Código genético
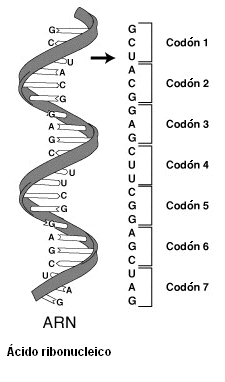
El código genético es el conjunto de reglas que define cómo se traduce una secuencia de nucleótidos en el ARN a una secuencia de aminoácidos en una proteína. Este código es común en todos los seres vivos y todos los virus (aunque hay pequeñas variaciones), lo cual demuestra que ha tenido un origen único y es universal, al menos en el contexto de nuestro planeta.[1]
El código define la relación entre cada secuencia de tres nucleótidos, llamada codón, y cada aminoácido.
La secuencia del material genético se compone de cuatro bases nitrogenadas distintas, que tienen una representación mediante letras en el código genético: adenina (A), timina (T), guanina (G) y citosina (C) en el ADN y adenina (A), uracilo (U), guanina (G) y citosina (C) en el ARN.[2]
Debido a esto, el número de codones posibles es 64,[3] de los cuales 61 codifican aminoácidos (siendo además uno de ellos el codón de inicio, AUG) y los tres restantes son sitios de parada (UAA, llamado ocre; UAG, llamado ámbar; UGA, llamado ópalo).[4] La secuencia de codones determina la secuencia de aminoácidos en una proteína en concreto, que tendrá una estructura y una función específica.
Descubrimiento del código genético
[editar]
Cuando Francis Crick, Rosalind Franklin, James Watson y Maurice Wilkins presentaron el modelo de la estructura del ADN se comenzó a estudiar en profundidad el proceso de traducción en las proteínas.
En 1955, Severo Ochoa y Marianne Grunberg-Manago aislaron la enzima polinucleótido fosforilasa,[5] capaz de sintetizar ARNm sin necesidad de molde a partir de cualquier tipo de nucleótidos que hubiera en el medio. Así, a partir de un medio en el cual tan solo hubiera UDP (uridína difosfato) se sintetizaba un ARNm en el cual únicamente se repetía el ácido uridílico, es decir, se generaba un polipéptido de un solo nucleótido, un poli-U.
George Gamow postuló que el código genético estaría formado por tripletes de cuatro bases nitrogenadas (A;U;C;G) y que a partir de estas se formarían los veinte aminoácidos esenciales para la vida. La primera demostración de que los codones constan de tres nucleótidos la proporcionó el experimento de Crick, Brenner y colaboradores. Marshall Warren Nirenberg y Heinrich J. Matthaei en 1961 en los Institutos Nacionales de Salud descubrieron la primera correspondencia codón-aminoácido. Empleando un sistema sin células, tradujeron una secuencia ARN de poli-uracilo (UUU...) y descubrieron que el polipéptido que se sintetizaba solo contenía fenilalanina. De esto se deduce que el codón UUU especifica el aminoácido fenilalanina. Continuando con el trabajo anterior, Marshall Warren Nirenberg y Philip Leder fueron capaces de determinar la traducción de 54 codones, utilizando diversas combinaciones de ARNm, pasadas a través de un filtro de ribosomas. Los ARNt se unían a tripletes específicos.
Posteriormente, Har Gobind Khorana completó el código, y poco después, Robert W. Holley determinó la estructura del ARN de transferencia, la molécula adaptadora que facilita la traducción. Este trabajo se basó en estudios anteriores de Severo Ochoa, quien recibió el premio Nobel en 1959 por su trabajo en la enzimología de la síntesis de ARN. En 1968, Khorana, Holley y Nirenberg recibieron el Premio Nobel en Fisiología o Medicina por su trabajo.
Transferencia de información
[editar]El genoma de un organismo se encuentra en el ADN[6] o, en el caso de algunos virus, en el ARN. La porción de genoma que codifica a una proteína o un ARN se conoce como gen.[7] Esos genes que codifican proteínas están compuestos por unidades de trinucleótidos llamadas codones, cada una de los cuales codifica un aminoácido. Cada nucleótido está formado por un fosfato, una desoxirribosa y una de las cuatro posibles bases nitrogenadas. Las bases purínicas adenina (A) y guanina (G) son más grandes y tienen dos anillos aromáticos. Las bases pirimidínicas citosina (C) y timina (T) son más pequeñas y solo tienen un anillo aromático. En la configuración en doble hélice, dos cadenas de ADN están unidas entre sí por puentes de hidrógeno en una asociación conocida como emparejamiento de bases. Además, estos puentes siempre se forman entre una adenina de una cadena y una timina de la otra y entre una citosina de una cadena y una guanina de la otra. Esto quiere decir que el número de residuos A y T será el mismo en una doble hélice y lo mismo pasará con el número de residuos de G y C. En el ARN, la timina (T) se sustituye por uracilo (U), y la desoxirribosa por una ribosa.
Cada gen que codifica una proteína se transcribe en una molécula plantilla, que se conoce como ARN mensajero o ARNm. Este, a su vez, se traduce en el ribosoma, en una cadena polipeptídica (formada por aminoácidos). En el proceso de traducción se necesita un ARN de transferencia, o ARNt, específico para cada aminoácido, con dicho aminoácido unido a él de forma covalente, guanosina trifosfato como fuente de energía y ciertos factores de traducción. Los ARNt tienen anticodones complementarios a los codones del ARNm y se pueden “cargar” covalentemente en su extremo 3' terminal con aminoácidos. Los ARNt individuales se cargan con aminoácidos específicos gracias a las enzimas llamadas aminoacil-ARNt sintetasas, que tienen alta especificidad tanto por un aminoácido como por un ARNt. Esta alta especificidad es el motivo fundamental del mantenimiento de la fidelidad en la traducción de proteínas.
Para un codón de tres nucleótidos (un triplete) son posibles 4³ = 64 combinaciones diferentes; los 64 codones están asignados a un aminoácido o a señales de parada en la traducción. Si, por ejemplo, tenemos una secuencia de ARN, UUUAAACCC, y la lectura del fragmento empieza en la primera U (convenio 5' a 3'), habría tres codones que serían UUU, AAA y CCC, cada uno de los cuales especifica un aminoácido. Esta secuencia de ARN se traducirá en una secuencia de tres aminoácidos específicos.
Características
[editar]Universalidad
[editar]El código genético es compartido por los organismos conocidos, incluyendo virus y orgánulos, aunque pueden aparecer pequeñas diferencias. Así, por ejemplo, el codón UUU codifica el aminoácido fenilalanina tanto en bacterias como en arqueas y en eucariontes. Este hecho indica que el código genético ha tenido un origen único en todos los seres vivos conocidos. La palabra «universal» en este contexto se aplica solamente a la vida en la Tierra, ya que no se ha establecido la existencia de vida en otro planeta.
Gracias a la genética molecular, se han distinguido 22 códigos genéticos,[1] que se diferencian del llamado código genético estándar por el significado de uno o más codones. La mayor diversidad se presenta en las mitocondrias, orgánulos de las células eucariotas que poseen su propio ADN separado del núcleo. El genoma del núcleo de algunas pocas eucariotas solo se diferencia del código estándar en los codones de iniciación y terminación.
Especificidad y continuidad
[editar]Ningún codón codifica más de un aminoácido; de no ser así, conllevaría problemas considerables para la síntesis de proteínas específicas para cada gen. Tampoco presenta solapamiento: los tripletes se hallan dispuestos de manera lineal y continua, de manera que entre ellos no existan ni comas ni espacios y sin compartir ninguna base nitrogenada. Su lectura se hace en un solo sentido (5' - 3'), desde el codón de iniciación hasta el codón de parada. Sin embargo, en un mismo ARNm pueden existir varios codones de inicio, lo que conduce a la síntesis de varios polipéptidos diferentes a partir del mismo transcrito.
Degeneración
[editar]El código genético tiene redundancia, pero no ambigüedad. Por ejemplo, aunque los codones GAA y GAG especifican ambos el mismo ácido glutámico, ningún codón especifica dos aminoácidos distintos. Las diferencias entre los codones que codifican un mismo aminoácido presentan diferencias en la tercera posición. Es por ello que, de forma general, la tolerancia al cambio en esta posición es mayor que en la primera y segunda, y por tanto tiende a estar menos representada en el caso de variaciones que resultan en patologías (situación que se concentra fundamentalmente en el primer nucleótido del codón).[8] Debido a que las mutaciones de transición (purina a purina o pirimidina a pirimidina) son más probables que las de transversión (purina a pirimidina o viceversa), la equivalencia de purinas o de pirimidinas en los lugares dobles degenerados añade una tolerancia complementaria a fallos.
Agrupamiento de codones por residuos aminoacídicos, volumen molar e hidropatía
[editar]Una consecuencia práctica de la redundancia es que muchos errores del código genético solo causen una mutación silenciosa o un error que no afectará a la proteína porque la hidrofilia o hidrofobia se mantiene por una sustitución equivalente de aminoácidos. Por ejemplo, un codón de NUN (N =cualquier nucleótido) tiende a codificar un aminoácido hidrófobo. NCN codifica residuos aminoacídicos que son pequeños en cuanto a tamaño y moderados en cuanto a hidropatía. NAN codifica un tamaño promedio de residuos hidrofílicos. UNN codifica residuos que no son hidrofílicos.[9][10] Estas tendencias pueden ser resultado de una relación del aminoacil ARNt sintetasas con los codones heredada un ancestro común de los seres vivos conocidos.
Incluso así, las mutaciones puntuales pueden causar la aparición de proteínas disfuncionales. Por ejemplo, un gen de hemoglobina mutado provoca la enfermedad de células falciformes. En la hemoglobina mutante un glutamato hidrofílico (Glu) se sustituye por una valina hidrofóbica (Val), es decir, GAA o GAG se convierte en GUA o GUG. La sustitución de glutamato por valina reduce la solubilidad de β-globina que provoca que la hemoglobina forme polímeros lineales unidos por interacciones hidrofóbicas entre los grupos de valina y causando la deformación falciforme de los eritrocitos. La enfermedad de las células falciformes no está causada generalmente por una mutación de nuevo. Más bien se selecciona en regiones de malaria (de forma parecida a la talasemia), ya que los individuos heterocigotos presentan cierta resistencia ante el parásito malárico Plasmodium (ventaja heterocigótica o heterosis).
La relación entre el ARNm y el ARNt a nivel de la tercera base se puede producir por bases modificadas en la primera base del anticodón del ARNt, y los pares de bases formados se llaman “pares de bases wobble” (tambaleantes). Las bases modificadas incluyen inosina y los pares de bases que no son del tipo Watson-Crick U-G.
Usos incorrectos del término
[editar]La expresión «código genético» se utiliza con mucha frecuencia en los medios de comunicación como sinónimo de genoma, de genotipo, o de ADN. Frases como «se analizó el código genético de los restos y coincidió con el de la desaparecida», o «se creará una base de datos con el código genético de todos los ciudadanos» son científicamente incorrectas. Es insensato, por ejemplo, aludir al «código genético de una determinada persona», porque el código genético es el mismo para todos los individuos.[11] Sin embargo, cada organismo tiene un genotipo propio, aunque es posible que lo comparta con otros si se ha originado por algún mecanismo de multiplicación asexual.
Tabla del código genético estándar
[editar]El código genético estándar se refleja en las siguientes tablas. La tabla 1 muestra qué aminoácido está codificado por cada uno de los 64 codones. La tabla 2 muestra qué codones especifican cada uno de los 20 aminoácidos que intervienen en la traducción. Estas tablas se llaman tablas de avance y retroceso respectivamente. Por ejemplo, el codón AAU es el aminoácido asparagina, y UGU y UGC representan cisteína (en la denominación estándar usando 3 letras, son los aminoácidos Asn y Cys, respectivamente).
| apolar | polar | básico | ácido | codón de parada |
| 2.ª base | |||||
|---|---|---|---|---|---|
| U | C | A | G | ||
| 1.ª base |
U | UUU (Phe/F) Fenilalanina UUC (Phe/F) Fenilalanina |
UCU (Ser/S) Serina UCC (Ser/S) Serina |
UAU (Tyr/Y) Tirosina UAC (Tyr/Y) Tirosina |
UGU (Cys/C) Cisteína UGC (Cys/C) Cisteína |
| UUA (Leu/L) Leucina | UCA (Ser/S) Serina | UAA Parada (Ocre) | UGA Parada (Ópalo) | ||
| UUG (Leu/L) Leucina | UCG (Ser/S) Serina | UAG Parada (Ámbar) | UGG (Trp/W) Triptófano | ||
| C | CUU (Leu/L) Leucina CUC (Leu/L) Leucina |
CCU (Pro/P) Prolina CCC (Pro/P) Prolina |
CAU (His/H) Histidina CAC (His/H) Histidina |
CGU (Arg/R) Arginina CGC (Arg/R) Arginina | |
| CUA (Leu/L) Leucina CUG (Leu/L) Leucina |
CCA (Pro/P) Prolina CCG (Pro/P) Prolina |
CAA (Gln/Q) Glutamina
CAG (Gln/Q) Glutamina |
CGA (Arg/R) Arginina CGG (Arg/R) Arginina | ||
| A | AUU (Ile/I) Isoleucina AUC (Ile/I) Isoleucina |
ACU (Thr/T) Treonina ACC (Thr/T) Treonina |
AAU (Asn/N) Asparagina AAC (Asn/N) Asparagina |
AGU (Ser/S) Serina AGC (Ser/S) Serina | |
| AUA (Ile/I) Isoleucina | ACA (Thr/T) Treonina | AAA (Lys/K) Lisina | AGA (Arg/R) Arginina | ||
| AUG (Met/M) Metionina, Comienzo | ACG (Thr/T) Treonina | AAG (Lys/K) Lisina | AGG (Arg/R) Arginina | ||
| G | GUU (Val/V) Valina GUC (Val/V) Valina |
GCU (Ala/A) Alanina GCC (Ala/A) Alanina |
GAU (Asp/D) Ácido aspártico GAC (Asp/D) Ácido aspártico |
GGU (Gly/G) Glicina GGC (Gly/G) Glicina | |
| GUA (Val/V) Valina GUG (Val/V) Valina |
GCA (Ala/A) Alanina GCG (Ala/A) Alanina |
GAA (Glu/E) Ácido glutámico GAG (Glu/E) Ácido glutámico |
GGA (Gly/G) Glicina GGG (Gly/G) Glicina | ||
Nótese que el codón AUG codifica la metionina, pero además sirve de sitio de iniciación; el primer AUG en un ARNm es la región que codifica el sitio donde se inicia la traducción de proteínas.
La siguiente tabla inversa indica qué codones codifican cada uno de los aminoácidos.
| Ala (A) | GCU, GCC, GCA, GCG | Lys (K) | AAA, AAG |
|---|---|---|---|
| Arg (R) | CGU, CGC, CGA, CGG, AGA, AGG | Met (M) | AUG |
| Asn (N) | AAU, AAC | Phe (F) | UUU, UUC |
| Asp (D) | GAU, GAC | Pro (P) | CCU, CCC, CCA, CCG |
| Cys (C) | UGU, UGC | Sec (U) | UGA |
| Gln (Q) | CAA, CAG | Ser (S) | UCU, UCC, UCA, UCG, AGU, AGC |
| Glu (E) | GAA, GAG | Thr (T) | ACU, ACC, ACA, ACG |
| Gly (G) | GGU, GGC, GGA, GGG | Trp (W) | UGG |
| His (H) | CAU, CAC | Tyr (Y) | UAU, UAC |
| Ile (I) | AUU, AUC, AUA | Val (V) | GUU, GUC, GUA, GUG |
| Leu (L) | UUA, UUG, CUU, CUC, CUA, CUG | ||
| Comienzo | AUG | Parada | UAG, UGA, UAA |
Aminoácidos 21 y 22
[editar]Existen otros dos aminoácidos codificados por el código genético en algunas circunstancias y en algunos organismos. Son la selenocisteína y la pirrolisina.
La selenocisteína (Sec, U)[12] es un aminoácido presente en multitud de enzimas (glutatión peroxidasas, tetraiodotironina 5' deiodinasas, tiorredoxina reductasas, formiato deshidrogenasas, glicina reductasas y algunas hidrogenasas). Está codificado por el codón UGA (que normalmente es de parada) cuando están presentes en la secuencia los elementos SecIS (secuencia de inserción de la selenocisteína).
El otro aminoácido, la pirrolisina (Pyl, O),[13][14] es un aminoácido presente en algunas enzimas de arqueas metanógenas. Está codificado por el codón UAG (que normalmente es de parada) cuando están presentes en la secuencia los elementos PylIS (secuencia de inserción de la pirrolisina).
Excepciones a la universalidad
[editar]Los códigos genéticos utilizados por todas las formas conocidas de vida son muy similares. No obstante existen algunas pequeñas variaciones. Como se mencionó con anterioridad, se conocen 22 códigos genéticos.[15] He aquí algunas diferencias con el estándar:
| Mitocondrias de vertebrados | AGA | Ter | * |
| AGG | Ter | * | |
| AUA | Met | M | |
| UGA | Trp | W | |
| Mitocondrias de invertebrados | AGA | Ser | S |
| AGG | Ser | S
AnGyBeL | |
| AUA | Met | M | |
| UGA | Trp | W | |
| AGG | Ausente en Drosophila | ||
| Mitocondrias de levaduras | AUA | Met | M |
| CUU | Thr | T | |
| CUC | Thr | T | |
| CUA | Thr | T | |
| CUG | Thr | T | |
| UGA | Trp | W | |
| CGA | Ausente | ||
| CGC | Ausente | ||
| Ciliados, Dasycladaceae y Hexamita (núcleo) | UAA | Gln | Q |
| UAG | Gln | Q | |
| Mitocondrias de mohos, protozoos y Coelenterate Mycoplasma y Spiroplasma (núcleo) |
UGA | Trp | W |
| Mitocondrias de equinodermos y platelmintos | AAA | Asn | N |
| AGA | Ser | S | |
| AGG | Ser | S | |
| UGA | Trp | W | |
| Euplotidae (núcleo) | UGA | Cys | C |
| Endomycetales (núcleo) | CUG | Ser | S |
| Mitocondrias de Ascidiacea | AGA | Gly | G |
| AGG | Gly | G | |
| AUA | Met | M | |
| UGA | Trp | W | |
| Mitocondrias de platelmintos (alternativo) | AAA | Asn | N |
| AGA | Ser | S | |
| AGG | Ser | S | |
| UAA | Tyr | Y | |
| UGA | W | ||
| Blepharisma (núcleo) | UAG | Gln | Q |
| Mitocondrias de Chlorophyceae | TAG | Leu | L |
| Mitocondrias de trematodos | TGA | Trp | W |
| ATA | Met | M | |
| AGA | Ser | S | |
| AGG | Ser | S | |
| AAA | Asn | N | |
| Mitocondrias de Scenedesmus obliquus | TCA | Ter | * |
| TAG | Leu | L | |
Origen del código genético
[editar]A pesar de las variaciones que existen, los códigos genéticos utilizados por todos los millones de formas conocidas de vida son muy similares. Esto sugiere que el código genético se estableció muy temprano en la historia de la vida y que tiene un origen común en todas las formas de vida actuales. El análisis filogenético sugiere que las moléculas ARNt evolucionaron antes que el conjunto actual de aminoacil-ARNt sintetasas.[16]
El código genético no es una asignación aleatoria de cada codon a cada aminoácido.[17] Por ejemplo, los aminoácidos que comparten la misma vía biosintética tienden a tener la primera base igual en sus codones,[18] y aminoácidos con propiedades físicas similares tienden a tener codones similares.[19][20]
Experimentos recientes demuestran que algunos aminoácidos tienen afinidad química selectiva por sus codones.[21] Esto sugiere que el complejo mecanismo actual de traducción del ARNm que implica la acción ARNt y enzimas asociadas puede ser un desarrollo posterior y que, en un principio, las proteínas se sintetizaran directamente sobre la secuencia de ARN, actuando este como ribozima y catalizando la formación de enlaces peptídicos (tal como ocurre con el ARNr 23S del ribosoma).
Se ha planteado la hipótesis de que el código genético estándar actual surgiera por expansión biosintética de un código más simple anterior. La vida primordial pudo adicionar nuevos aminoácidos (por ejemplo, subproductos del metabolismo), algunos de los cuales se incorporaron más tarde a la maquinaria de codificación genética. Se tienen pruebas, aunque circunstanciales, de que formas de vida primitivas empleaban un menor número de aminoácidos diferentes,[22] aunque no se sabe con exactitud qué aminoácidos y en qué orden entraron en el código genético.
Otro factor interesante a tener en cuenta es que la selección natural ha favorecido la adaptación del código para minimizar los efectos de errores.[23] Esto ha llevado a pensar que el código genético primitivo podría haber constado de codones de dos nucleótidos, lo que resulta bastante coherente con la hipótesis del balanceo del ARNt durante su acoplamiento (la tercera base no establece puentes de hidrógeno de Watson y Crick).
Véase también
[editar]Referencias
[editar]- ↑ a b «The Genetic Codes» (en inglés).
- ↑ Yılmaz, İrfan (6 de mayo de 2014). Evolución (en turco). Işık Yayıncılık Ticaret. ISBN 9789752787278. Consultado el 28 de noviembre de 2017.
- ↑ Aird, Kishori (2007-03). El ADN y la elección cuántica. Vesica Piscis. ISBN 9788493459789. Consultado el 28 de noviembre de 2017.
- ↑ Pumarola, A. (1987). Microbiología y parasitología médica. Elsevier España. ISBN 9788445800607. Consultado el 28 de noviembre de 2017.
- ↑ Ondarza, N. (1 de enero de 1994). Biología Molecular: Antes y después de la doble Hélice. Siglo XXI. ISBN 9682319064. Consultado el 28 de noviembre de 2017.
- ↑ Aldridge, Susan (julio de 1999). El hilo de la vida: De los genes a la ingeniería genética. Ediciones AKAL. ISBN 9788483230503. Consultado el 28 de noviembre de 2017.
- ↑ Orozco, Esther (1 de julio de 2011). Asi Estamos Hechos...Como Somos?: De la Lectura del Genoma a la Clonación Humana. Fondo de Cultura Económica. ISBN 9789681684426. Consultado el 28 de noviembre de 2017.
- ↑ Telenti, A., et al. (2016). «Deep sequencing of 10,000 human genomes». Proceedings of the National Academy of Sciences, 201613365.
- ↑ Yang et al. 1990. In Reaction Centers of Photosynthetic Bacteria. M.-E. Michel-Beyerle. (Ed.) (Springer-Verlag, Germany) 209-218
- ↑ Genetic Algorithms and Recursive Ensemble Mutagenesis in Protein Engineering «Copia archivada». Archivado desde el original el 15 de marzo de 2011. Consultado el 13 de enero de 2011.
- ↑ Viedma, Carlos de la Puente (2011). Fundamentos de Neurosociología. Editorial Complutense. ISBN 9788499381114. Consultado el 28 de noviembre de 2017.
- ↑ IUPAC-IUBMB Joint Commission on Biochemical Nomenclature (JCBN) and Nomenclature Committee of IUBMB (NC-IUBMB) (1999). «Newsletter 1999» (reprint, with permission). European Journal of Biochemistry 264 (2): 607-609. doi:10.1046/j.1432-1327.1999.news99.x. Archivado desde el original el 9 de junio de 2011. Consultado el 31 de mayo de 2009.
- ↑ John F. Atkins and Ray Gesteland (2002). The 22nd Amino Acid. Science 296 (5572): 1409–1410.
- ↑ Krzycki J (2005). The direct genetic encoding of pyrrolysine. Curr Opin Microbiol 8 (6): 706–712.
- ↑ «Las proteínas y el código genético Proteins and genetic code».
- ↑ De Pouplana, L.R.; Turner, R.J.; Steer, B.A.; Schimmel, P. (1998). «Genetic code origins: tRNAs older than their synthetases?». Proceedings of the National Academy of Sciences 95 (19): 11295. PMID 9736730. doi:10.1073/pnas.95.19.11295.
- ↑ Freeland SJ, Hurst LD (septiembre de 1998). «The genetic code is one in a million». J. Mol. Evol. 47 (3): 238-48. PMID 9732450. doi:10.1007/PL00006381. Archivado desde el original el 15 de septiembre de 2000.
- ↑ Taylor FJ, Coates D (1989). «The code within the codons». BioSystems 22 (3): 177-87. PMID 2650752. doi:10.1016/0303-2647(89)90059-2.
- ↑ Di Giulio M (octubre de 1989). «The extension reached by the minimization of the polarity distances during the evolution of the genetic code». J. Mol. Evol. 29 (4): 288-93. PMID 2514270. doi:10.1007/BF02103616.
- ↑ Wong JT (febrero de 1980). «Role of minimization of chemical distances between amino acids in the evolution of the genetic code». Proc. Natl. Acad. Sci. U.S.A. 77 (2): 1083-6. PMID 6928661. doi:10.1073/pnas.77.2.1083.
- ↑ Knight, R.D. and Landweber, L.F. (1998). Rhyme or reason: RNA-arginine interactions and the genetic code. Chemistry & Biology 5(9), R215-R220. PDF version of manuscript
- ↑ Brooks, Dawn J.; Fresco, Jacques R.; Lesk, Arthur M.; and Singh, Mona. (2002). Evolution of Amino Acid Frequencies in Proteins Over Deep Time: Inferred Order of Introduction of Amino Acids into the Genetic Code Archivado el 13 de diciembre de 2004 en Wayback Machine.. Molecular Biology and Evolution 19, 1645-1655.
- ↑ Freeland S.J.; Wu T. and Keulmann N. (2003) The Case for an Error Minimizing Genetic Code. Orig Life Evol Biosph. 33(4-5), 457-77.
Enlaces externos
[editar]Servicios en línea para convertir ADN (DNA, en inglés) en proteína:
Tablas del código genético
Revisiones
- Niremberg y Khorana, los hackers del ADN Archivado el 20 de agosto de 2011 en Wayback Machine., en el Museo Virtual Interactivo sobre la Genética y el ADN.
| Control de autoridades |
|
|---|
Datos: Q180618
Multimedia: Genetic code / Q180618