76] Relativno velika komora omogućava transport velikih čeonih grupa poput grupa prisutnih u lipidu A. Znatne konformacione promene su neophodne da bi se prenele velike šećerne čeone grupe kroz membranu. Razlika između dve strukture bez nukleotida (apo) je ~30° pomeranje TM4/TM5 heliksa relativno na TM3/TM6 helikse. U zatvorenom apo stanju (iz V. cholerae MsbA), NB domeni su poravnati i mada su bliže, oni ne formiraju ATP sendvič, i P petlje suprotnih monomera su locirane jedna pored druge. U poređenju sa otvorenom konformacijom, dimerni interfejs TM domena u zatvorenoj konformaciji sa licem ka unutra ima ekstenzivne kontakte. Kod obe apo konformacije MsbA, otvor komore je na unutrašnjoj strani. Struktura MsbA-AMP-PNP (5’-adenilil-β-γ-imidodifosfat) iz S. typhimurium je slična sa Sav1866. NB domeni u toj konformaciji sa vezanim nukleotidom i licem ka spoljašnjoj strani, zajedno formiraju kanonički ATP dimerni sendvič, drugim rečima, nukleotid je smešten između P-petlje i LSGGQ motiva. Konformaciona tranzicija iz MsbA-zatvorene-apo do MsbA-AMP-PNP obuhvata dva koraka: ~10° pomeranje TM4/TM5 heliksa ka TM3/TM6, približavanje NB domena mada ne i poravnavanje, čemu sledi zaokretanje TM4/TM5 heliksa za ~20° izvan ravni. Zaokretanje rezultira u separaciji TM3/TM6 heliksa od TM1/TM2, što dovodi do promene iz konformacije iz otvorene ka unutra u konformaciju koja je otvorena ka spoljašnjosti. Stoga, promene u orijentaciji i rastojanju NB domena dramatično rearanžiraju pakovanje transmembranskih heliksa i efektivno menjaju pristup komori iz unutrašnjosti membrane u pristup sa spoljašnje strane.[38] Strukture određene za MsbA su baza transportnog modela putem zaokretanja.[18] Opisane strukture takođe naglašavaju dinamičku prirodu ABC eksportera, što isto tako proizilazi iz fluoroscentnih i EPR studija.[51][76][77]
Mehanizam transporta eksportera
[уреди | уреди извор]
ABC eksporteri imaju transportni mehanizam koji je u skladu sa modelom naizmeničnog pristupa i modelom ATP prekidača. U apo stanjima eksportera, konformacija je orijentisana ka unutrašnjosti, a TM i NB domeni su relativno udaljeni. Kod MsbA, na primer, komora je dovoljno velika da prihvati šećerne grupe iz lipopolisaharida (LPS). Nekoliko grupa naučnika je iznelo pretpostavku da vezivanje supstrata inicira transportni ciklus. ATP vezivanje koje indukuje NBD dimerizaciju i formiranje ATP sendviča, je pogonska sila konformacionih promena u TM domenima. U MsbA proteinu, šećerne čeone grupe su sekvestrirane unutar komore tokom "energetskog stimulusa". Šupljina je obložena naelektrisanim i polarnim ostacima koji su verovatno solvatisani, i čime se formira energetski nepodesno okruženje za hidrofobne supstrate, a energetski podesno za polarne delove amfifilnih jedinjenja ili šećernih grupa iz LPS. Pošto lipid ne može dugo da bude stabilan u okruženju komore, lipid A i drugi hidrofobni molekuli prelaze u energetski povoljniju poziciju unutar spoljašnjeg sloja membrane. Pomeranje takođe može da bude vođeno smicanjem čvrstih tela TM domena dok se hidrofobni repovi LPS molekula provlače kroz lipidni dvosloj. Prepakivanjem heliksa se konformacija menja u stanje sa otvorom ka spoljašnjosti membrane. ATP hidroliza može da proširi periplasmatični otvor i da pogura supstrat ka spoljašnjem sloju lipidnog dvosloja. Hidrolizom drugog ATP molekula i oslobađanjem Pi odvajaju se NB domeni, čemu sledi restauracija odmarajućeg stanja i otvaranje komore ka citoplazmi za sledeći ciklus.[38][41][49][52][73][74][76][78]
Uloga u otpornosti na višestruke lekove
[уреди | уреди извор]Poznato je da ABC transporteri imaju ključnu ulogu u razviću otpornosti na višestruke lekove (MDR). U MDR slučajevima, pacijenti koji koriste lekove konačno razviju otpornost ne samo na lek koji uzimaju, nego i na nekoliko različitih tipova lekova. To je uzrokovano višestrukim faktorima, jedan od kojih je povišeno izlučivanje leka iz ćelija posredstvom ABC transportera. Na primer, ABCB1 protein (P-glikoprotein) iznosi tumorno supresivne lekove iz ćelija. Pgp koji se naziva i MDR1, ABCB1, je prototip ABC transportera, i on je najekstenzivnije izučavan gen te familije. Poznato je da Pgp transportuje organska katjonska ili neutralna jedinjenja. Za nekoliko članova ABCC familije, takođe poznatih kao MRP, je pokazano da imaju MDR dejstvo na organska anjonska jedinjenja. Najbolje izučeni član ABCG familije je ABCG2, takođe poznat kao BCRP (engl. breast cancer resistance protein - protein otpornosti raka dojke), proizvodi otpornost na većinu inhibitora topoizomeraze I ili II, kao što su topotekan, irinotekan, i doksorubicin.
Nije razjašnjeno kako ti proteini mogu da translociraju lekove koji su različiti u tolikoj meri, mada jedan model (model hidrofobnog usisivača) navodi da su u P-glikoproteinu lekovi vezani nezavisno od lipidne faze na bazi njihove hidrofobnosti.
Revertovanje otpornosti na višestruke lekove
[уреди | уреди извор]Otporost na lekove je čest klinički problem koji se javlja kod pacijenata obolelih od infektivnih bolesti i pacijenta obolelih od kancera. Prokariotiski i eukariotski mikroorganizmi kao i neoplastične ćelije su često otporni na lekove. MDR se često objašnjava povišenim izražavanjem ABC transportera. Inhibicija ABC transportera dejstvom jedinjenja niske molekulske težine je bila ekstenzivno izučavana kod obolelih od raka; međutim, klinički rezultati su bili razočaravajući. Nedavno su razne RNKi strategije primenjene u revertovanju MDR u različitim modelima tumora i utvrđeno je da je ta tehnologija efektivna u poništavanju MDR posredovanog ABC-transporterima u ćelijama raka, te se stoga smatra obećavajućom strategijom za prevazilaženje MDR upotrebom genskih terapeutika. Primena RNKi tehnologije takođe ima potencijal da nađe primenu u prevazilaženju MDR tretmana infektivnih bolesti uzrokovanih mikrobnim patogenima.[79][80]
Fiziološka uloga
[уреди | уреди извор]Pored uzrokovanja MDR kod tumorskih ćelija, ABC transporteri su takođe izraženi u membranama zdravih ćelija, gde posreduju transport raznih endogenih supstanci, kao i niza supstanci koje su strane telu. Na primer, ABC transporteri kao što je Pgp, MRP i BCRP ograničavaju apsorpciju mnogih lekova iz intestinalnog trakta, i pumpaju lekove iz ćelija jetre u žuč, kao način uklanjanja stranih materija iz tela. Veliki broj lekova je bilo transportovan samim ABC transporterima, ili utiče na transport drugih lekova putem tih proteina. Ovaj drugi scenario može da dovede do interakcija između lekova, te da uzrokuje promene u dejstvu lekova.[81]
Metodi za karakterisanja interakcija ABC transportera
[уреди | уреди извор]Postoji nekoliko tipova testova koji omogućavaju detekciju interakcija ABC transportera sa endogenim i ksenobiotičkim jedinjenjima.[82] Kompleksnost testova je u opsegu od relativno jednostavnih membranskih,[83] poput testa vezikularnog transporta, ATPaznog testa, preko kompleksnijih ćelijskih testova i do složenih in vivo[84] detekcionih metodologija.[85]
Membranski testovi
[уреди | уреди извор]Testom vezikularnog transporta se detektuje translokacija molekula posredstvom ABC transportera.[86] Membrane pripremljene pod podesnim uslovima sadrže obrnuto orijentisane vezikule sa mestima vezivanja ATP i mestima vezivanja supstrata transportera na spoljašnjoj strani. Vezikule preuzimaju substrate transportera na način zavistan od ATP. Brza filtracija koristeći filtere sa staklenim vlaknima ili nitroceluloznim membranama se koristi za separaciju vezikula od inkubacionog rastvora, pri čemu se testirano jedinjenje zarobljeno unutar vezikula zadržava na filteru. Količina transportovanih neobeleženih molekula se određuje putem HPLC, LC/MS, LC/MS/MS. Alternativno, jedinjenja se obeležavaju radioaktivno ili sa fluorescentnim oznakama, tako da se radioaktivnost ili fluorescencija koji su zadržavani na filteru mogu kvantifikovati.
Razni tipovi membrana iz različitih izvora (e.g. ćelije insekta, transfektovane ili odabrane ćelijske linije sisara) se koriste u vezikularnim transportnim studijama. Membrane su komercijalno dostupne[87] ili se mogu pripremiti iz raznih ćelija ili čak tkiva, e.g. kanalikularne membrane jetre. Ovaj tip testa ima prednost u pogledu merenja stvarne dispozicije supstrata kroz ćelijsku membranu. Njegov nedostatak je da se jedinjenja sa srednjom do visoke pasivne permeabilnosti ne zadržavaju unutar vezikula, što čini direktna merenja transporta teško izvodivim.
Test vezikularnog transporta može da bude izveden na indirektan način, tako što interagujući testirani lekovi modulišu brzinu transporta reporterskog jedinjenja. Takav tip testa je posebno podesan za detekciju mogućih interakcija između lekova i interakcija lekova sa endogenim supstratima. Ovi testovi nisu senzitivni na pasivnu permeabilnost jedinjenja i stoga detektuju sva interagujuća jedinjenja. Ovaj tip testa ne pruža informacije o tome da li je testirano jedinjenje inhibitor transportera, ili supstrat transportera koji inhibira njegovu funkciju na kompetitivan način. Tipičan primer indirektnog vezikularnog transportnog testa je detekcija inhibicije tauroholatnog transporta posredstvom ABCB11 (BSEP).
Testovi bazirani na celim ćelijama
[уреди | уреди извор]Ćelije koje izražavaju efluksne transportere aktivno pumpaju supstrate iz ćelija, što dovodi do snižavenja brzine akumulacije supstrata, nižih intracelularnih koncentracija tokom stacionarnog stanja, ili veće brzine eliminacije supstrata iz ćelija zasićenih supstratom. Transportovani radioaktivni supstrati ili obležavajće fluorescentne boje se mogu direktno meriti, ili na indirektan način se modulacija akumulacije probnog supstrata (e.g. fluorescentne boje, kao što je Rho123, ili kalcein) može odrediti u prisustvu testiranog leka.
Kalcein-AM je visoko permeabilni derivat kalceina, te lako penetrira neoštećene ćelije, gde ga endogene esteraze brzo hidrolizuju do fluorescentnog kalceina. U kontrastu sa kalceinom AM, kalcein ima nisku permeabilnosti i stoga je zarobljen u ćelijama, gde se akumulira. Pošto je kalcein-AM ekselentan supstrat MDR1 i MRP1 efluks transportera, ćelije koje izražavaju MDR1 i/ili MRP1 transportere ispumpavaju kalcein-AM iz ćelije pre nego što ga esteraze mogu hidrolizovati. Rezultat toga je niža ćelijska brzina akumulacije kalceina. Što je veća MDR aktivnost u ćelijskoj membrani, to se manje kalceina akumulira u citoplazmi. U MDR izražavajućim ćelijama, dodatak MDR inhibitora ili MDR supstrata u višku dramatično povišava brzinu akumulacije kalceina. Aktivnost transportera višestrukih lekova se odražava kao razlika između količina boje akumulirane u prisustvu i odsustvu inhibitora. Koristeći selektivne inhibitore, može se uočiti razlika između transportne aktivnosti MDR1 i MRP1. Ovaj test se može koristiti za testiranje lekova za transporterske interakcije, kao i za kvantifikaciju MDR aktivnosti ćelija. Kalceinski test je u vlasništvu preduzeća SOLVO Biotechnology.
Potfamilije
[уреди | уреди извор]Ljudske potfamilije
[уреди | уреди извор]Poznato je 48 ABC transportera koji su prisutni kod ljudi. Ljudska genomska organizacija ih je klasifikovala u sedam familija.[88]
| Familija | Članovi | Funkcije | Primeri |
|---|---|---|---|
| ABCA | Ova familija sadrži neke od najvećih transportera (preko 2.100 aminokiselina dugačkih). Pet njih je locirano u klasteru na 17q24 hromozomu. | Odgovorni su za transport holesterola i lipida, između ostalog. | ABCA12 ABCA1 |
| ABCB | Sastoji se od 4 puna i 7 polutransportera. | Neki od njih su locirani u krvno–moždanoj barijeri, jetri, mitohondrijama. Oni na primer transportuju peptide i žuč. | ABCB5 |
| ABCC | Sastoji se od 12 punih transportera. | Koriste se u jonskom transportu, receptorima ćelijske površine, sekreciji toksina. Ova grupa obuhvata CFTR protein, koji uzrokuje cističnu fibrozu kad je u deficitu. | ABCC6 |
| ABCD | Sastoji se od 4 polu transportera | Koriste se u peroksizomima. | ABCD1 |
| ABCE/ABCF | Sastoji se od 1 ABCE i 3 ABCF proteina. | Oni zapravo nisu transporteri, nego samo ATP-vezujući domeni koji su izvedeni iz ABC familije, mada bez transmembranskih domena. Ovi proteini uglavnom regulišu proteinsku sintezu ili izražavanje. | ABCE, ABCF1, ABCF2 |
| ABCG | Sastoji se od 6 „reverznih” polutransportera, sa NBF na NH3+ kraju i TM na COO- kraju. | Transportuju lipide, raznovrsne supstrate lekova, žuč, holesterol, i druge steroide. | ABCG2 ABCG1 |
ABCA
[уреди | уреди извор]ABCA potfamilija se sastoji od 12 punih transportera podeljenih u dve podgrupe. Prva podgrupa se sastoji od sedam gena koji su mapirani na šest različitih hromozoma. Oni su ABCA1-4, A7, A12, i A13. Druga podgrupa se sastoji od ABCA5-6, i A8-10. Celokupna podgrupa 2 je organizovana kao jedan klaster gena na hromozomu 17q24. Geni druge podgrupe se razlikuju od gena sličnih ABCA1 po tome što imaju 37-38 eksona za razliku od 50 eksona u ABCA1.
ABCA1 podgrupa je implicirana u razviću genetičkih bolesti. U recesivnoj Tangierovoj bolesti, ABCA1 protein je mutiran. Takođe, ABCA4 je u regionu hromozoma 1p21 koji sadrži gen za Stargardtovu bolest. Utvrđeno je da je taj gen visoko izražen u štapićastim fotoreceptorima i da je motiran kod obolelih od Stargardtove bolesti, recesivnog retinitisnog pigmentizma, i većine recesivnih kupasto-štapićastih distrofija.[9]
ABCB
[уреди | уреди извор]ABCB potfamilija se sastoji od četiri puna transportera i dva polutransportera. Ona je jedina ljudska potfamilija koja sadrži pune i polu tipove transporters. ABCB1 (glikoprotein permeabilnosti, P-gp, Pgp) je otkriven kao protein koji je prekomerno izražen u pojedinim ćelijama tumora otpornim na lekove. Ovaj glikoprotein je kod ljudi kodiran ABCB1 genom.[89][90] On je prvenstveno izražen u krvno-moždanoj barijeri i jetri.[91] Smatra se da učestvuje u zaštiti ćelija od toksina. Ćelije koje prekomerno izražavaju ovaj protein manifestuju otpornost na višestruke lekove.[9][92][93][94]
ABCC
[уреди | уреди извор]Podfamlija ABCC sadrži trinaest članova. Devet transportera ove grupe se naziva proteinima otpornosti na višestruke lekove (MRP). MRP proteini su široko zastupljeni u prirodi i posreduju mnoštvo važnih funkcija.[95] Poznato je da oni učestvuju u jonskom transportu, sekreciji toksina, i prenosu signala.[9] Od devet MRP proteina njih četiri, MRP4, 5, 8, 9, (ABCC4, 5, 11, i 12), imaju tipične ABC strukture sa četiri domena, što obuhvata dva domena koji premošćavaju membranu, pri čemu svakom od njih sledi domen vezivanja nukleotida. Oni se skraćeno nazivaju MRP. Preostalih 5 MRP proteina, MRP1, 2, 6, 7 (ABCC1, 2, 3, 6 i 10) su poznati kao dugački MRP proteini. Oni sadrže dodatni peti domen na njihovom N-terminusu.[95]
CFTR, transporter koji učestvuje u bolesti cistična fibroza,[96][97] se takođe smatra delom ove familije. Cistična fibroza se javlja nakon mutacije i gubitka CFTR funkcije.[9] CFTR transportuje hloridne[98] i tiocijanatne[99] jone kroz epitelijalne ćelijske membrane. Mutacije CFTR gena utiču na funkcionisanje kanala hloridnih jona u tim ćelijskim membranama, što dovodi do cistične fibroze i kongenitalnog odsustva vas deferensa.[100]
Sulfonilurejni receptori (SUR), koji učestvuju u sekreciji insulina, i u neuronskim i mišićnim funkcijama, su takođe deo ove familije proteina.[101][102] Mutacije SUR proteina su potencijalni uzrok neonatalnog dijabetes melitusa.[103] SUR je isto tako mesto vezivanja lekova poput sulfonilureja i aktovatora otvaranja kalijumskih kanala kao što je diazoksid.
ABCD
[уреди | уреди извор]ABCD potfamilija se sastoji od četiri gena koji kodiraju polutransportere ekskluzivno izražene u peroksizomu. ABCD1 je odgovoran za X-vezanu formu adrenoleukodistrofije (ALD).[104] To je bolest karakterisana neurodegeneracijom i adrenalnom deficijencijom koja se tipično inicira u kasnom detinjstvu.[105] U ćelijama ALD pacijenata se akumuliraju nerazgranate zasićene masne kiseline, mada precizna uloga ABCD1 u tom procesu još uvek nije utvrđena. Funkcija drugih ABCD gena za sad nije poznata. Postoje nagoveštaji da oni imaju srodne funkcije u metabolizmu masnih kiselina.[9]
ABCE i ABCF
[уреди | уреди извор]Ove dve podgrupe se sastoje od gena koji imaju domene ATP vezivanja koji su blisko srodni sa drugim ABC transporterima, mada ovi geni ne kodiraju transmembranske domene. ABCE se ima samo jednog člana, OABP ili ABCE1,[106] za koji je poznato da prepoznaje pojedine oligodendrocite proizvedene u responsu na specifične virusne infekcije. Svi članovi ABCF podgrupe se sastoje od para domena vezivanja ATP.[9][107][108]
ABCG
[уреди | уреди извор]Šest polutransportera sa mestima ATP vezivanja na N terminusu i transmembranskim domenima na C terminusu sačinjavaju ABCG potfamiliju. Takva orijentacija je suprotna svim drugim ABC genima. Postoji samo 5 ABCG gena u ljudskom genomu,[108][109][110] dok Drosophelia genom sadrži 15, a kvasac 10 gena.
ABCG2 gen je otkriven u ćelijskim linijama koje su izdvojene zbog njihovog visokog nivoa otpornosti na mitoksantron, i zbog odsustva izražavanja ABCB1 ili ABCC1. ABCG2 može da eksportuje antrociklinske antikancerne lekove, kao i topotekan, mitoksantron, ili doksorubicin. Utvrđeno je da hromozomske translokacije uzrokuju amplifikaciju ili preuređenje ABCG2 gena prisutnog u rezistentnim ćelijskim linijama. Normalna funkcija ABCG2 proteina nije poznata.[9]
Prokariotske potfamilije
[уреди | уреди извор]Sledeći klasifikacioni sistem za transmembranske transportere rastvorka je razvijen:[111]
Importeri
[уреди | уреди извор]Permeaze preuzimanja ABC-tipa
- 3.A.1.1 Transporter-1 ugljenohidratnog preuzimanja (CUT1)
- 3.A.1.2 Transporter-2 ugljenohidratnog preuzimanja (CUT2)
- 3.A.1.3 Transporter preuzimanja polarnih aminokiselina (PAAT)
- 3.A.1.4 Transporter preuzimanja hidrofobnih aminokiselina (HAAT)
- 3.A.1.5 Transporter preuzimanja peptida/opina/nikela (PepT)
- 3.A.1.6 Transporter preuzimanja sulfata/volframa (SulT)
- 3.A.1.7 Transporter preuzimanja fosfata (PhoT)
- 3.A.1.8 Transporter preuzimanja molibdata (MolT)
- 3.A.1.9 Transporter preuzimanja fosfonata (PhnT)
- 3.A.1.10 Transporter preuzimanja feri gvožđa (FeT)
- 3.A.1.11 Transporter preuzimanja poliamin/opin/fosfonata (POPT)
- 3.A.1.12 Transporter preuzimanja kvaternarnih amina (QAT)
- 3.A.1.13 Transporter preuzimanja vitamina B12 (B12T)
- 3.A.1.14 Transporter preuzimanja helata gvožđa (FeCT)
- 3.A.1.15 Transporter preuzimanja helata mangana/cinka/gvožđa (MZT)
- 3.A.1.16 Transporter preuzimanja nitrata/nitrita/cijanata (NitT)
- 3.A.1.17 Transporter preuzimanja taurina (TauT)
- 3.A.1.18 Transporter preuzimanja kobalta (CoT)
- 3.A.1.19 Transporter preuzimanja tiamina (ThiT)
- 3.A.1.20 Transporter brahispirnog gvožđa (BIT)
- Transporter preuzimanja siderofora-Fe3+ (SIUT)
- Transporter preuzimanja nikla (NiT)
- Transporter preuzimanja metionina (MUT)
- 2.A.52 Transporter preuzimanja nikla/kobalta (NiCoT)
- 3.A.1.106 Lipidni eksporter (LipidE)
Eksporteri
[уреди | уреди извор]Efluksne permeaze ABC-tipa (prokariotske)
- 3.A.1.101 Kapsularna polisaharidna eksporterska (CPSE) familija
- 3.A.1.102 Lipooligosaharidna eksporterska (LOSE) familija
- 3.A.1.103 Lipopolisaharidna eksporterska (LPSE) familija
- 3.A.1.104 Teihoinsko kiselinska eksporterska (TAE) familija
- 3.A.1.105 Familija eksportera lekova (DrugE1)
- 3.A.1.106 Familija eksportera putativnog lipida A (LipidE)
- 3.A.1.107 Familija eksportera putativnog hema (HemeE)
- 3.A.1.108 Familija eksportera β-glukana (GlucanE)
- 3.A.1.109 Familija eksportera proteina-1 (Prot1E)
- 3.A.1.110 Familija eksportera proteina-2 (Prot2E)
- 3.A.1.111 Familija eksportera peptida-1 (Pep1E)
- 3.A.1.112 Familija eksportera peptida-2 (Pep2E)
- 3.A.1.113 Familija eksportera peptida-3 (Pep3E)
- 3.A.1.114 Familija eksportera glikolipida (DevE)
- 3.A.1.115 Familija eksportera Na+ (NatE)
- 3.A.1.116 Familija eksportera mikrocina B17 (McbE)
- 3.A.1.117 Familija eksportera 2 lekova (DrugE2)
- 3.A.1.118 Familija eksportera mikrocina J25 (McjD)
- 3.A.1.119 Familija eksportera 3 lekova/siderofora (DrugE3)
- (Putativna) ATPaza-1 otpornositi na lekove (Drug RA1)
- (Putativna) ATPaza-2 otpornositi na lekove (Drug RA2)
- Makrolidni eksporter (MacB)
- Peptidni-4 eksporter (Pep4E)
- 3-komponentni peptidni-5 eksporter (Pep5E)
- Lipoproteinska translokaza (LPT)
- β-Eksotoksinski I eksporter (βETE)
- AmfS peptidni eksporter (AmfS-E)
- SkfA peptidni eksporter (SkfA-E)
- CydDC cisteinski i glutationski eksporter (CydDC-E)
Kristalne strukture
[уреди | уреди извор]Mnoštvo struktura domena ABC protena koji su rastvorni u vodi je proizvedeno zadnjih godina.[1][112] Oni imaju značajnu ulogu u izučavanju mehanizma dejstva manje poznatih članova ove proteinske familije.
Vidi još
[уреди | уреди извор]Reference
[уреди | уреди извор]- ^ а б Jones PM, George AM (2004). „The ABC transporter structure and mechanism: perspectives on recent research”. Cell Mol Life Sci. 61 (6): 682—99. PMID 15052411. doi:10.1007/s00018-003-3336-9.
- ^ Ponte-Sucre, A, ur. (2009). ABC Transporters in Microorganisms. Caister Academic Press. ISBN 978-1-904455-49-3.
- ^ а б в г д ђ е ж з и ј к л љ м Davidson AL, Dassa E, Orelle C, Chen J (2008). „Structure, function, and evolution of bacterial ATP-binding cassette systems”. Microbiol. Mol. Biol. Rev. 72 (2): 317—64,table of contents. PMC 2415747
. PMID 18535149. doi:10.1128/MMBR.00031-07.
- ^ а б в г Goffeau, A.; B. de Hertogh; and P.V. Baret. 2004. ABC Transporters. In: Encyclopedia of Biological Chemistry. Vol. 1, 1–5.
- ^ Wang B, Dukarevich M, Sun EI, Yen MR, Saier MH (2009). „Membrane porters of ATP-binding cassette transport systems are polyphyletic”. The Journal of Membrane Biology. 231 (1): 1—10. PMID 19806386. doi:10.1007/s00232-009-9200-6.
- ^ Khwaja M, Ma Q, Saier MH (2005). „Topological analysis of integral membrane constituents of prokaryotic ABC efflux systems”. Research in Microbiology. 156 (2): 270—7. PMID 15748994. doi:10.1016/j.resmic.2004.07.010.
- ^ ter Beek J, Guskov A, Slotboom DJ (2014). „Structural diversity of ABC transporters”. The Journal of General Physiology. 143 (4): 419—35. PMC 3971661
- ^ а б Choi CH (2005). „ABC transporters as multidrug resistance mechanisms and the development of chemosensitizers for their reversal”. Cancer Cell International. 5: 30. PMC 1277830
- ^ а б в г д ђ е ж Dean M, Hamon Y, Chimini G (2001). „The human ATP-binding cassette (ABC) transporter superfamily”. Journal of Lipid Research. 42 (7): 1007—17. PMID 11441126.
- ^ Scott MP, Lodish HF, Berk A, Kaiser, C, Krieger M, Bretscher A, Ploegh H, Amon A (2012). Molecular Cell Biology. San Francisco: W. H. Freeman. ISBN 978-1-4292-3413-9.
- ^ Henderson DP, Payne SM (1994). „Vibrio cholerae iron transport systems: roles of heme and siderophore iron transport in virulence and identification of a gene associated with multiple iron transport systems”. Infect. Immun. 62 (11): 5120—5. PMC 303233
- ^ Cangelosi GA, Ankenbauer RG, Nester EW (1990). „Sugars induce the Agrobacterium virulence genes through a periplasmic binding protein and a transmembrane signal protein”. Proc. Natl. Acad. Sci. U.S.A. 87 (17): 6708—12. PMC 54606
- ^ Kemner JM, Liang X, Nester EW (1997). „The Agrobacterium tumefaciens virulence gene chvE is part of a putative ABC-type sugar transport operon”. J. Bacteriol. 179 (7): 2452—8. PMC 178989
- ^ Poolman B, Spitzer JJ, Wood JM (2004). „Bacterial osmosensing: roles of membrane structure and electrostatics in lipid-protein and protein-protein interactions”. Biochim. Biophys. Acta. 1666 (1-2): 88—104. PMID 15519310. doi:10.1016/j.bbamem.2004.06.013.
- ^ а б в г д ђ Davidson AL, Chen J (2004). „ATP-binding cassette transporters in bacteria”. Annu. Rev. Biochem. 73: 241—68. PMID 15189142. doi:10.1146/annurev.biochem.73.011303.073626.
- ^ Zhou Z, White KA, Polissi A, Georgopoulos C, Raetz CR (1998). „Function of Escherichia coli MsbA, an essential ABC family transporter, in lipid A and phospholipid biosynthesis”. J. Biol. Chem. 273 (20): 12466—75. PMID 9575204. doi:10.1074/jbc.273.20.12466.
- ^ Poole RK, Gibson F, Wu G (1994). „The cydD gene product, component of a heterodimeric ABC transporter, is required for assembly of periplasmic cytochrome c and of cytochrome bd in Escherichia coli”. FEMS Microbiol. Lett. 117 (2): 217—23. PMID 8181727. doi:10.1111/j.1574-6968.1994.tb06768.x.
- ^ а б в г д ђ е ж Pohl A, Devaux PF, Herrmann A (2005). „Function of prokaryotic and eukaryotic ABC proteins in lipid transport”. Biochim. Biophys. Acta. 1733 (1): 29—52. PMID 15749056. doi:10.1016/j.bbalip.2004.12.007.
- ^ Gedeon C, Behravan J, Koren G, Piquette-Miller M (2006). „Transport of glyburide by placental ABC transporters: implications in fetal drug exposure”. Placenta. 27 (11–12): 1096—102. PMID 16460798. doi:10.1016/j.placenta.2005.11.012.
- ^ а б в г д ђ е ж з и ј к л љ м н Rees DC, Johnson E, Lewinson O (2009). „ABC transporters: the power to change”. Nat. Rev. Mol. Cell Biol. 10 (3): 218—27. PMC 2830722
- ^ а б в Locher KP, Lee AT, Rees DC (2002). „The E. coli BtuCD structure: a framework for ABC transporter architecture and mechanism”. Science. 296 (5570): 1091—8. PMID 12004122. doi:10.1126/science.1071142.
- ^ Hvorup RN, Goetz BA, Niederer M, Hollenstein K, Perozo E, Locher KP (2007). „Asymmetry in the structure of the ABC transporter-binding protein complex BtuCD-BtuF”. Science. 317 (5843): 1387—90. PMID 17673622. doi:10.1126/science.1145950.
- ^ а б в Dawson RJ, Locher KP (2006). „Structure of a bacterial multidrug ABC transporter”. Nature. 443 (7108): 180—5. PMID 16943773. doi:10.1038/nature05155.
- ^ а б в Hollenstein K, Frei DC, Locher KP (2007). „Structure of an ABC transporter in complex with its binding protein”. Nature. 446 (7132): 213—6. PMID 17322901. doi:10.1038/nature05626.
- ^ а б Oldham ML, Khare D, Quiocho FA, Davidson AL, Chen J (2007). „Crystal structure of a catalytic intermediate of the maltose transporter”. Nature. 450 (7169): 515—21. PMID 18033289. doi:10.1038/nature06264.
- ^ Kadaba NS, Kaiser JT, Johnson E, Lee A, Rees DC (2008). „The high-affinity E. coli methionine ABC transporter: structure and allosteric regulation”. Science. 321 (5886): 250—3. PMC 2527972
- ^ а б в г Pinkett HW, Lee AT, Lum P, Locher KP, Rees DC (2007). „An inward-facing conformation of a putative metal-chelate-type ABC transporter”. Science. 315 (5810): 373—7. PMID 17158291. doi:10.1126/science.1133488.
- ^ а б Moody JE, Millen L, Binns D, Hunt JF, Thomas PJ (2002). „Cooperative, ATP-dependent association of the nucleotide binding cassettes during the catalytic cycle of ATP-binding cassette transporters”. J. Biol. Chem. 277 (24): 21111—4. PMC 3516282
- ^ Hung LW, Wang IX, Nikaido K, Liu PQ, Ames GF, Kim SH (1998). „Crystal structure of the ATP-binding subunit of an ABC transporter”. Nature. 396 (6712): 703—7. PMID 9872322. doi:10.1038/25393.
- ^ а б в Verdon G, Albers SV, Dijkstra BW, Driessen AJ, Thunnissen AM (2003). „Crystal structures of the ATPase subunit of the glucose ABC transporter from Sulfolobus solfataricus: nucleotide-free and nucleotide-bound conformations”. J. Mol. Biol. 330 (2): 343—58. PMID 12823973. doi:10.1016/S0022-2836(03)00575-8.
- ^ а б Karpowich N, Martsinkevich O, Millen L, Yuan YR, Dai PL, MacVey K, Thomas PJ, Hunt JF (2001). „Crystal structures of the MJ1267 ATP binding cassette reveal an induced-fit effect at the ATPase active site of an ABC transporter”. Structure. 9 (7): 571—86. PMID 11470432. doi:10.1016/S0969-2126(01)00617-7.
- ^ а б в г Chen J, Lu G, Lin J, Davidson AL, Quiocho FA (2003). „A tweezers-like motion of the ATP-binding cassette dimer in an ABC transport cycle”. Mol. Cell. 12 (3): 651—61. PMID 14527411. doi:10.1016/j.molcel.2003.08.004.
- ^ а б в Diederichs K, Diez J, Greller G, Müller C, Breed J, Schnell C, Vonrhein C, Boos W, Welte W (2000). „Crystal structure of MalK, the ATPase subunit of the trehalose/maltose ABC transporter of the archaeon Thermococcus litoralis”. EMBO J. 19 (22): 5951—61. PMC 305842
- ^ а б Gaudet R, Wiley DC (2001). „Structure of the ABC ATPase domain of human TAP1, the transporter associated with antigen processing”. EMBO J. 20 (17): 4964—72. PMC 125601
- ^ Schmitt L, Benabdelhak H, Blight MA, Holland IB, Stubbs MT (2003). „Crystal structure of the nucleotide-binding domain of the ABC-transporter haemolysin B: identification of a variable region within ABC helical domains”. J. Mol. Biol. 330 (2): 333—42. PMID 12823972. doi:10.1016/S0022-2836(03)00592-8.
- ^ а б Yuan YR, Blecker S, Martsinkevich O, Millen L, Thomas PJ, Hunt JF (2001). „The crystal structure of the MJ0796 ATP-binding cassette. Implications for the structural consequences of ATP hydrolysis in the active site of an ABC transporter”. J. Biol. Chem. 276 (34): 32313—21. PMID 11402022. doi:10.1074/jbc.M100758200.
- ^ а б в г д ђ Smith PC, Karpowich N, Millen L, Moody JE, Rosen J, Thomas PJ, Hunt JF (2002). „ATP binding to the motor domain from an ABC transporter drives formation of a nucleotide sandwich dimer”. Mol. Cell. 10 (1): 139—49. PMC 3516284
- ^ а б в г д Ward A, Reyes CL, Yu J, Roth CB, Chang G (2007). „Flexibility in the ABC transporter MsbA: Alternating access with a twist”. Proc. Natl. Acad. Sci. U.S.A. 104 (48): 19005—10. PMC 2141898
- ^ а б Hopfner KP, Karcher A, Shin DS, Craig L, Arthur LM, Carney JP, Tainer JA (2000). „Structural biology of Rad50 ATPase: ATP-driven conformational control in DNA double-strand break repair and the ABC-ATPase superfamily”. Cell. 101 (7): 789—800. PMID 10892749. doi:10.1016/S0092-8674(00)80890-9.
- ^ Fetsch EE, Davidson AL (2002). „Vanadate-catalyzed photocleavage of the signature motif of an ATP-binding cassette (ABC) transporter”. Proc. Natl. Acad. Sci. U.S.A. 99 (15): 9685—90. PMC 124977
- ^ а б в г Reyes CL, Ward A, Yu J, Chang G (2006). „The structures of MsbA: Insight into ABC transporter-mediated multidrug efflux”. FEBS Lett. 580 (4): 1042—8. PMID 16337944. doi:10.1016/j.febslet.2005.11.033.
- ^ Ambudkar SV, Kim IW, Xia D, Sauna ZE (2006). „The A-loop, a novel conserved aromatic acid subdomain upstream of the Walker A motif in ABC transporters, is critical for ATP binding”. FEBS Lett. 580 (4): 1049—55. PMID 16412422. doi:10.1016/j.febslet.2005.12.051.
- ^ а б Geourjon C, Orelle C, Steinfels E, Blanchet C, Deléage G, Di Pietro A, Jault JM (2001). „A common mechanism for ATP hydrolysis in ABC transporter and helicase superfamilies”. Trends Biochem. Sci. 26 (9): 539—44. PMID 11551790. doi:10.1016/S0968-0004(01)01907-7.
- ^ Ye J, Osborne AR, Groll M, Rapoport TA (2004). „RecA-like motor ATPases--lessons from structures”. Biochim. Biophys. Acta. 1659 (1): 1—18. PMID 15511523. doi:10.1016/j.bbabio.2004.06.003.
- ^ а б Zaitseva J, Jenewein S, Jumpertz T, Holland IB, Schmitt L (2005). „H662 is the linchpin of ATP hydrolysis in the nucleotide-binding domain of the ABC transporter HlyB”. EMBO J. 24 (11): 1901—10. PMC 1142601
- ^ Maegley KA, Admiraal SJ, Herschlag D (1996). „Ras-catalyzed hydrolysis of GTP: a new perspective from model studies”. Proc. Natl. Acad. Sci. U.S.A. 93 (16): 8160—6. PMC 38640
- ^ Matte A, Tari LW, Delbaere LT (1998). „How do kinases transfer phosphoryl groups?”. Structure. 6 (4): 413—9. PMID 9562560. doi:10.1016/S0969-2126(98)00043-4.
- ^ а б Hollenstein K, Dawson RJ, Locher KP (2007). „Structure and mechanism of ABC transporter proteins”. Curr. Opin. Struct. Biol. 17 (4): 412—8. PMID 17723295. doi:10.1016/j.sbi.2007.07.003.
- ^ а б в г д ђ е Higgins CF, Linton KJ (2004). „The ATP switch model for ABC transporters”. Nat. Struct. Mol. Biol. 11 (10): 918—26. PMID 15452563. doi:10.1038/nsmb836.
- ^ Locher, K. P. (2004). „Structure and mechanism of ABC transporters”. Curr. Opin. Struct. Biol. 14 (4): 426—31. PMID 15313236. doi:10.1016/j.sbi.2004.06.005.
- ^ а б в г д ђ е ж Oldham ML, Davidson AL, Chen J (2008). „Structural insights into ABC transporter mechanism”. Curr. Opin. Struct. Biol. 18 (6): 726—33. PMC 2643341
- ^ а б в г Chang, G. (2003). „Multidrug resistance ABC transporters”. FEBS Lett. 555 (1): 102—5. PMID 14630327. doi:10.1016/S0014-5793(03)01085-8.
- ^ Ling, V. (1997). „Multidrug resistance: molecular mechanisms and clinical relevance”. Cancer Chemother. Pharmacol. 40 Suppl (7): S3—8. PMID 9272126. doi:10.1007/s002800051053.
- ^ Senior AE, al-Shawi MK, Urbatsch IL (1995). „The catalytic cycle of P-glycoprotein”. FEBS Lett. 377 (3): 285—9. PMID 8549739. doi:10.1016/0014-5793(95)01345-8.
- ^ Martin C, Higgins CF, Callaghan R (2001). „The vinblastine binding site adopts high- and low-affinity conformations during a transport cycle of P-glycoprotein”. Biochemistry. 40 (51): 15733—42. PMID 11747450. doi:10.1021/bi011211z.
- ^ Manciu L, Chang XB, Buyse F, Hou YX, Gustot A, Riordan JR, Ruysschaert JM (2003). „Intermediate structural states involved in MRP1-mediated drug transport. Role of glutathione”. J. Biol. Chem. 278 (5): 3347—56. PMID 12424247. doi:10.1074/jbc.M207963200.
- ^ Kreimer DI, Chai KP, Ferro-Luzzi Ames G (2000). „Nonequivalence of the nucleotide-binding subunits of an ABC transporter, the histidine permease, and conformational changes in the membrane complex”. Biochemistry. 39 (46): 14183—95. PMID 11087367. doi:10.1021/bi001066.
- ^ Vigano C, Margolles A, van Veen HW, Konings WN, Ruysschaert JM (2000). „Secondary and tertiary structure changes of reconstituted LmrA induced by nucleotide binding or hydrolysis. A fourier transform attenuated total reflection infrared spectroscopy and tryptophan fluorescence quenching analysis”. J. Biol. Chem. 275 (15): 10962—7. PMID 10753896. doi:10.1074/jbc.275.15.10962.
- ^ Sonveaux N, Vigano C, Shapiro AB, Ling V, Ruysschaert JM (1999). „Ligand-mediated tertiary structure changes of reconstituted P-glycoprotein. A tryptophan fluorescence quenching analysis”. J. Biol. Chem. 274 (25): 17649—54. PMID 10364203. doi:10.1074/jbc.274.25.17649.
- ^ Rosenberg MF, Velarde G, Ford RC, Martin C, Berridge G, Kerr ID, Callaghan R, Schmidlin A, Wooding C, Linton KJ, Higgins CF (2001). „Repacking of the transmembrane domains of P-glycoprotein during the transport ATPase cycle”. EMBO J. 20 (20): 5615—25. PMC 125677
- ^ McMurry L, Petrucci RE, Levy SB (1980). „Active efflux of tetracycline encoded by four genetically different tetracycline resistance determinants in Escherichia coli”. Proc. Natl. Acad. Sci. U.S.A. 77 (7): 3974—7. PMC 349750
- ^ Rea, P. A. (2007). „Plant ATP-binding cassette transporters”. Annu Rev Plant Biol. 58: 347—75. PMID 17263663. doi:10.1146/annurev.arplant.57.032905.105406.
- ^ Bailly A, Yang H, Martinoia E, Geisler M, Murphy AS (2011). „Plant Lessons: Exploring ABCB Functionality Through Structural Modeling”. Front Plant Sci. 2: 108. PMC 3355715
- ^ Geisler M, Murphy AS (2006). „The ABC of auxin transport: the role of p-glycoproteins in plant development”. FEBS Lett. 580 (4): 1094—102. PMID 16359667. doi:10.1016/j.febslet.2005.11.054.
- ^ а б в г Yang H, Murphy AS (2009). „Functional expression and characterization of Arabidopsis ABCB, AUX 1 and PIN auxin transporters in Schizosaccharomyces pombe”. Plant J. 59 (1): 179—91. PMID 19309458. doi:10.1111/j.1365-313X.2009.03856.x.
- ^ Blakeslee JJ, Peer WA, Murphy AS (2005). „Auxin transport”. Curr. Opin. Plant Biol. 8 (5): 494—500. PMID 16054428. doi:10.1016/j.pbi.2005.07.014.
- ^ Kretzschmar T, Burla B, Lee Y, Martinoia E, Nagy R (2011). „Functions of ABC transporters in plants”. Essays Biochem. 50 (1): 145—60. PMID 21967056. doi:10.1042/bse0500145.
- ^ Kubeš, Martin; Yang, Haibing; Richter, Gregory L.; Cheng, Yan; Młodzińska, Ewa; Wang, Xia; Blakeslee, Joshua J.; Carraro, Nicola; Petrášek, Jan; Zažímalová, Eva; Hoyerová, Klára; Peer, Wendy Ann; Murphy, Angus S. (2012). „The Arabidopsis concentration-dependent influx/efflux transporter ABCB4 regulates cellular auxin levels in the root epidermis”. The Plant Journal. 69 (4): 640—654. ISSN 0960-7412. doi:10.1111/j.1365-313X.2011.04818.x.
- ^ Dawson RJ, Locher KP (2007). „Structure of the multidrug ABC transporter Sav1866 from Staphylococcus aureus in complex with AMP-PNP”. FEBS Lett. 581 (5): 935—8. PMID 17303126. doi:10.1016/j.febslet.2007.01.073.
- ^ Velamakanni S, Yao Y, Gutmann DA, van Veen HW (2008). „Multidrug transport by the ABC transporter Sav1866 from Staphylococcus aureus”. Biochemistry. 47 (35): 9300—8. PMID 18690712. doi:10.1021/bi8006737.
- ^ Reuter G, Janvilisri T, Venter H, Shahi S, Balakrishnan L, van Veen HW (2003). „The ATP binding cassette multidrug transporter LmrA and lipid transporter MsbA have overlapping substrate specificities”. J. Biol. Chem. 278 (37): 35193—8. PMID 12842882. doi:10.1074/jbc.M306226200.
- ^ Raetz CR, Reynolds CM, Trent MS, Bishop RE (2007). „Lipid A modification systems in gram-negative bacteria”. Annu. Rev. Biochem. 76: 295—329. PMC 2569861
- ^ а б Chang G, Roth CB (2001). „Structure of MsbA from E. coli: A homolog of the multidrug resistance ATP binding cassette (ABC) transporters”. Science. 293 (5536): 1793—800. PMID 11546864. doi:10.1126/science.293.5536.1793. (Retracted, see [Chang G, Roth CB, Reyes CL, Pornillos O, Chen Y, Chen AP (2006). „Retraction”. Science. 314 (5807): 1875. PMID 17185584. doi:10.1126/science.314.5807.1875b.])
- ^ а б Reyes CL, Chang G (2005). „Structure of the ABC transporter MsbA in complex with ADP•vanadate and lipopolysaccharide”. Science. 308: 1028—1031. PMID 15890884. doi:10.1126/science.1107733. (Retracted, see [Chang G, Roth CB, Reyes CL, Pornillos O, Chen Y, Chen AP (2006). „Retraction”. Science. 314 (5807): 1875. PMID 17185584. doi:10.1126/science.314.5807.1875b.])
- ^ Buchaklian AH, Funk AL, Klug CS (2004). „Resting state conformation of the MsbA homodimer as studied by site-directed spin labeling”. Biochemistry. 43 (26): 8600—6. PMID 15222771. doi:10.1021/bi0497751.
- ^ а б в Dong J, Yang G, McHaourab HS (2005). „Structural basis of energy transduction in the transport cycle of MsbA”. Science. 308 (5724): 1023—8. PMID 15890883. doi:10.1126/science.1106592.
- ^ Borbat PP, Surendhran K, Bortolus M, Zou P, Freed JH, Mchaourab HS (2007). „Conformational motion of the ABC transporter MsbA induced by ATP hydrolysis”. PLoS Biol. 5 (10): e271. PMC 2001213
- ^ Gutmann DA, Ward A, Urbatsch IL, Chang G, van Veen HW (2010). „Understanding polyspecificity of multidrug ABC transporters: closing in on the gaps in ABCB1”. Trends Biochem. Sci. 35 (1): 36—42. PMID 19819701. doi:10.1016/j.tibs.2009.07.009.
- ^ Lage, L (2009). „ABC Transporters as Target for RNA Interference-mediated Reversal of Multidrug Resistance”. ABC Transporters in Microorganisms. Caister Academic Press. ISBN 978-1-904455-49-3.
- ^ Akiyama, S. (2002). „[Mechanisms of drug resistance and reversal of the resistance]”. Hum. Cell. 14 (4): 257—60. PMID 11925925.
- ^ ATP-Binding Cassette Efflux Transporters and Passive Membrane Permeability in Drug Absorption and Disposition
- ^ Glavinas H, Krajcsi P, Cserepes J, Sarkadi B (2004). „The role of ABC transporters in drug resistance, metabolism and toxicity”. Curr Drug Deliv. 1 (1): 27—42. PMID 16305368. doi:10.2174/1567201043480036.
- ^ Glavinas H, Méhn D, Jani M, Oosterhuis B, Herédi-Szabó K, Krajcsi P (2008). „Utilization of membrane vesicle preparations to study drug-ABC transporter interactions”. Expert Opin Drug Metab Toxicol. 4 (6): 721—32. PMID 18611113. doi:10.1517/17425255.4.6.721.
- ^ Jeffrey P, Summerfield SG (2007). „Challenges for blood-brain barrier (BBB) screening”. Xenobiotica. 37 (10-11): 1135—51. PMID 17968740. doi:10.1080/00498250701570285.
- ^ This entire volume is dedicated to various methods used: Nikaido H, Hall J (1998). „ABC Transporters: Biochemical, Cellular, and Molecular Aspects”. Methods in enzymology. 292: 3—853. doi:10.1016/S0076-6879(98)92003-1.
- ^ Horio, M.; Gottesman, Michael M.; I, Pastan (1988). „ATP-dependent transport of vinblastine in vesicles from human multidrug-resistant cells”. Proc Natl Acad Sci USA. 85 (10): 3580—4. PMC 280257
- ^ Vesicular Transport Assay
- ^ „Human ATP-Binding Cassette Transporters”. Архивирано из оригинала 01. 06. 2014. г. Приступљено 01. 06. 2014.
- ^ Ueda, K.; DP, Clark; CJ, Chen; IB, Roninson; Gottesman, Michael M.; I, Pastan (1987). „The human multidrug resistance (mdr1) gene. cDNA cloning and transcription initiation”. J. Biol. Chem. 262 (2): 505—8. PMID 3027054.
- ^ Kerb R, Hoffmeyer S, Brinkmann U (2001). „ABC drug transporters: hereditary polymorphisms and pharmacological impact in MDR1, MRP1 and MRP2”. Pharmacogenomics. 2 (1): 51—64. PMID 11258197. doi:10.1517/14622416.2.1.51.
- ^ Fromm, M. F. (2003). „The influence of MDR1 polymorphisms on P-glycoprotein expression and function in humans”. Adv. Drug Deliv. Rev. 54 (10): 1295—310. PMID 12406646. doi:10.1016/S0169-409X(02)00064-9.
- ^ Dean, Michael (1. 11. 2002). „The Human ATP-Binding Cassette (ABC) Transporter Superfamily”. National Library of Medicine (US), NCBI. Pristupljeno 14. 3. 2019.
- ^ Brinkmann, U. (2002). „Functional polymorphisms of the human multidrug resistance (MDR1) gene: correlation with P glycoprotein expression and activity in vivo”. Novartis Found. Symp. Novartis Foundation Symposia. 243. ISBN 978-0-470-84635-3. PMID 11990778. doi:10.1002/0470846356.ch15.
- ^ Váradi A, Szakács G, Bakos E, Sarkadi B (2002). „P glycoprotein and the mechanism of multidrug resistance”. Novartis Found. Symp. Novartis Foundation Symposia. 243. ISBN 978-0-470-84635-3. PMID 11990782. doi:10.1002/0470846356.ch5.
- ^ а б Chen ZS, Tiwari AK (2011). „Multidrug resistance proteins (MRPs/ABCCs) in cancer chemotherapy and genetic diseases”. FEBS J. 278 (18): 3226—45. PMC 3168698
- ^ Gadsby DC, Vergani P, Csanády L (2006). „The ABC protein turned chloride channel whose failure causes cystic fibrosis”. Nature. 440 (7083): 477—83. Bibcode:2006Natur.440..477G. PMID 16554808. doi:10.1038/nature04712.
- ^ Rommens JM, Iannuzzi MC, Kerem B, Drumm ML, Melmer G, Dean M, Rozmahel R, Cole JL, Kennedy D, Hidaka N (1989). „Identification of the cystic fibrosis gene: chromosome walking and jumping”. Science. 245 (4922): 1059—65. Bibcode:1989Sci...245.1059R. PMID 2772657. doi:10.1126/science.2772657.
- ^ Riordan JR, Rommens JM, Kerem B, Alon N, Rozmahel R, Grzelczak Z, Zielenski J, Lok S, Plavsic N, Chou JL (1989). „Identification of the cystic fibrosis gene: cloning and characterization of complementary DNA”. Science. 245 (4922): 1066—73. Bibcode:1989Sci...245.1066R. PMID 2475911. doi:10.1126/science.2475911.
- ^ Childers M, Eckel G, Himmel A, Caldwell J (2007). „A new model of cystic fibrosis pathology: lack of transport of glutathione and its thiocyanate conjugates”. Med. Hypotheses. 68 (1): 101—12. PMID 16934416. doi:10.1016/j.mehy.2006.06.020.
- ^ Grangeia A, Sá R, Carvalho F, Martin J, Girodon E, Silva J, Ferráz L, Barros A, Sousa M (2007). „Molecular characterization of the cystic fibrosis transmembrane conductance regulator gene in congenital absence of the vas deferens”. Genet. Med. 9 (3): 163—72. PMID 17413420. doi:10.1097/GIM.0b013e3180318aaf.
- ^ Campbell JD, Sansom MS, Ashcroft FM (2003). „Potassium channel regulation”. EMBO Rep. 4 (11): 1038—42. PMC 1326373
- ^ Aguilar-Bryan L, Clement JP 4th, Gonzalez G, Kunjilwar K, Babenko A, Bryan J (1. 1. 1998). „=Toward understanding the assembly and structure of KATP channels”. Physiol Rev. 78 (1): 227—45. PMID 9457174.
- ^ Reis AF, Velho G (2002). „Sulfonylurea receptor -1 (SUR1): genetic and metabolic evidences for a role in the susceptibility to type 2 diabetes mellitus”. Diabetes Metab. 28 (1): 14—9. PMID 11938023.
- ^ „Entrez Gene: ABCD1 ATP-binding cassette, sub-family D (ALD), member 1”.
- ^ Moser, Hugo W.; Smith, Kirby D.; Watkins, Paul A.; Powers, James; Moser, Ann (2001). „131. X-Linked Adrenoleukodystrophy”. Ur.: Scriver, C.W.; Beaudet, A.L.; Sly, W.S.; Valle, D.; Childs, B.; Kinzler, K.W.; Vogelstein, B. Metabolic and Molecular Bases of Inherited Disease. 2 (8th izd.). New York: McGraw Hill. ISBN 978-0-07-136320-4.
- ^ Jinsheng Dong, Ruby Lai, Klaus Nielsen, Christie A. Fekete, Hongfang Qiu, and Alan G. Hinnebusch "The Essential ATP-binding Cassette Protein RLI1 Functions in Translation by Promoting Preinitiation Complex Assembly" J. Biol. Chem. 279: 42157-42168.
- ^ Richard M, Drouin R, Beaulieu AD (1998). „ABC50, a novel human ATP-binding cassette protein found in tumor necrosis factor-alpha-stimulated synoviocytes”. Genomics. 53 (2): 137—45. PMID 9790762. doi:10.1006/geno.1998.5480.
- ^ а б Allikmets R, Gerrard B, Hutchinson A, Dean M (1997). „Characterization of the human ABC superfamily: isolation and mapping of 21 new genes using the expressed sequence tags database”. Hum Mol Genet. 5 (10): 1649—55. PMID 8894702. doi:10.1093/hmg/5.10.1649.
- ^ Chen H, Rossier C, Lalioti MD, Lynn A, Chakravarti A, Perrin G, Antonarakis SE (1996). „Cloning of the cDNA for a human homologue of the Drosophila white gene and mapping to chromosome 21q22.3”. Am J Hum Genet. 59 (1): 66—75. PMC 1915121
- ^ Engel T, Bode G, Lueken A, Knop M, Kannenberg F, Nofer JR, Assmann G, Seedorf U (2006). „Expression and functional characterization of ABCG1 splice variant ABCG1(666)”. FEBS Lett. 580 (18): 4551—9. PMID 16870176. doi:10.1016/j.febslet.2006.07.006.
- ^ Saier, M. H. (2000). „A functional-phylogenetic classification system for transmembrane solute transporters”. Microbiol. Mol. Biol. Rev. 64 (2): 354—411. PMC 98997
- ^ „Structural View of Biology - Health and Disease”.
Literatura
[уреди | уреди извор]- Moser, Hugo W.; Smith, Kirby D.; Watkins, Paul A.; Powers, James; Moser, Ann (2001). „131. X-Linked Adrenoleukodystrophy”. Ur.: Scriver, C.W.; Beaudet, A.L.; Sly, W.S.; Valle, D.; Childs, B.; Kinzler, K.W.; Vogelstein, B. Metabolic and Molecular Bases of Inherited Disease. 2 (8th izd.). New York: McGraw Hill. ISBN 978-0-07-136320-4.
- Lage, L (2009). „ABC Transporters as Target for RNA Interference-mediated Reversal of Multidrug Resistance”. ABC Transporters in Microorganisms. Caister Academic Press. ISBN 978-1-904455-49-3.
- Scott MP, Lodish HF, Berk A, Kaiser, C, Krieger M, Bretscher A, Ploegh H, Amon A (2012). Molecular Cell Biology. San Francisco: W. H. Freeman. ISBN 978-1-4292-3413-9.
- Ponte-Sucre, A, ur. (2009). ABC Transporters in Microorganisms. Caister Academic Press. ISBN 978-1-904455-49-3.
- Szentpétery Z, Kern A, Liliom K, Sarkadi B, Váradi A, Bakos E (2004). „The role of the conserved glycines of ATP-binding cassette signature motifs of MRP1 in the communication between the substrate-binding site and the catalytic centers”. J Biol Chem. 279 (40): 41670—8. PMID 15252017. doi:10.1074/jbc.M406484200.
- Fitzgerald ML, Okuhira K, Short GF, Manning JJ, Bell SA, Freeman MW (2004). „ATP-binding cassette transporter A1 contains a novel C-terminal VFVNFA motif that is required for its cholesterol efflux and ApoA-I binding activities”. J Biol Chem. 279 (46): 48477—85. PMID 15347662. doi:10.1074/jbc.M409848200.
- Linton, K. J. (2011). The ABC Transporters of Human Physiology and Disease: The Genetics and Biochemistry of ATP Binding Cassette. World Scientific Publishing Company. ISBN 978-981-4280-06-8.
- SV, Ambudkar; Kimchi-Sarfaty C; ZE, Sauna; Gottesman, Michael M. (2003). „P-glycoprotein: from genomics to mechanism”. Oncogene. 22 (47): 7468—85. PMID 14576852. doi:10.1038/sj.onc.1206948.
- Jamroziak K, Robak T (2004). „Pharmacogenomics of MDR1/ABCB1 gene: the influence on risk and clinical outcome of haematological malignancies”. Hematology. 9 (2): 91—105. PMID 15203864. doi:10.1080/10245330310001638974.
- Ishikawa, T.; Y, Onishi; H, Hirano; et al. (2005). „Pharmacogenomics of drug transporters: a new approach to functional analysis of the genetic polymorphisms of ABCB1 (P-glycoprotein/MDR1)”. Biol. Pharm. Bull. 27 (7): 939—48. PMID 15256718. doi:10.1248/bpb.27.939.
- Lee W, Lockhart AC, Kim RB, Rothenberg ML (2005). „Cancer pharmacogenomics: powerful tools in cancer chemotherapy and drug development”. Oncologist. 10 (2): 104—11. PMID 15709212. doi:10.1634/theoncologist.10-2-104.
- Gambrelle, J.; S, Labialle; G, Dayan; et al. (2005). „Multidrug resistance in uveal melanoma”. Journal français d'ophtalmologie. 28 (6): 652—9. PMID 16141933.
- Al-Shawi MK, Omote H (2006). „The Remarkable Transport Mechanism of P-glycoprotein; a Multidrug Transporter”. J. Bioenerg. Biomembr. 37 (6): 489—96. PMC 1459968
- Orlowski S, Martin S, Escargueil A (2006). „P-glycoprotein and 'lipid rafts': some ambiguous mutual relationships (floating on them, building them or meeting them by chance?)”. Cell. Mol. Life Sci. 63 (9): 1038—59. PMID 16721513. doi:10.1007/s00018-005-5554-9.
- Annese, V.; MR, Valvano; O, Palmieri; et al. (2006). „Multidrug resistance 1 gene in inflammatory bowel disease: a meta-analysis”. World J. Gastroenterol. 12 (23): 3636—44. PMID 16773678.
- Sekine, I.; JD, Minna; K, Nishio; et al. (2007). „A literature review of molecular markers predictive of clinical response to cytotoxic chemotherapy in patients with lung cancer”. Journal of thoracic oncology : official publication of the International Association for the Study of Lung Cancer. 1 (1): 31—7. PMID 17409824.
- Kumar YS, Adukondalu D, Sathish D, Vishnu YV, Ramesh G, Latha AB, Reddy PC, Sarangapani M, Rao YM (2010). „P-Glycoprotein- and cytochrome P-450-mediated herbal drug interactions”. Drug Metabol Drug Interact. 25 (1-4): 3—16. PMID 21417789. doi:10.1515/DMDI.2010.006.
Spoljašnje veze
[уреди | уреди извор]- Classification of ABC transporters in TCDB
- ABCdb Архивирано на сајту Wayback Machine (14. март 2023) Archaeal and Bacterial ABC Systems database, ABCdb
- ATP-Binding+cassette+transporters на US National Library of Medicine Medical Subject Headings (MeSH)
| Normativna kontrola: Državne |
|---|