Sinapomorfia

Una sinapomorfia o sinapomorfía (del griego σύν- syn- 'con', ἀπό- apó- = en la distancia μορφή morphḗ 'forma') es un carácter homólogo apomórfico (es decir, una novedad evolutiva) compartido por todos los individuos de un taxón.[1] Es decir, una sinapomorfía es una novedad evolutiva que permite diferenciar a un taxón de otros taxones. En Biología sistemática, los grupos monofiléticos (que incluyen a un ancestro y todos sus descendientes) de organismos pueden ser reconocidos a través de aquellos caracteres que se originaron en su ancestro común, que son por lo tanto compartidos por todo el grupo (incluido el ancestro común), es decir, por las sinapomorfias. El concepto fue primero formalizado por Hennig (1966) y Wagner (1980) y está en el centro de la metodología del análisis filogenético.[2]
La sinapomorfía se opone tanto a la simplesiomorfia como a la homoplasia: las simplesiomorfias son caracteres primitivos compartidos que se originaron en el ancestro común más reciente; las homoplasias son caracteres superficialmente similares, pero evolucionados independientemente.
Las sinapomorfias se constituyen así en el fundamento para la clasificación filogenética (la basada en el parentesco) de los seres vivos. Gracias a ellas se definen grupos holofiléticos (monofiléticos en el sentido de Willi Hennig). Los caracteres con que se describen grupos parafiléticos, definidos por la exclusión de un grupo holofilético de otro mayor, son simplesiomorfias. Solo las sinapomorfias, no las simplesiomorfías, constituyen argumentos válidos en favor de la monofilia (holofilia) del grupo de taxones que la comparten.
Los conceptos de apomorfía y de plesiomorfía son relativos a un cierto nivel de generalidad, es decir, que lo que aparece como apomorfía en un determinado nivel de generalidad puede ser una plesiomorfía en otro nivel. Por ejemplo, en las ratas y los monos, los cinco dedos del pie constituyen una simplesiomorfia, pero en los tetrápodos en general el rasgo puede ser una sinapomorfia.
Las sinapomorfias en cladística
[editar]En cladística, las sinapomorfías son caracteres útiles para construir la filogenia del grupo estudiado. Son ellas las que definen los clados (grupos monofiléticos). Para ello, al construir el cladograma del grupo estudiado, las sinapomorfias se agregan al gráfico como una marquita en la rama del ancestro común, justo debajo de la primera ramificación del clado (ver gráfico). No es necesario que las sinapomorfias estén presentes en todos los miembros de un clado, pues algunos pueden haberla perdido en forma secundaria a su adquisición. También algunos pueden poseer la sinapomorfia en forma muy modificada (por ejemplo es una sinapomorfia el polen tricolpado del clado de los tricolpados, hoy llamados Eudicotyledoneae, aun cuando el clado posee muchas formas derivadas de polen).
Hay tres formas de definir un clado, algunos lo definen a partir de la aparición de la apomorfia, otros a partir de que se ramificó del clado más abarcativo a este, y otros a partir de la ramificación en más subclados:
Las sinapomorfías son una de las 3 formas de definir un clado Según el PhyloCode. |
Aquí un ejemplo real de un cladograma con sus sinapomorfias marcadas:
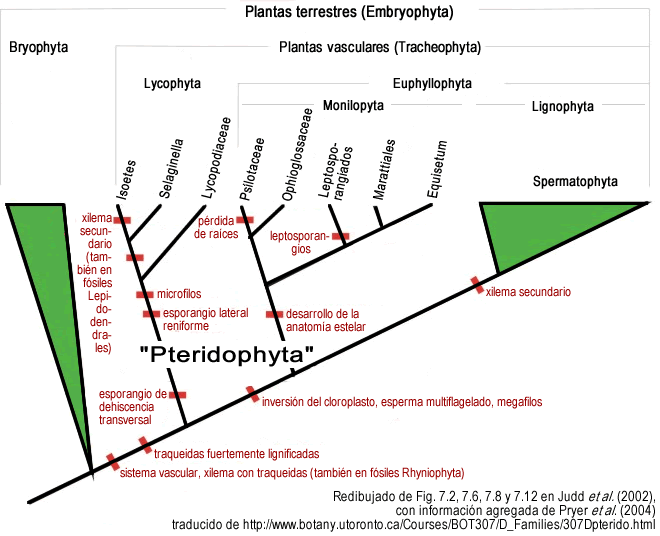 Ejemplo de cladograma al que se le han agregado las sinapomorfías Las sinapomorfías se marcan con una barrita que "corta" a la rama en el lugar en que se hipotetiza que fue originada. En este caso la barrita fue pintada de rojo oscuro. |
Nótese que las sinapomorfias marcadas en el cladograma del ejemplo son sólo sinapomorfias para los taxones que están por arriba de la sinapomorfia, y que son afectados por ella. Por ejemplo, es incorrecto decir que los megafilos ("hojas grandes") son una sinapomorfia de las espermatofitas: los megafilos son una sinapomorfia de todo el clado que está por arriba de ellos, que son las eufilofitas (de hecho los megafilos son una simplesiomorfia para las espermatofitas, ya que son un carácter ancestral más abarcativo, compartido por ese clado y otros más).
Sinapomorfia y homoplasia
[editar]¿Cómo se dan cuenta los científicos de qué caracteres son sinapomorfias que definen un clado y qué caracteres fueron originados muchas veces en forma independiente, y no definen un clado? No es exagerado decir que antes dependían más de la intuición de los biólogos sistemáticos que de otra cosa. Algunas cosas ayudan: si el carácter es complicado de construir en términos genéticos (es decir, si necesita muchas mutaciones o mutaciones muy improbables) entonces probablemente ese carácter haya sido originado una sola vez en la historia de los seres vivos, y sea una sinapomorfía, compartido solo por el ancestro común donde se originó una única vez, y sus descendientes.
Hoy en día el término es más utilizado en los análisis moleculares del ADN que en los morfológicos: las mutaciones improbables del ADN son sospechadas como sinapomorfias (es "improbable" una inversión del ADN de muchas kilobases, una pérdida o duplicación de una parte importante de un cromosoma, el traslado de un sector importante —muy grande— de un cromosoma a otro cromosoma, entre otros). Una vez el árbol filogenético basado en el estudio molecular del ADN está delineado, se buscan a posteriori las sinapomorfias morfológicas. Se priorizan los caracteres del ADN por sobre los morfológicos para buscar sinapomorfias, porque los caracteres morfológicos para ser moldeados, además de depender del ADN, dependen de la presión de selección natural, por lo que hay una gran cantidad de caracteres análogos (que son morfológicamente similares porque fueron seleccionados por la presión de selección, pero no tienen el mismo origen genético, y no comparten un antecesor común) que dificultan la búsqueda de sinapomorfias. Un ejemplo es el tallo suculento de las cactáceas y el de las euforbiáceas, que fueron originados independientemente por la presión de selección natural que los adaptó a sobrevivir en ambientes desérticos.
Ejemplos
[editar]- El doble aparato mandibular de los cíclidos es una sinapomorfia para este clado. Su aparición permitió una masiva proliferación de los cíclidos en los lagos africanos.
- La reducción de las alas posteriores de los dípteros para originar los halterios son una apomorfia compartida (sinapomorfia) por todos los miembros del orden Diptera, que por tanto es un taxón monofilético.
- La presencia de pelo es una sinapomorfia de los mamíferos, aunque algunos grupos (cetáceos) lo hayan perdido posteriormente en el curso de la evolución.
Véase también
[editar]Referencias
[editar]- ↑ Currie, Philip J.; Padia, Kevin (1997). Encyclopedia of Dinosaurs. Elsevier. p. 543. ISBN 978-0-08-049474-6. Parámetro desconocido
|name-list-format=ignorado (ayuda) - ↑ Barton, Nicholas; Briggs, Derek; Eisen, Jonathan; Goldstein, David; Patel, Nipam (2007). «Phylogenetic Reconstruction». Evolution. Cold Spring Harbor Laboratory Press. Parámetro desconocido
|name-list-format=ignorado (ayuda)
- Judd, W. S. Campbell, C. S. Kellogg, E. A. Stevens, P.F. Donoghue, M. J. 2002. Plant systematics: a phylogenetic approach, Second Edition. Sinauer Axxoc, USA.
- PhyloCode ( https://web.archive.org/web/20030802181518/http://www.phylocode.org/ )
- Cladistics, Berkeley
| Control de autoridades |
|
|---|
Datos: Q962872