Rakutuum

1. Tuumake
2. Tuum
3. Ribosoom
4. Vesiikul
5. Karedapinnaline tsütoplasmavõrgustik
6. Golgi kompleks
7. Tsütoskelett
8. Siledapinnaline tsütoplasmavõrgustik
9. Mitokonder
10. Vakuool
11. Tsütoplasma
12. Lüsosoom
13. Tsentrosoom
Rakutuum on kahekihilise membraaniga ümbritsetud rakuorganell, mis esineb tsütoplasmas vaid eukarüootidel. Rakutuum hõlmab umbes 10% raku kogu ruumalast ja sinna on koondunud peaaegu kogu rakus olev geneetiline materjal.[1] Tuumas sisalduv DNA on tihedalt kokku pakitud histoonide abil ja moodustab kromosoomi. Rakutuuma ülesandeks on säilitada ja edasi anda geneetilist materjali ning kontrollida raku elutegevust läbi geeniekspressiooni.Enamikul eukarüootilistest rakkudest on üks tuum. See kehtib paljude loomade ja taimede rakkude kohta ning ka inimese kehas leiduvate rakkude kohta.Raku tuum esineb eukarüootsetes rakkudes. Sest see hoiab ja kaitseb raku geneetilist materjali. Rakutuuma katab tuumaümbris, mille välimine membraan on ühenduses tsütoplasmavõrgustikuga. Sisemist membraani toetavad seestpoolt tuuma lamiinid. Sisemise ja välimise tuumamembraani vahele jääb perinukleaarne ruum. Tuumamembraani läbivad teatud vahemike järel tuuma poori kompleksid, mis reguleerivad karüoplasma ja tsütoplasma vahelist transporti. Kompleksi suudavad passiivse transpordi teel läbida ioonid ja väikesed molekulid ning aktiivse transpordi abil suuremad molekulid. Tuumas esineb ka erinevaid intranukleaarseid kehi, millest tähtsaim ja ka mikroskoobis enim märgatav on tuumake. Tuumakese peamine ülesanne on ribosoomi subühikute moodustamine rRNA-st ja valkudest.Mõned rakutuuma ülesanded on geneetilise materjali säilitamine, signaaliülekandmine, ribosoomide tootmine, raku jagunemine.
Ajalugu
[muuda | muuda lähteteksti]
Arvatakse, et rakutuuma avastas Antonie van Leeuwenhoek (1632–1723), sest vanim pilt tuumast, mis on säilinud, oli tema joonistatud. Oma pildil kujutas ta lõhe punase vereraku tuuma, mis erinevalt imetaja erütrotsüüdist sisaldab tuuma.[2] Detailsemalt kujutas rakutuuma aga 1831. aastal botaanik Robert Brown. Ta uuris mikroskoobi all orhideesid ja avastas läbipaistmatu ala taime pindmises kihis ning nimetas selle tuumaks.[3] Brown ei suutnud siiski selgitada tuuma rolli raku elutsüklis. Tuuma funktsiooni pakkus esimesena välja 1838. aastal Matthias Schleiden, kes arvas, et tuum on kuidagi seotud uute rakkude moodustumisega. Alles 20. sajandi alguses seoses mitoosiprotsesside avastamisega ja Mendeli seaduste taasavastamisega selgus, et tuum on geneetilise info kandja.
Ehitus
[muuda | muuda lähteteksti]Raku tuum on ümbritsetud kahekihilise membraaniga, mida nimetatakse tuumamembraaniks.Loomorganismides on tuum kõige suurem rakusisene organell.[4] Imetajate rakkudes on tuuma läbimõõt keskmiselt 6 mikromeetrit (μm).[5] Rakutuum sisaldab karüoplasmat, mis on tsütoplasmasarnane viskoosne vedelik.[6] Tuumal on kindel koht raku sees, kuid selle täpne asukoht võib erineda sõltuvalt rakutüübist. Tuum on tavaliselt tsentraalses või veidi nihkes positsioonis, kuid mõnel juhul võib see olla ka äärmistes piirkondades.
Tuumaümbris ja poorid
[muuda | muuda lähteteksti]Tuumaümbris, mida teatakse ka tuumamembraani nime all, koosneb kaksikmembraanist, kus need on üksteisega paralleelsed ja asetsevad teineteisest 10–50 nanomeetri (nm) kaugusel. Tuum on täielikult ümbritsetud tuumamembraaniga, mis eraldab geneetilist materjali tsütoplasmast ja kaitseb makromolekulide sissetungimise eest tsütoplasmast karüoplasmasse.[7]
Tuumapoorid
[muuda | muuda lähteteksti]Tuumaümbrist läbistavad teatud vahemaade tagant umbes 100 liiki valkudest (selgroogsetel) koosnevad tuuma poori kompleksid (nuclear pore complex – NPC). Keskmiselt on imetajatel rakutuumas 3000–4000 poori.[4] Kompleksi moodustavaid valke nimetatakse ka nukleoporiinideks. NPC-d omavad oktagonaalset sümmeetriat ning sellise kompleksi molekulmass on umbes 125 miljonit daltonit. Poori läbimõõt on keskmiselt 100 nm. Kanali sisemine läbimõõt on selles oleva regulatoorse süsteemi tõttu vaid 9 nm. Seetõttu ei saa makromolekulid ilma aktiivse transpordi abita tsütoplasmast tuuma liikuda.[8]
Rakutuumas sünteesitakse suurel hulgal erinevaid makromolekule (ribosoomide subühikud, mRNA, tRNA), mis talitlevad tsütoplasmas. Samas moodustatakse tsütoplasmas erinevaid valke (DNA- ja RNA-polümeraase, geenide regulaatorvalke), mis peavad lõpuks jõudma rakutuuma. Üks ja seesama tuumapoor on võimeline molekule transportima mõlemasuunaliselt.
Valkude transport läbi tuumapoori
[muuda | muuda lähteteksti]Selleks, et makromolekulid (molekulmassiga üle 60 000 Da) saaksid liikuda tuuma ja tsütoplasma vahel, on vajalikud tuuma poori kompleksid, aktiivse transpordi läbiviimine ning teatud signaalide olemasolu, mis lubaks transporti läbi viia. Lisaks on vajalik ka teatud temperatuur ja ATP energia olemasolu. Passiivse difusiooni teel saavad tuumapoorist läbi liikuda vaid ioonid ja väikesed valgu molekulid (molekulmass alla 5000 Da).
Importiiniks kutsutaks üht valku viiest, mis tunneb ära NLS-järjestusega karüofiilse valgu ning seob selle tuumapoori valkudele. Valkude pääsemiseks läbi tuumapoori karüoplasmasse on vajalik teatud aminohappeline (AH) järjestus, mis sisaldab tavaliselt Arg-i ja Lys-i. Sellist AH järjestust nimetatakse nukleaarse lokalisatsiooni signaaliks (NLS), see on lühike 5–12 AH-d sisaldav järjestus. Esineb kaht peamist tüüpi NLS: kas lühikesed nelja- kuni seitsmealuselisest aminohappest koosnevad järjestused või pikemad kaheosalised NLS-id, mis koosnevad kahest aluseliste aminohapete plokist, mida eraldab umbes kümme aminohapet.
Ka valkude transpordiks tuumast tsütoplasmasse on vajalikud teatud signaaljärjestused, kuid neid kutsutakse NES (nuclear export signal) signaalideks. NES-signaaljärjestuses on vajalikud hüdrofoobsed aminohapped (Leu ja Ile), mis esinevad teatud kindlas asetuses. Enamik valke, mis sisaldavad NLS-signaali, sisaldavad ka NES-signaaljärjestust, sest valke, mida tuumas enam vaja ei ole, ei ole võimalik tuumas lagundada ja sellised valgud tuleb transportida tagasi tsütoplasmasse.[9]
Tuumake
[muuda | muuda lähteteksti]Tuumakeseks nimetatakse tuumas asetsevat häguste piiridega eristatavat moodustist, millel puudub membraan. Tuumake moodustub tuumakese organisaatorpiirkonna (NOR) ümber, see on koht kromosoomis, mis sisaldab rRNA e ribosomaalse RNA geene. NOR-id paiknevad kromosoomi lühikeste õlgade otstes. Tuumakese peamiseks ülesandeks on rRNA süntees ja ribosoomi subühikute moodustamine. Inimesel esineb diploidses rakus 10 NOR-i ja neis paikneb tihedalt koos umbes 200 rRNA geeni. Kui teoreetiliselt võiks tuumas esineda 10 tuumakest, siis tavaliselt moodustavad nad tihedalt koos ühe suure tuumakese. Ka ei toimu kõigilt geenidelt samaaegne rRNA transkriptsioon ning seetõttu on tuumakesi vähem nähtaval.[10]
Elektronmikroskoopiliselt on võimalik eristada tuumakeses 3 piirkonda: 1. tihe fibrillaarne komponent, mis sisaldab sünteesitavat RNA-d; 2. granulaarne komponent, mis sisaldab formeeruvaid ribosoomi partikleid; 3. fibrillaarne tsenter, mis sisaldab DNA-d mida parasjagu ei transkribeerita.
Tuumakese suurus oleneb sellest, kui suures mahus toimub transkriptsioon. Rakkudes, kus toimub intensiivne valkude süntees, võib tuumake hõivata tuuma ruumalast kuni 25%.[11]
Esimese sammuna moodustatakse RNA polümeraas I abiga 45S pre-rRNA, millest edasise töötluse teel saadakse 4 erinevat rRNA-d (28 S, 5.8 S, 5S ja 18 S RNA), mis liidetakse, kas ribosoomi suure (60S) või väikese (40S) subühiku koosseisu.[11]
Kromosoomid
[muuda | muuda lähteteksti]Pikemalt artiklis Kromosoom
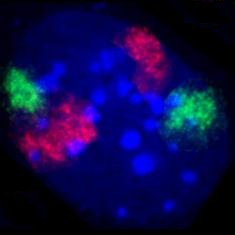
Enamik raku DNAst on koondunud kromosoomidesse, mis asetsevad rakutuumas. Igas inimese rakus on umbes 2 m DNAd. Suurema osa rakutsüklist on kromosoomid dekondenseerunud ning esineb valkude ja DNA kompleks, mida nimetatakse kromatiiniks. Mitoosi või meioosi ajal kromosoomid kondenseeruvad ja muutuvad nähtavaks valgusmikroskoobist vaadates. Metafaasi aegsete kromosoomide kuju, suurus ja arv määravad karüotüübi, mis on igale liigile iseloomulik. Interfaasi ajal ei paikne kromosoomid mitte juhuslikult, vaid neil on oma kindlad territooriumid.[12] Kromosoomi osad, millelt toimub aktiivne transkriptsioon, võivad oma kohalt nihkuda. Väike hulk DNA-st on koondunud ka mitokondrisse.
Kromatiini saab jagada vastavalt oma ehitusele 2 alatüübiks. Kromatiini osa, mis on nähtav valgusmikroskoobis, nimetatakse eukromatiiniks. Eukromatiin sisaldab geene, mida transkribeeritakse tihedamini ning seetõttu paikneb eukromatiin ka hajusamalt ja tuuma keskel. Kromatiini piirkond, kus DNA on rohkem kokkupakitud, nimetatakse heterokromatiiniks. Heterokromatiin sisaldab vähesel määral geene. Peamiselt koosneb ta kõrgelt korduvatest DNA järjestustest, mis vastutavad kromosoomi struktuursete osade nagu telomeeride ja tsentromeeride moodustamise eest. Seetõttu nimetatakse seda osa kromatiinist ka konstitutiivseks heterokromatiiniks.[13] Kuna heterokromatiini aladelt ei toimu tihedat traskriptsiooni, siis paikneb see peamiselt membraani läheduses.
Tuuma funktsioon
[muuda | muuda lähteteksti]Tuuma peamiseks ülesandeks on geeni ekspressiooni, DNA replikatsiooni reguleerimine ja kontroll. Tuum võimaldab hoida eukarüootidel DNA replikatsiooni ja RNA transkriptsiooni eraldatud tsütoplasmas olevast translatsioonist. See võimaldab prokarüootidest efektiivsemat geeni regulatsiooni.
Geeni ekspressioon
[muuda | muuda lähteteksti]Geeni ekspressiooni esimene etapp on transkriptsioon, mille käigus DNA ahela abil sünteesitakse RNA ahel. Selle tulemusena moodustub mRNA, mis peab läbima translatsiooni, et moodustuks valk. Enne translatsiooni algust peab mRNA liikuma tuumast tsütoplasmasse.[14]
Tuum sisaldab transkriptsiooni läbiviimiseks vajalikke valke, näiteks helikaasi, RNA polümeraasi ja topoisomeraasi, lisaks veel erinevaid traskriptsioonifaktoreid. Helikaas vastutab DNA kaksikahela lahtikeeramise eest, et RNA polümeraas saaks sellele kinnituda. RNA polümeraas sünteesib kasvavat RNA molekuli. Topoisomeraas keerab DNA ahelat lahti ja kinni.[15]
Pre-mRNA töötlus
[muuda | muuda lähteteksti]Transkriptsiooni tulemusel saadakse pre-mRNA. Seejärel peab see läbima töötluse tuumas, mille käigus saadakse mRNA, mis saab liikuda tsütoplasmasse. Kui mRNA ei läbi vajalikke modifikatsioone, siis selline mRNA lagundatakse tsütoplasmas ning seda ei kasutata translatsiooniks.
Kolm peamist töötluse etappi on 1) 5-otsa7-metüülguanosiini mütsi lisamine, mis on töötluse esimene etapp 2) 3-otsapol(A)saba lisatakse pärast transkriptsiooni lõppu 3) RNA splaissimine, mille käigus splaissosoom eemaldab intronid ning liidab kokku järelejäänud eksonid. Imetajate geenides on aga võimalik eksone ühendada alternatiivse splaissingu abil mitut moodi. Sel juhul on ühelt pre-mRNAlt võimalik saada erinevaid produkte.
Mitme tuumaga ja tuumatud rakud
[muuda | muuda lähteteksti]Kuigi enamikul rakkudest esineb vaid üks tuum, on eukarüootide hulgas ka rakutüüpe, mis on tuumatud või hulktuumsed. Tuumatud rakud on näiteks imetajate küpsed punased vererakud e erütrotsüüdid, mis ei sisalda ka mitokondreid. Samas võib tuuma puudumine olla ka vigase rakujagunemise tagajärg. Tuumatud rakud ei ole võimelised jagunema.
Hulgituumsed rakud on enamikul Acantharia liikidel ja mükoriisadel. Kaks tuuma raku kohta on aga sooleparasiitidel Giardiatel. Arengu käigus on hulktuumsed ka inimese skeletilihaseid moodustavad rakud.
Vaata ka
[muuda | muuda lähteteksti]Viited
[muuda | muuda lähteteksti]- ↑ [ http://cellbio.ebc.ee/rakubio/tuum.html], konspekt.
- ↑ Leeuwenhoek, A. van: Opera Omnia, seu Arcana Naturae ope exactissimorum Microscopiorum detecta, experimentis variis comprobata, Epistolis ad varios illustres viros. J. Arnold et Delphis, A. Beman, Lugdinum Batavorum 1719–1730. Cited after: Dieter Gerlach, Geschichte der Mikroskopie. Verlag Harry Deutsch, Frankfurt am Main, Germany, 2009. ISBN 978-3-8171-1781-9.
- ↑ Brown, Robert (1866). "On the Organs and Mode of Fecundation of Orchidex and Asclepiadea". Miscellaneous Botanical Works I: 511–514.
- ↑ 4,0 4,1 Lodish, H; Berk A, Matsudaira P, Kaiser CA, Krieger M, Scott MP, Zipursky SL, Darnell J. (2004). Molecular Cell Biology (5th ed.). New York: WH Freeman. ISBN 0-7167-2672-6.
{{cite book}}: CS1 hooldus: mitu nime: autorite loend (link) - ↑ Bruce Alberts, Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, Peter Walter, toim (2002). Molecular Biology of the Cell, Chapter 4, pages 191–234 (4th ed.). Garland Science.
{{cite book}}: CS1 hooldus: mitu nime: toimetajate loend (link) - ↑ Clegg JS (veebruar 1984). "Properties and metabolism of the aqueous cytoplasm and its boundaries". Am. J. Physiol. 246 (2 Pt 2): R133–51. PMID 6364846.
- ↑ Paine P, Moore L, Horowitz S (1975). "Nuclear envelope permeability". Nature. 254 (5496): 109–114. DOI:10.1038/254109a0. ISSN 0028-0836. PMID 1117994.
{{cite journal}}: CS1 hooldus: mitu nime: autorite loend (link)</ref Välimine tuumamembraan on otseses ühenduses karedapinnalise tsütoplasmavõrgustiku membraaniga ning sarnaselt sellega on ka tuumamembraani välimisel pinnal ribosoom.</ref name="Paine">WERNER W. FRANKE, ULRICH SCHEER, GEORG KROHNE, and ERNST-DIETER JARASCH (1981). "The Nuclear Envelope and the Architecture of the Nuclear Periphery". The Journal of Cell Biology. 91 (3 Pt 2): 39–50. PMC 2112808. PMID 7033243.{{cite journal}}: CS1 hooldus: mitu nime: autorite loend (link) - ↑ Rodney Rhoades, Richard Pflanzer, toim (1996). "Ch3". Human Physiology (3rd ed.). Saunders College Publishing.
- ↑ [1], 05.10.2012.
- ↑ Hernandez-Verdun, Daniele (2006). "Nucleolus: from structure to dynamics". Histochem. Cell. Biol. 125 (1–2): 127–137. DOI:10.1007/s00418-005-0046-4. PMID 16328431.
- ↑ 11,0 11,1 Bruce Alberts, Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, Peter Walter, toim (1989). Molecular Biology of the Cell, Chapter 9, pages 541–543 (2th ed.). Garland Publishing.
{{cite book}}: CS1 hooldus: mitu nime: toimetajate loend (link) - ↑ Lamond, Angus I.; William C. Earnshaw (24. aprill 1998). "Structure and Function in the Nucleus". Science. 280 (5363): 547–553. DOI:10.1126/science.280.5363.547. PMID 9554838.
- ↑ Grigoryev S, Bulynko Y, Popova E (2006). "The end adjusts the means: heterochromatin remodelling during terminal cell differentiation". Chromosome Res. 14 (1): 53–69. DOI:10.1007/s10577-005-1021-6. PMID 16506096.
{{cite journal}}: CS1 hooldus: mitu nime: autorite loend (link) - ↑ Nierhaus, Knud H.; Daniel N. Wilson (2004). Protein Synthesis and Ribosome Structure: Translating the Genome. Wiley-VCH. ISBN 3-527-30638-2.
- ↑ Nicolini, Claudio A. (1997). Genome Structure and Function: From Chromosomes Characterization to Genes Technology. Springer. ISBN 0-7923-4565-7.
| Pildid, videod ja helifailid Commonsis: Rakutuum |