Inhibidor enzimàtic

Els inhibidors enzimàtics són molècules que uneixen els enzims i fan disminuir la seva activitat. Per mitjà del bloqueig de l'activitat dels enzims, es pot matar un patogen i corregir el desbalanç metabòlic; tal com fan moltes drogues, les quals són inhibidors enzimàtics. Aquests, també, han estat utilitzats com a herbicides i pesticides. No totes les molècules que uneixen als enzims són inhibidors; sinó que també hi ha activadors enzimàtics, molècules que s'uneixen als enzims i incrementen la seva activitat enzimàtica. La unió d'un inhibidor a un enzim pot fer aturar l'entrada del substrat al centre actiu o pot evitar la catalització de la reacció. La substància inhibidora pot ser reversible o irreversible. Els inhibidors irreversibles són aquells que, normalment, en reaccionar amb l'enzim el modifiquen químicament. Aquests inhibidors, per tant, modifiquen els residus dels aminoàcids que són necessaris per a l'activitat enzimàtica. En canvi, els inhibidors reversibles són aquells que s'uneixen de forma no covalent. No hi ha una sola classe d'inhibidors reversibles; sinó que dins aquest grup hi trobem subgrups segons si la inhibició és produïda per la unió dels inhibidors amb l'enzim, per la seva unió amb el complex substrat-enzim o per unir-se tant a un com a l'altre. Moltes drogues són inhibidors enzimàtics; per tant, el seu descobriment i millorament es troba en l'àrea activa de la recerca de la bioquímica i la farmacologia. Un inhibidor enzimàtic mèdic, és sempre avaluat per la seva especificitat (la seva mancança en la unió a altres proteïnes) i la seva potència (la constant de dissociació que indica la concentració necessària per inhibir l'enzim). Una alta especificitat i potència assegura que la droga tindrà pocs efectes secundaris i una baixa toxicitat. Els inhibidors enzimàtics també són formats a la natura i es troben implicats en el procés de regulació del metabolisme. Per exemple, enzims metabòlics poden ser inhibits per la concentració elevada de substrat. Aquesta classe de feedback negatiu disminueix el flux quan la quantitat de productes és elevada i manté l'homeòstasi a la cèl·lula. Altres inhibidors d'enzims cel·lulars són proteïnes que s'uneixen específicament a la diana de l'enzim. Aquests, poden ajudar en el control d'enzims, com les proteases o nucleases, per tal d'evitar malmetre la cèl·lula. Un bon exemple és l'inhibidor de la ribonucleasa, que uneix a les ribonucleases per mitjà d'interaccions estretes entre proteïnes.[1] Els inhibidors naturals d'enzims, poden ser verinosos i usats com a defensa contra depredadors o com una via per matar presses.
Inhibidors reversibles
[modifica]Tipus d'inhibidors reversibles
[modifica]
.
Els inhibidors reversibles s'uneixen a enzims de forma no covalent com són els ponts d'hidrogen, les interaccions hidròfobes i les unions iòniques. Molts dels ponts dèbils entre l'inhibidor i el centre actiu, es combinen per formar ponts forts i específics. En contrast amb els substrats d'inhibidors irreversibles, els reversibles generalment no se sotmeten a reaccions, ja que la seva unió pot ser fàcilment trencada per dilució o diàlisis. Hi ha quatre classes reversibles d'inhibidors enzimàtics. Aquests han estat classificats segons l'efecte que produeixen en variar la concentració de substrat.[2]
- En la inhibició compatible, el substrat i la partícula inhibidora no poden unir-se al mateix temps; tal com mostra la figura de l'esquerra. Això normalment succeeix perquè l'inhibidor presenta afinitat per la part de l'enzim on també s'uneix el substrat; d'aquesta manera, es produeix una competició entre el substrat i l'inhibidor per unir-se a l'enzim. Per aquesta raó, aquesta classe d'inhibició pot ser evitada afegint una concentració elevada de substrat; ja que llavors la probabilitat que l'enzim s'uneixi amb el substrat augmenta. La majoria dels enzims competitius tenen una estructura semblant a la del substrat real (vegeu els exemples següents).
- En la inhibició incompetitiva, la unió amb el complex enzim-substrat, no hauria de ser confosa amb l'anterior. En aquest cas, la velocitat màxima (Vmàx) i les unions eficients (Km), ambdós, disminueixen.
- En la inhibició mixta, l'inhibidor pot unir-se a l'enzim al mateix temps que l'enzim amb el substrat. Tot i així, la unió amb l'inhibidor afecta la unió amb el substrat i viceversa. La inhibició pot ser reduïda però no per mitjà de l'addició de substrat. Encara que és possible que l'inhibidor s'uneixi al centre actiu, la inhibició normalment resulta d'un efecte al·lostèric on l'inhibidor s'uneix a diferents parts de l'enzim. La unió de l'inhibidor modifica la conformació de l'enzim (estructura terciària o la seva forma tridimensional); la qual cosa, redueix l'afinitat del substrat pel centre actiu.
- La inhibició no competitiva es forma per la inhibició mesclada on la unió de l'inhibidor amb l'enzim redueix la seva activitat però no afecta la unió amb el substrat. Com a resultat, la inhibició depèn solament de la concentració de l'inhibidor.[3]
Quantitativa descripció dels inhibidors reversibles
[modifica]Els inhibidors reversibles poden ser descrits quantitativament en termes. L'inhibidor s'uneix a l'enzim i al complex enzim-substrat i modifica les constants cinètiques de l'enzim. En la cinètica de Michaelis-Menten; un enzim (E) s'uneix al substrat (S) per formar el complex enzim-substrat (ES). Després de la catàlisi, el complex es trenca per formar producte (P) i l'enzim queda lliure. L'inhibidor (I) pot unir-se a l'enzim o al complex enzim-substrat per les constants de dissociació Ki i Ki', respectivament.
- Els inhibidors competitius que es poden unir a l'enzim però no al complex enzim-substrat. La Km (concentració de substrat quan la velocitat de la reacció és ½ de la velocitat màxima) augmenta; però la velocitat màxima no es veu afectada (l'inhibidor no modifica la catàlisi perquè no es pot unir al complex enzim-substrat).
- Els inhibidors no competitius tenen igual afinitat per l'enzim que pel complex enzim-substrat (Ki=Ki'). Per tant, no modifiquen la Km però disminueixen la velocitat màxima.
- Els inhibidors mixtos, s'uneixen a l'enzim i al complex enzim-substrat; però l'afinitat per aquestes dues formes de l'enzim són diferents (Ki ≠ Ki'). Aquests inhibidors interfereixen en la unió amb el substrat incrementant Km i en la catàlisi del complex enzim-substrat disminuint la velocitat màxima.
Quan un enzim té múltiples substrats, els inhibidors poden mostrar diferents classes d'inhibicions segons el substrat considerat. El resultat, és que el centre actiu té dos llocs d'unió diferents. Per exemple, un inhibidor respecte al seu substrat A s'uneix a la primera unió; però l'inhibidor no competitiu, s'uneix a l'altre lloc d'unió del centre actiu.
Mesurant les constants de dissociació de l'inhibidor reversible
[modifica]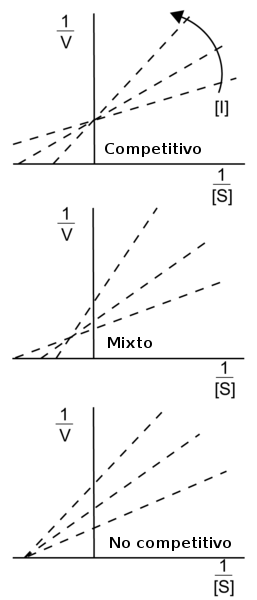
Tal com s'ha indicat prèviament, un inhibidor enzimàtic és caracteritzat per dues constants de dissociació, Ki i Ki', de l'enzim i del complex enzim-substrat respectivament. Els inhibidors enzimàtics poden ser mesurats directament; un mètode extremadament acurat és el calomeria isotèrmica de titulació, on l'inhibidor és tractat amb una solució on es troba l'enzim i amb calor; a partir de la qual és calculada la que és absorbida o alliberada.[4] Tot i així, l'altra constant de dissociació Ki' és difícil de calcular de forma directa, ja que la durada de vida del complex enzim-substrat és molt curta; ja que es tracta d'un pas entremig que té la finalitat de formar producte. Per aquest motiu Ki' és, normalment, mesurada de manera indirecta per mitjà de l'observació de l'activitat de l'enzim en diferents substrats i amb concentracions diferents d'inhibidors;[5] i seguint la modificació de la cinètica de Michaelis-Menten equació de Michaelis-Menten:
on modificant els factors α i α', que són definides per la concentració d'inhibidor, es poden determinar a partir de:
Per això, en presència d'un inhibidor, l'efectivitat de l'enzim que era Km i Vmàx, es converteixen en (α/α')Km i (1/α')Vmàx, respectivament. En canvi, modificant l'equació de Michaelis Menten s'assumeix que la unió de l'inhibidor a l'enzim ha arribat a l'equilibri; el qual disminuirà el procés d'inhibició que tinguin un sub-nanomolar de constants de dissociació. En aquest cas, és més pràctic tractar com si fos una unió forta i estreta entre l'inhibidor i el substrat; és a dir, com si es tractés d'un inhibidor irreversible (vegeu a sota); en canvi, és encara possible estimar Ki' si Ki és mesurada independentment. Aquests efectes sobre la cinètica enzimàtica que es poden produir segons les diferents classes d'enzims es poden representar gràficament per mitjà de la representació gràfica amb els diagrames de Lineaweaver-Burk i Eadie-Hofstee. Per exemple, en la recta de Lineaweaver-Burk de la dreta, la inhibició competitiva talla l'eix de les y, il·lustrant que no es veu afectada la velocitat màxima. De forma semblant, la recta talla l'eix de les x, mostrant que aquesta classe d'inhibidors tampoc modifiquen la Km. Així i tot, pot resultar difícil estimar la Ki i la Ki' adequadament a partir d'aquests diagrames;[6] per tant, és recomanable estimar-les a partir d'un dels mètodes no-lineals de regressió que seran més fiables, tal com s'ha indicat anteriorment.
Casos especials
[modifica]- El mecanisme d'inhibició parcialment competitiva és semblant al dels no competitius, excepte pel fet que el complex enzim-substrat té una activitat catalítica; la qual pot ser més elevada o més baixa (enzim parcialment competitiu) respecte altres complexos enzim-substrat. Aquesta inhibició tendeix a disminuir la velocitat màxima, però no afecta el valor de Km.[7]
- La inhibició incompetitiva succeeix quan l'inhibidor s'uneix només al complex enzim-substrat; el qual, és catalíticament inactiu. Aquest mètode d'inhibició és poc comuna i causa la disminució de Km i de la Vmàx.[7]
- La inhibició a partir de la quantitat de producte i substrat és aquella on el producte o el substrat reaccionen amb l'enzim per tal d'inhibir o activar una reacció. Aquesta reacció pot seguir l'estructura de les inhibicions competitives, les incompetitives o les mixtes. En la inhibició de formació de substrat hi ha una disminució de l'activitat de l'enzim a causa de l'alta concentració d'aquest. Això demostra l'existència de dos llocs d'unió que té l'enzim amb el substrat. A baixa concentració de substrat, l'alta afinitat del centre actiu és ocupada i se segueixen les normals constants cinètiques. En canvi, a altes concentracions d'aquest, el segon lloc d'inhibició també esdevé ocupat; inhibint l'enzim.[8] La inhibició de producte serveix normalment per regular el metabolisme que s'anomena feedback negatiu .
- Inhibició per poques unions estretes succeeix quan l'inicial complex enzim-inhibidor (EI) és isomeritzat amb una unió més forma d'EI*, però el procés d'inhibició és reversible. Això manifesta que té una inhibició lenta sobre l'enzim. Sota aquestes condicions, la tradicional constant cinètica de Michaelis Menten aporta un fals valor de Ki; el qual és dependent del temps. El verdader valor de Ki es pot obtenir a partir de l'anàlisi de més complexos que encenen (kon) o apaguen (koff) els valors de les constants per l'associació de l'inhibidor. Observeu l'inhibidor irreversible de sota per a més informació.
Exemples
[modifica]Com els enzims s'uneixen al substrat estretament i la majoria dels inhibidors reversibles s'uneixen al centre actiu dels enzims, no sorprèn que alguns d'aquests inhibidors tinguin una estructura molt semblant a l'estructura del substrat. Un exemple d'aquesta semblança són els inhibidors de proteases, una classe molt efectiva de droga antiretroviral usada per al tractament de HIV.[9] L'estructura del ritonavir, un inhibidor de proteases està basada en un pèptid i conté tres enllaços peptídics, tal com es mostra en la imatge de la dreta. Per tant, com que té una estructura semblant a la del substrat competeix amb aquest per unir-se al centre actiu. Els inhibidors enzimàtics són normalment dissenyats per imitar la transició d'un estat o com a intermediari d'un enzim en catalitzar una reacció. Això assegura que l'efecte de l'inhibidor afecta la inhibició en el període estacionari fent disminuir l'afinitat (disminuint Ki). Un exemple d'aquesta inhibició de la transició del període d'inhibició és la droga antiviral oseltamivir; la qual imita l'estructura de l'anell de l'ió oxoni en les reaccions de l'enzim viral neuraminidasa.[10]

Però, no tots els inhibidors estan basats en les estructures del substrat. Un exemple és l'estructura d'un altre inhibidor de proteases anomenat tipranavir (es pot observar a l'esquerra). Aquesta molècula no està basada en un pèptid i no s'assembla a la proteïna del substrat. Aquests no pèptids inhibidors poden ser més estables que els inhibidors que contenen unions peptídiques perquè no seran substrats per a les proteases i seran menys degradats.[11]
En el disseny d'una droga és important considerar la concentració de substrat que és exposada en l'enzim. Per exemple, alguns inhibidors de proteïna-cinases tenen estructures semblants a l'adenosina trifosfat; un dels substrats dels enzims. Tot i així, les drogues que són simples inhibidors competitius hauran de competir amb altes concentracions d'ATP de la cèl·lula. Les proteïnes cinases poden ser també inhibides per mitjà de la competició en els punts d'unió on les quinases interaccionen amb el substrat, ja que moltes d'aquestes es troben en concentracions més baixes que l'ATP. Com a conseqüència, si dos inhibidors de proteïnes quinasa s'uneixen a un centre actiu sobre el qual tenen una afinitat semblant, però sols han de competir amb l'ATP, llavors l'inhibidor competitiu inhibirà més eficaçment en els llocs d'unió.[12]
Inhibidors irreversibles
[modifica]
Els inhibidors irreversibles, normalment, modifiquen un enzim covalentment, per la qual cosa la inhibició no pot ser invertida. Els inhibidors irreversibles acostumen a contenir grups funcionals reactius com aldehids, grups nitrogenats, haloalcans o alquens. Aquests grups electrofílics reaccionen amb les cadenes d'aminoàcids per formar unions covalents. Els residus modificats són aquells que contenen en les seves cadenes laterals nucleòfiles com, per exemple, un grup hidroxil o un grup sulfhidril. Això inclou als aminoàcids serina, cisteïna, treonina o tirosina.[13]
Tipus d'inhibicions irreversibles
[modifica]La inhibició irreversible és diferent de la inactivació enzimàtica reversible. Els inhibidors irreversibles són generalment específics per un tipus d'enzim i no inactiven a totes les proteïnes. No funcionen destruint l'estructura proteica, sinó alterant específicament l'estructura tridimensional del centre actiu, inhabilitant-lo. Per exemple, el pH i les temperatures extremes causen la desnaturalització de quasi totes les proteïnes, però aquest no és un efecte específic. De manera similar, alguns tractaments químics no específics destrueixen l'estructura de la proteïna: per exemple, si són sotmeses a una elevada concentració d'àcid clorhídric, el qual hidrolitzarà els enllaços peptídics que mantenen units els aminoàcids de les proteïnes.[14] Els inhibidors irreversibles donen lloc a una inhibició depenent del temps i, per això, la seva potència no pot ser caracteritzada mitjançant la determinació del valor IC50. Això és degut al fet que la quantitat d'enzim actiu a una concentració donada d'inhibidor irreversible serà diferent depenent del temps de pre-incubació de l'inhibidor amb l'enzim. Per això, en lloc del valor IC50 s'utilitza el paràmetre kobs/[I],[15] on kobs és el primer valor observat de la taxa d'inactivació (obtingut en representar en una gràfica log%activitat VS. temps) i [I] és la concentració d'inhibidor. El paràmetre kobs/[I] és vàlid sempre que l'inhibidor no es trobi a concentracions saturants (cas en el qual tindríem que kobs=kinact).
Anàlisi de la inhibició irreversible
[modifica]
Els inhibidors irreversibles formen inicialment un complex reversible i no covalent amb l'enzim (EI o ESI), que reaccionarà posteriorment per produir una modificació covalent per donar el que es denomina el “complex del punt mort” EI*. La taxa a la qual es forma EI* s'anomena taxa d'inactivació o Kinact. Atès que la formació d'EI pot competir amb ES, la unió dels inhibidors irreversibles pot ser previnguda per competència, tant amb el substrat com amb un segon inhibidor irreversible. Aquest efecte de protecció és una bona evidència d'una reacció específica de l'inhibidor irreversible amb el centre actiu. Els passos d'unió i inactivació d'aquesta reacció són analitzats incubant l'enzim amb l'inhibidor i mesurant l'activitat que va quedant al llarg del temps. L'activitat anirà disminuint amb el temps, generalment seguint una dinàmica de decreixement exponencial. Ajustant aquestes dades a una equació de rang podem obtenir la taxa d'inactivació a una determinada concentració d'inhibidor. Això es fa a diferents concentracions d'inhibidor. Si un complex EI reversible està involucrat, el rang d'inactivació serà saturable i en ajustar-ho a la corba obtindrem els valors de Kinact i Ki.[16]
Un altre mètode que és àmpliament utilitzat en aquestes anàlisis és l'espectrometria de masses. En aquest cas, la mesura exacta de la massa de l'enzim natiu sense modificar i de l'enzim inactiu ens la dona l'augment de massa causat per la reacció amb l'inhibidor, amb la qual cosa obtindrem l'estequiometria de la reacció.[17] Per dur a terme aquesta tècnica sol ser necessari l'ús d'un espectròmetre de masses MALDI-TOF. Una tècnica complementària a aquesta és l'empremta peptídica, que implica la digestió de proteïnes natives o modificades amb una peptidasa com la tripsina. Això genera una sèrie de pèptids que poden ser analitzats utilitzant un espectròmetre de masses. El pèptid que canviï la seva massa després de dur a terme una reacció amb l'inhibidor serà aquell que contingui el centre de la modificació.
Casos especials
[modifica]
Pirodoxal-5'fosfat (Py) i l'enzim (E) no es mostren. Adaptat de[18]
No tots els inhibidors irreversibles formen unions covalents amb l'enzim que tenen com a diana. Alguns inhibidors reversibles s'uneixen tan fortament al seu objectiu que són pràcticament irreversibles. Aquests inhibidors d'unió forta solen mostrar una cinètica semblant a la dels inhibidors irreversibles que formen enllaços covalents. En aquests casos, alguns d'aquests inhibidors s'uneixen ràpidament a l'enzim formant un complex EI de baixa afinitat, que després experimenta una reacció més lenta cap a un complex EI* molt fortament unit. Aquest comportament cinètic es denomina unió lenta.[19]
Aquest lent reajust posterior normalment implica un canvi conformacional quan l'enzim es plega al voltant de la molècula inhibidora. Com a exemples d'inhibidors d'unió lenta és necessari destacar algun fàrmac important com el metotrexat,[20] el allopurinol[21] i la forma activa de l'aciclovir.[22]
Exemples d'inhibidors irreversibles
[modifica]
El diisopropilfluorofosfat (DFP) és un d'inhibidor irreversible de la proteasa. L'enzim hidrolitza l'enllaç entre el fòsfor i el fluor, però el residu de fosfat es manté unit a una serina en el centre actiu, inactivant-lo.[23] A més, el DFP també reacciona amb el centre actiu de l'acetilcolinesterasa en la sinapsi de les neurones, la qual cosa el converteix en una potent neurotoxina, amb una dosi letal a partir de quantitats inferiors a 100mg.[24]
La inhibició suïcida és un tipus comú d'inhibició irreversible, on l'enzim converteix l'inhibidor en una substància reactiva en el seu centre actiu. Un exemple d'això és l'inhibidor de biosintetitzadors de poliaminas α-difluorometillornitina o DFMO, que és un anàleg de l'aminoàcid ornitina, i que s'utilitza per tractar la Tripanosomiasis Africana (la malaltia del son). L'ornitina descarboxilasa pot catalitzar la descarboxilació del DFMO substituint a l'ornitina. Tot i així, aquesta reacció de descarboxilació és seguida per l'eliminació de l'àtom de fluor, la qual cosa converteix a aquest intermediari en una imina, una espècie altament electrofílica. Aquesta forma reactiva del DFMO reacciona posteriorment amb un residu de cisteïna o lisina en el centre actiu per inactivar l'enzim irreversiblement.[18]
Atès que la inhibició irreversible implica tot sovint la formació inicial d'un complex. EI no covalent, a vegades és possible que un inhibidor pugui unir-se a un enzim de diverses maneres. Com a exemple cal destacar el cas de l'enzim tripanotió reductasa, que pertany al protozou i paràsit humà Trypanosoma cruzi, on dues molècules d'un inhibidor anomenat mostassa quinacrina presenten la capacitat d'unir-se al seu centre actiu. Hi ha una molècula que s'uneix de forma reversible, però l'altra s'uneix de forma covalent en reaccionar amb un residu d'aminoàcid a través del seu grup mostassa nitrogenada.[25]
Descobriment i disseny d'inhibidors
[modifica]
La investigació i desenvolupament de nous fàrmacs és un llarg procés en el qual el primer pas és, gairebé sempre, el descobriment d'un nou inhibidor enzimàtic. En el passat, l'única forma de descobrir aquests nous inhibidors era mitjançant el sistema de prova i error: provant un elevat nombre de compostos contra l'enzim al qual es volia inhibir i esperant aconseguir que algun fos efectiu. Aquesta aproximació encara segueix aconseguint actualment un gran èxit gràcies a nous sistemes com la química combinatòria, que ràpidament produeix un gran nombre de compostos nous, i a la tecnologia basada en la investigació de processament d'alt rendiment, mitjançant la qual es poden revisar ràpidament aquestes enormes biblioteques químiques d'inhibidors útils.[26]
Més recentment, s'ha començat a aplicar un nou tipus d'investigació alternativa: el disseny racional de fàrmacs que utilitzen l'estructura tridimensional del centre actiu d'un enzim per predir quines molècules podrien ser inhibidores.[27] Una vegada realitzada la predicció, es proven i se selecciona el millor. Aquest nou inhibidor després es fa servir per intentar obtenir una estructura de l'enzim en un complex enzim-substrat per mostrar com s'uneix la molècula al centre actiu. Aquesta estructura és després inspeccionada i es duen a terme certs canvis en l'estructura de l'inhibidor a fi d'optimitzar la unió amb l'enzim. Aquest cicle de prova i optimització es repeteix de forma successiva fins a obtenir un inhibidor suficientment potent; és a dir, quan s'aconsegueix que el valor de la constant de dissociació estigui al voltant de <10-9 M.[28]
Aplicacions dels inhibidors
[modifica]Els inhibidors enzimàtics poden trobar-se en la natura, però també són dissenyats i produïts com a part de la farmacologia i la bioquímica. Els verins naturals són sovint inhibidors enzimàtics que han evolucionat per defensar a una planta o un animal contra els seus depredadors. Aquestes toxines naturals inclouen alguns dels compostos més verinosos coneguts fins ara. Els inhibidors artificials són tot sovint emprats com a medicaments, però també existeixen alguns utilitzats com insecticides (com el malathion), herbicides (com el glifosfat) o desinfectants (com el triclosà).
Quimioterapia
[modifica]
Els inhibidors enzimàtics són utilitzats principalment com a fàrmacs en el tractament de diverses malalties. Molts d'aquests inhibidors són capaços d'actuar sobre enzims humans per així corregir determinades patologies. Això no obstant, no tots els fàrmacs són inhibidors enzimàtics. Alguns d'ells, tals com els fàrmacs anti-epilèptics, alteren l'activitat enzimàtica de manera indirecta, augmentant o disminuint la síntesi de tal enzim. Aquests efectes són denominats inducció i inhibició enzimàtica i consisteixen en alteracions en el patró d'expressió gènica, la qual cosa no està relacionada amb el tipus d'inhibició enzimàtica descrita. Altres drogues interaccionen amb altres dianes cel·lulars que no són enzims, com els canals iònics o els receptors de membrana.

Un exemple d'inhibidor enzimàtic terapèutic és el sildenafil, utilitzat com a tractament de la disfunció erèctil. Aquest compost és un potent inhibidor de la fosfodiesterasa tipus 5 específic de GMPc, un enzim que degrada una molècula de senyalització cel·lular, el GMPc. Aquesta molècula de senyalització és la responsable de la relaxació del múscul llis i permet el flux de sang cap a l'interior dels cossos cavernosos del penis, la qual cosa provoca l'erecció. El sildenafil inhibeix l'activitat de l'enzim que degrada el senyal, el GMPc, unint-se al mateix lloc que aquest a causa de la seva similitud estructural. Això permet que el GMPc.[29] no sigui degradat i pugui romandre actiu durant períodes més llargs de temps.
Un altre exemple de similitud estructural entre inhibidors i substrats enzimàtics és el que es dona entre el metotrexat, un fàrmac, i l'àcid fòlic, un coenzim. L'àcid fòlic és la forma oxidada del substrat de la dihidrofolat reductasa, un enzim implicat en la biosíntesi de timidina, purines i aminoàcids. El metotrexat és un potent inhibidor d'aquest enzim que, en estar relacionat amb la síntesi de nucleòtids, presenta una toxicitat específica d'aquelles cèl·lules amb una ràpida taxa de creixement. Per això, el metotrexat s'utilitza sovint com a fàrmac anticancerós en quimioteràpia[30]
Una altra classe d'inhibidors enzimàtics són utilitzats amb el fi d'inhibir aquells enzims necessaris per a la supervivència de patògens. Per exemple, els bacteris presenten una gruixuda paret cel·lular composta principalment d'un polímer, que és un peptidoglicà.[31]
Certs antibiòtics com la penicil·lina i la vancomicina inhibeixen a l'enzim responsable de la producció i encreuament de les fibres de peptidoglicans, la qual cosa dona lloc a una pèrdua de força de la paret cel·lular i, en conseqüència, a la lisi de la cèl·lula, incapaç llavors de resistir l'elevada pressió osmòtica. També s'utilitzen altres tipus de toxicitats selectives mitjançant antibiòtics que aprofiten les diferències presents en l'estructura dels ribosomes o en la síntesi d'àcids grassos. El disseny de fàrmacs es veu molt facilitat en aquells casos en els quals l'enzim diana és essencial per a la supervivència del patogen i no està present o és molt diferent en humans.
Control metabòlic
[modifica]Els inhibidors enzimàtics també són importants pel que fa a control metabòlic. Moltes de les rutes metabòliques que tenen lloc a la cèl·lula són inhibides per metabòlits que controlen l'activitat enzimàtica mitjançant processos de regulació al·lostèrica o inhibició per substrat. Com a exemple cal destacar la regulació al·lostèrica de la glucòlisi. Aquesta ruta catabòlica consumeix glucosa i produeix ATP, NADH i piruvat. Una de les etapes clau en la regulació de la glucòlisi és la reacció catalitzada per la fosfofructoquinasa-1 (PFK1). Quan els nivells d'ATP augmenten, l'ATP s'uneix al centre al·lostèric de la PFK1, amb el que es redueix l'activitat de l'enzim, la glucòlisi s'inhibeix i els nivells d'ATP es redueixen de nou. Aquest control per retroalimentació negativa (feedback negatiu) ajuda a mantenir els nivells d'ATP constants en la cèl·lula. Tot i així, les rutes metabòliques no estan únicament regulades per mitjà de la inhibició; l'activació enzimàtica és igualment important. L'enzim PFK1 presenta activadors al·lostèrics, com són la fructosa 2,6-bifosfat i el ADP.[32]
La inhibició enzimàtica fisiològica també pot ser produïda per inhibidors proteics específics. Aquest mecanisme es pot observar en el pàncrees, on se sintetitzen multitud de precursors d'enzims digestius denominats zimògens. Molts d'ells són activats per la tripsina, una proteasa, per la qual cosa és molt important inhibir l'activitat de la tripsina en el pàncrees amb el fi de prevenir fenòmens d'autodigestió. Un dels mecanismes per mantenir la tripsina inactiva és la producció d'un potent i específic inhibidor de tripsina en el pàncrees. Aquest inhibidor s'uneix amb una alta afinitat a la tripsina, prevenint així la seva activitat[33] Tot i que l'inhibidor de tripsina és una proteïna, no és hidrolitzat per la mateixa tripsina, ja que elimina les molècules d'aigua del seu centre actiu i desestabilitza l'estat de transició.[34] Altres exemples d'inhibidors enzimàtics fisiològics són l'inhibidor barstar, la funció del qual és inhibir l'activitat d'una ribonucleasa bacteriana anomenada barnasa,[35] i els inhibidors de les protein-fosfatases.[36]
Inhibidors de l'acetilcolinesterasa
[modifica]
L'acetilcolinesterase (AChE) és un enzim que es troba en els animals, des dels insectes fins als humans. És essencial en l'activitat que duen a terme les neurones, ja que la seva funció consisteix en la ruptura del neurotransmissor acetilcolina en els seus constituents, acetat i colina. És un cas únic entre els neurotransmissors, ja que la majoria d'ells, com la serotonina, la dopamina o la noradrenalina, són reabsorbits en l'espai sinàptic. Existeix un ampli nombre d'inhibidors de la AChE que són utilitzats tant en el camp de la medicina com en el camp de l'agricultura. Alguns d'aquests inhibidors són reversibles, com la fisostigmina i la neostigmina, els quals són utilitzats en el tractament de la miastenia gravis i en l'aplicació d'anestèsia. Els pesticides del tipus carbamat són un altre exemple d'inhibidors reversibles de la AChE. Com a exemple d'inhibidors irreversibles de la AChE, calen destacar els insecticides organofosforats, com el malathion, el paratió i els clorpirifos.
Verins naturals
[modifica]Tant alguns animals com algunes plantes són capaços de sintetitzar una àmplia gamma de substàncies verinoses de divers origen: metabòlits[37] secundaris, pèptids i proteïnes, que poden actuar com inhibidors enzimàtics. Les toxines naturals solen ser petites molècules orgàniques amb tanta diversitat que probablement existeixen inhibidors per la majoria de processos metabòlics. Aquestes inhibicions poden produir-se a diferents nivells de la cèl·lula: inhibició de receptors de membrana, de canals iònics o de proteïnes estructurals. Per exemple, el paclitaxel (taxol), una molècula orgànica obtinguda del Taxus, s'uneix amb una elevada afinitat als dímers de tubulina, impedint així el procés de polimerització dels microtúbuls; crucial, entre altres coses, en la divisió cel·lular.[38]
Molts d'aquests verins naturals actuen com neurotoxines capaces de causar paràlisi que poden conduir a la mort, podent ser utilitzades com a estratègia defensiva davant de depredadors o com a sistema de caça en la captura de presses.[39] Tot i les seves característiques tòxiques, alguns d'aquests verins naturals són molt valorats pels seus potencials usos terapèutics quan són administrats en dosis adequades. Com a exemple de neurotoxina a destacar cal anomenar els glicoalcaloides, obtinguts a partir de les plantes que pertanyen a la família Solanaceae (entre les quals s'inclouen la patata, el tomàquet i l'albergínia), que són inhibidors de l'acetilcolinesterasa. La inhibició d'aquest enzim causa un augment descontrolat dels nivells del neurotransmissor acetilcolina, la qual cosa comporta la paràlisi muscular i la mort. La neurotoxicitat també pot ser el resultat de la inhibició de receptors, com en el cas de l'atropina, obtinguda a partir de l'espècie Atropa belladonna, que funciona com un antagonista competitiu dels receptors muscarínics de l'acetilcolina.[40]
Tot i que moltes de les toxines naturals són metabòlits secundaris, algunes són pèptids o proteïnes. Com a exemple, cal destacar la toxina peptídica α-amanitina, trobada en els fongs de l'espècie Amanita phalloides, que és un potent inhibidor enzimàtic capaç d'impedir l'activitat de l'ARN polimerasa II en la transcripció de l'ADN.[41]
Una altra toxina peptídica és la microcistina, trobada en certes algues i capaç d'inhibir certes protein-fosfatases.[42] Aquesta toxina pot contaminar les reserves d'aigua si les algues que la produeixen arriben a determinats nivells de concentració. S'ha demostrat el seu alt potencial carcinogènic i la seva capacitat de causar hemorràgia hepàtica aguda i mort després d'una exposició a altes dosis.[43] Les proteïnes també poden arribar a ser verins naturals, com és el cas de l'inhibidor de la tripsina, que es troba en alguns llegums (explicat anteriorment). Menys comuns són els enzims tòxics. Aquest tipus d'enzims actuen com a inhibidors irreversibles de dianes enzimàtiques que modifiquen químicament els seus substrats enzimàtics. Un exemple és el ricí, una planta que té una proteïna tòxica extremadament potent. Aquest enzim és una glicosidasa, que inactiva els ribosomes. Com la proteïna del ricí és un inhibidor catalític irreversible, això permet que tan sols una molècula de la proteïna pugui matar una cèl·lula[44]
Referències
[modifica]- ↑ Shapiro R, Vallee BL. Interaction of human placental ribonuclease with placental ribonuclease inhibitor. Biochemistry. 1991 Feb 26;30(8):2246–55. PMID: 1998683
- ↑ Berg J., Tymoczko J. and Stryer L. (2002) Biochemistry. W. H. Freeman and Company ISBN 0-7167-4955-6
- ↑ Irwin H. Segel, Enzyme Kinetics : Behavior and Analysis of Rapid Equilibrium and Steady-State Enzyme Systems. Wiley–Interscience; New edition (1993), ISBN 0-471-30309-7
- ↑ Holdgate GA. Making cool drugs hot: isothermal titration calorimetry as a tool to study binding energetics. Biotechniques. 2001 Jul;31(1):164–6 PMID: 11464510
- ↑ Leatherbarrow RJ. Using linear and non-linear regression to fit biochemical data. Trends Biochem Sci. 1990 Dec;15(12):455–8. PMID: 2077683
- ↑ Tseng SJ, Hsu JP. A comparison of the parameter estimating procedures for the Michaelis–Menten model. J. Theor. Biol. 1990 Aug 23;145(4):457–64. PMID: 2246896
- ↑ 7,0 7,1 Irwin H. Segel, Enzyme Kinetics : Behavior and Analysis of Rapid Equilibrium and Steady-State Enzyme Systems. Wiley-Interscience; New Ed edition (1993), ISBN 0-471-30309-7
- ↑ Dixon, M. Webb, E.C., Thorne, C.J.R. and Tipton K.F., Enzymes (3rd edition) Longman, London (1979) See p. 126
- ↑ Hsu JT, Wang HC, Chen GW, Shih SR. Antiviral drug discovery targeting to viral proteases. Curr Pharm Des. 2006; 12(11):1301–14. PMID: 16611117
- ↑ Lew W, Chen X, Kim CU (2000). "Discovery and development of GS 4104 (oseltamivir): an orally active influenza neuraminidase inhibitor". Curr. Med. Chem. 7 (6): 663–72. PMID: 10702632
- ↑ Fischer PM (2003). "The design, synthesis and application of stereochemical and directional peptide isomers: a critical review". Curr. Protein Pept. Sci. 4 (5): 339–56. doi:10.2174/1389203033487054. PMID: 14529528
- ↑ Bogoyevitch MA, Barr RK, Ketterman AJ. Peptide inhibitors of protein kinases—discovery, characterisation and use. Biochim. Biophys. Acta. 2005 Dec 30;1754(1–2):79–99. PMID: 16182621
- ↑ Lundblad R. L. Chemical Reagents for Protein Modification CRC Press Inc (2004) ISBN 0-8493-1983-8
- ↑ N. Price, B. Hames, D. Rickwood (Ed.) Proteins LabFax Academic Press (1996) ISBN 0-12-564710-7
- ↑ Adam GC, Cravatt BF, Sorensen EJ. (2001) Profiling the specific reactivity of the proteome with non-directed activity-based probes. Chem. Biol. 8(1):81-95.
- ↑ Maurer T, Fung HL. Comparison of Methods for Analyzing Kinetic Data From Mechanism-Based Enzyme Inactivation: Application to Nitric Oxide Synthase. AAPS PharmSci. (2000) 2(1)E8. PMID: 11741224
- ↑ Loo JA, DeJohn DE, Du P, Stevenson TI, Ogorzalek Loo RR (1999). "Application of mass spectrometry for target identification and characterization". Med Res Rev 19 (4): 307–19. doi:10.1002/(SICI)1098-1128(199907)19:4<307::AID-MED4>3.0.CO;2-2. PMID: 10398927.
- ↑ 18,0 18,1 ^ a b Poulin R, Lu L, Ackermann B, Bey P, Pegg AE. Mechanism of the irreversible inactivation of mouse ornithine decarboxylase by alpha-difluoromethylornithine. Characterization of sequences at the inhibitor and coenzyme binding sites. J Biol Chem. 1992 Jan 5;267(1):150–8. PMID: 1730582
- ↑ Szedlacsek, S.E. and Duggleby, R.G. Kinetics of slow and tight-binding inhibitors. Meth. Enzymol., (1995) 249: 144–180. PMID: 7791610
- ↑ Stone SR, Morrison JF. Mechanism of inhibition of dihydrofolate reductases from bacterial and vertebrate sources by various classes of folate analogues. Biochim Biophys Acta. 1986 Feb 14;869(3):275–85. PMID: 3511964
- ↑ Hille R, Massey V. Tight binding inhibitors of xanthine oxidase. Pharmacol Ther. 1981;14(2):249–63. PMID: 4322209
- ↑ Reardon JE. Herpes simplex virus type 1 and human DNA polymerase interactions with 2'-deoxyguanosine 5'-triphosphate analogues. Kinetics of incorporation into DNA and induction of inhibition. J Biol Chem. 1989 Nov 15;264(32):19039–44. PMID: 2553730
- ↑ J. A. Cohen, R. A. Oosterbaan and F. Berends Organophosphorus compounds Meth. Enzymol. (1967) 11, 686
- ↑ Brenner, G. M. (2000): Pharmacology. Philadelphia, PA: W.B. Saunders Company. ISBN 0-7216-7757-6
- ↑ Saravanamuthu A, Vickers TJ, Bond CS, Peterson MR, Hunter WN, Fairlamb AH. Two interacting binding sites for quinacrine derivatives in the active site of trypanothione reductase: a template for drug design. J Biol Chem. 2004 Jul 9;279(28):29493–500. PMID: 15102853
- ↑ Koppitz M, Eis K (2006). "Automated medicinal chemistry". Drug Discov. Today 11 (11-12): 561–8. doi:10.1016/j.drudis.2006.04.005. PMID: 16713909.
- ↑ Scapin G (2006). "Structural biology and drug discovery". Curr. Pharm. Des. 12 (17): 2087–97. doi:10.2174/138161206777585201. PMID: 16796557.
- ↑ Hunter WN. Rational drug design: a multidisciplinary approach. Mol Med Today. 1995 Apr;1(1):31, 34. PMID: 9415135
- ↑ Maggi M, Filippi S, Ledda F, Magini A, Forti G. Erectile dysfunction: from biochemical pharmacology to advances in medical therapy. Eur J Endocrinol. 2000 Aug;143(2):143–54 PMID: 10913932
- ↑ McGuire JJ. Anticancer antifolates: current status and future directions. Curr Pharm Des. 2003;9(31):2593–613. PMID: 14529544
- ↑ Katz AH, Caufield CE. Structure-based design approaches to cell wall biosynthesis inhibitors. Curr Pharm Des. 2003;9(11):857–66. PMID: 12678870
- ↑ Okar DA, Lange AJ. Fructose-2,6-bisphosphate and control of carbohydrate metabolism in eukaryotes. Biofactors. 1999;10(1):1–14.
- ↑ Nicholas Price, Lewis Stevens, Fundamentals of Enzymology, Oxford University Press, (1999) ISBN 0-19-850229-X
- ↑ Smyth TP. Substrate variants versus transition state analogues as noncovalent reversible enzyme inhibitors. Bioorg Med Chem. 2004 Aug 1;12(15):4081–8. PMID: 15246086
- ↑ Hartley RW. Barnase and barstar: two small proteins to fold and fit together. Trends Biochem Sci. 1989 Nov;14(11):450–4. PMID: 2696173
- ↑ Oliver CJ, Shenolikar S. Physiologic importance of protein phosphatase inhibitors. Front Biosci. 1998 Sep 1;3:D961–72. PMID: 9727084
- ↑ Tan G, Gyllenhaal C, Soejarto DD. Biodiversity as a source of anticancer drugs. Curr Drug Targets. 2006 Mar;7(3):265-77. PMID: 16515527
- ↑ Abal M, Andreu JM, Barasoain I. Taxanes: microtubule and centrosome targets, and cell cycle dependent mechanisms of action. Curr Cancer Drug Targets. 2003 Jun;3(3):193–203. PMID: 12769688
- ↑ Hostettmann K, Borloz A, Urbain A, Marston A, Natural Product Inhibitors of Acetylcholinesterase Current Organic Chemistry, 2006 May;10(8):825–47
- ↑ DeFrates LJ, Hoehns JD, Sakornbut EL, Glascock DG, Tew AR. Antimuscarinic intoxication resulting from the ingestion of moonflower seeds. Ann Pharmacother. 2005 Jan;39(1):173-6. PMID: 15572604
- ↑ Vetter J. Toxins of Amanita phalloides. Toxicon. 1998 Jan;36(1):13–24. PMID: 9604278
- ↑ Holmes CF, Maynes JT, Perreault KR, Dawson JF, James MN. Molecular enzymology underlying regulation of protein phosphatase-1 by natural toxins. Curr Med Chem. 2002 Nov;9(22):1981–9. PMID: 12369866
- ↑ Bischoff K. The toxicology of microcystin-LR: occurrence, toxicokinetics, toxicodynamics, diagnosis and treatment. Vet Hum Toxicol. 2001 Oct;43(5):294-7. PMID: 11577938
- ↑ Hartley MR, Lord JM. Cytotoxic ribosome-inactivating lectins from plants. Biochim Biophys Acta. 2004 Sep 1;1701(1–2):1–14. PMID: 15450171