서식지의 단편화
Habitat fragmentation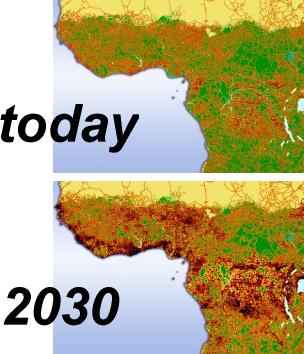
서식지 조각화는 유기체가 선호하는 환경(습지)에서 개체군 조각화 및 생태계 붕괴를 야기하는 불연속(조각화)의 출현을 설명한다.서식지 조각화의 원인은 물리적 환경의 배치를[1] 서서히 바꾸는 지질학적 과정(분화의 주요[1] 원인 중 하나로 추정됨)과 환경을 훨씬 더 빠르게 변화시키고 많은 종의 멸종을 야기하는 토지 전환과 같은 인간 활동을 포함한다.좀 더 구체적으로 말하면, 서식지 조각화는 크고 인접한 서식지가 작고 고립된 서식지로 [2][3]나뉘는 과정이다.


정의.
서식지의 단편화라는 용어는 5가지 개별 현상을 포함한다.
- 서식지의 총 면적 감소
- 내부 감소: 가장자리 비율
- 하나의 서식지 조각이 다른 서식지 영역으로부터 격리됨
- 하나의 서식지를 여러 개의 작은 패치로 분할
- 각 서식지의 평균 크기 감소
"응축...서식지의 양을 감소시킬 뿐만 아니라 작은 고립된 패치를 만들어냄으로써 나머지 서식지의 특성도 변화시킨다." (van den Berg et al.[failed verification] 2001)서식지의 단편화는 현상의 경관 수준과 패치 수준 프로세스입니다.즉, 패치 영역, 가장자리 효과 및 패치 모양의 [4]복잡성에 대해 설명합니다.
과학 문헌에서는, 「습지 단편화」라는 용어가 서식지의 손실의 경우에 적용되는가, 또는 주로 서식지의 면적이 큰폭으로 감소하지 않고, 서식지가 작은 조각으로 잘라지는 현상에 적용되는가 하는 논란이 있다."습지 단편화"라는[3] 보다 엄격한 정의를 사용하는 과학자들은 서식지의 상실을 "습지 손실"이라고 부르고 서식지의 연관성이 낮아지고 전체적인 서식지가 적은 상황을 묘사한다면 두 용어 모두를 명시적으로 언급할 것이다.
게다가, 서식지 조각화는 생물 다양성에 대한 침습적 위협으로 여겨지는데, 생물 다양성이 생물학적 침입, 과도한 개발, [5]또는 오염보다 많은 종들에게 영향을 미치기 때문이다.
또한, 서식지 조각화의 영향은 토종 식물과 같은 종들이 변화하는 환경에 효과적으로 적응할 수 있는 능력을 손상시킨다.궁극적으로, 이것은 한 세대에서 다음 세대로, 특히 더 작은 개체 크기에서 사는 종들의 유전자 흐름을 막는다.반면에, 더 큰 개체군의 종들은 발생할 수 있는 더 많은 유전자 돌연변이를 가지고 있고 그러한 환경에서 종의 생존을 증가시킬 수 있는 유전자 재조합의 영향을 가지고 있다.전반적으로, 서식지 조각화는 서식지 붕괴와 서식지 손실을 초래하고, 둘 다 생물 다양성의 파괴와 관련이 있다.
원인들
자연 원인
화산 활동, 화재, 기후 변화와 같은 자연적인 과정을 통한 서식지 파괴의 증거가 화석 [1][failed verification]기록에서 발견됩니다.예를 들어, 3억 년 전 유러아메리카 열대 우림의 서식지가 파괴되면서 양서류의 다양성이 크게 상실되었지만, 동시에 건조한 기후는 [1]파충류들 사이의 다양성의 폭발을 촉진시켰다.
인적 원인
서식지의 분할은 농업, 농촌 개발, 도시화, 수력 발전 저수지 조성 등의 인간 활동을 위해 자생식물이 제거될 때 사람에 의해 자주 발생한다.한때 연속되었던 서식지는 분리된 조각으로 나뉘었다.집중적인 개간 후, 분리된 조각들은 경작지, 목초지, 포장도로 또는 심지어 척박한 땅에 의해 서로 격리된 매우 작은 섬들이 되는 경향이 있다.후자는 종종 열대림에서 자르고 태우는 농사의 결과이다.호주 뉴사우스웨일스 주 중서부의 밀밭에서는 자생식물의 90%가 개간되고 북미의 높은 풀밭의 99% 이상이 개간되어 서식지가 극단적으로 파괴되고 있다.
내인성 vs 외인성
서식지의 분열을 초래할 수 있는 두 가지 유형의 과정이 있습니다.외생적인 과정과 내생적인 과정이 있다.내생은 종 생물학의 한 부분으로 발달하는 과정이기 때문에 그것들은 전형적으로 생물, 행동, 그리고 종 내 또는 종 간의 상호작용의 변화를 포함한다.내인성 위협은 번식 패턴 또는 마이그레이션 패턴을 변경할 수 있으며, 대부분의 경우 외인성 프로세스에 의해 트리거됩니다.외인성 과정은 종 생물학과 무관하며 서식지 감소, 서식지 세분화 또는 서식지 고립을 포함할 수 있다.이러한 과정은 종들의 행동을 근본적으로 변화시킴으로써 내생적 과정에 상당한 영향을 미칠 수 있다.서식지의 세분화 또는 고립은 계절적 이동의 변화를 포함하여 종의 분산이나 이동을 초래할 수 있다.이러한 변화는 종의 밀도의 감소, 경쟁의 증가 또는 [6]포식 증가로 이어질 수 있습니다.
시사점
서식지 및 생물 다양성 손실
서식지 조각화가 생물 다양성에 영향을 미치는 주요 방법 중 하나는 유기체가 사용할 수 있는 적절한 서식지의 양을 줄이는 것이다.서식지 조각화는 종종 서식지 파괴와 이전에 지속되었던 [7]서식지의 세분화를 수반한다.식물과 다른 생물들은 [8]서식지의 변화하는 공간적 구성에 빠르게 반응할 수 없기 때문에 어떤 형태의 서식지 조각화에 의해 불균형적으로 영향을 받는다.
서식지 파괴 과정에서 발생할 수 있는 서식지 손실은 [9]종에게 가장 큰 위협으로 여겨진다.그러나, 경관 내 서식지의 양(파편화 그 자체라고[3] 함)의 영향과는 무관하게, 경관 내 서식지 패치의 구성 효과는 [10]작다고 제안되었다.경험적 연구를 검토한 결과, 과학 문헌의 종 발생, 풍부성 또는 다양성에 대한 서식지 단편화의 유의한 영향 381개 중 76%는 긍정적이고 24%는 [11]부정적이었다.이러한 결과에도 불구하고, 과학 문헌은 긍정적인 [12]효과보다 부정적인 효과를 강조하는 경향이 있다.서식지 조각화의 긍정적인 효과는 여러 개의 작은 서식지가 동등한 [11]크기의 하나의 큰 부분보다 더 높은 보존 가치를 가질 수 있다는 것을 의미한다.따라서 토지 공유 전략은 토지 절약 [11]전략보다 생물종에 더 긍정적인 영향을 미칠 수 있다.서식지 손실의 부정적인 영향은 일반적으로 서식지 조각화보다 훨씬 더 큰 것으로 보이지만, 두 사건은 밀접하게 연관되어 있고 관찰은 보통 [13]서로 독립적이지 않다.

영역은 단편[14] 내 종 수를 결정하는 주요 요소이며, 전지구적 개체수 멸종 위험에 대한 인구통계학적 및 유전학적 과정의 상대적 기여는 서식지 구성, 확률적 환경 변화 및 종의 [15]특성에 따라 달라진다.기후, 자원 또는 기타 요인의 경미한 변동은 현저하지 않고 많은 인구에서 신속하게 수정될 수 있으며, 소규모 고립된 인구에서는 치명적일 수 있다.그러므로 서식지의 파편화는 종 [14]멸종의 중요한 원인이다.세분화된 모집단의 인구 역학은 비동기적으로 변화하는 경향이 있다.세분화되지 않은 풍경에서 인구 감소는 인구 증가 근처의 이민에 의해 "회복"될 수 있다.파편화된 풍경에서는 파편 사이의 거리가 이러한 현상을 막을 수 있습니다.또한, 어떤 장벽에 의해 이민의 원천으로부터 분리된 거주지의 점유되지 않은 조각들은 인접한 조각들보다 다시 채워질 가능성이 낮다.콜롬비아 반점 개구리와 같은 작은 종들도 구조 효과에 의존하고 있다.연구에 따르면 청소년의 25%가 200m 이상 거리를 이동하는 반면 성인의 4%는 200m 이상 거리를 이동하는 것으로 나타났다.이 중 95%가 새 지역에 남아 있어 [16]생존을 위해 이 여정이 필요하다는 것을 보여준다.
또한, 서식지 조각화는 가장자리 효과로 이어진다.빛, 온도 및 바람의 미세 기후 변화는 조각 주변과 [17]조각의 내부 및 외부 부분의 생태를 바꿀 수 있습니다.습도가 떨어지고 기온과 바람의 수준이 높아짐에 따라 이 지역에서 화재가 발생할 가능성이 높아진다.외래종과 해충종은 그러한 교란된 환경에서 쉽게 자리를 잡을 수 있고, 가축의 근접성은 종종 자연 생태계를 해친다.또한, 조각의 가장자리를 따라 서식하는 서식지는 기후가 다르고 내륙 서식지와는 다른 종을 선호한다.따라서 작은 조각은 내부 서식지를 필요로 하는 종에게 불리하다.인접 서식지의 보존 비율은 유전적, 종적 생물다양성 보존과 밀접한 관련이 있다.일반적으로 10%의 잔류 인접 서식지는 50%의 생물다양성 [18]손실을 가져온다.
많은 제3세계 국가에서 남아 있는 육지 야생동물 서식지의 대부분은 서식지 상실을 방해하는 도로와 같은 도시 확장의 개발을 통해 분열을 경험했다.수생 생물들의 서식지는 댐과 물의 [19]다양성으로 인해 산산조각이 났다.이러한 서식지의 조각들은 짝과 먹이를 찾을 수 있는 넓은 영역을 필요로 하는 종들을 지탱하기에 충분히 크거나 연결되어 있지 않을 수 있다.서식지의 손실과 분열은 철새 종들이 휴식처를 찾고 이동 [19]경로를 따라 먹이를 먹는 것을 어렵게 만든다.
정보에 근거한 보존
서식지 조각은 종종 종이 위협받거나 [20]멸종위기에 처하게 되는 원인이다.생존 가능한 서식지의 존재는 모든 종의 생존에 매우 중요하며, 많은 경우에, 남아있는 서식지의 조각화는 보존 생물학자들에게 어려운 결정으로 이어질 수 있습니다.보존할 수 있는 자원의 양이 한정되어 있는 경우, 기존의 고립된 서식지를 보호하거나 가능한 한 가장 큰 인접지를 얻기 위해 토지를 환매하는 것이 바람직하다.드문 경우지만, 보존에 의존하는 종은 고립된 서식지에 분포됨으로써 질병 보호의 어느 정도 척도를 얻을 수 있습니다, 그리고 전체적인 서식지의 손실을 통제했을 때 몇몇 연구는 종의 풍부함과 조각화 사이의 긍정적인 관계를 보여주었습니다; 이 현상은 유효하지만 서식지의 양 가설이라고 불립니다.이 주장에 대해 [10][21]논란이 되고 있다.보존과 가장 관련이 있는 조각의 크기에 대한 지속적인 논쟁은 종종 SLOSS(Single Large or Mouth Small)라고 불린다.생물 다양성 핫스팟에서의 서식지 상실은 국지적인 멸종 위기를 초래할 수 있으며, 일반적으로 핫스팟에서의 서식지 상실은 멸종위기와 멸종된 고유종들의 [22]수를 나타내는 좋은 지표가 될 수 있다.
서식지 단편화 문제에 대한 한 가지 해결책은 토종 식물의 통로를 보존하거나 심음으로써 파편들을 연결하는 것이다.경우에 따라서는 브리지 또는 언더패스가 2개의 [23]fragment를 결합하기에 충분할 수 있습니다.이것은 고립 문제를 완화할 수 있지만 내부 서식지의 손실은 줄일 수 없다.야생동물 통로는 식량원이나 다른 천연자원이 그들의 핵심 서식지에 부족할 때 동물들이 이동하거나 새로운 지역을 차지하도록 도울 수 있고, 동물들은 유전적 다양성을 증가시키기 위해 이웃 지역에서 새로운 짝을 찾을 수 있다.계절적으로 이주하는 종은 인간의 발달 장벽을 방해하지 않을 때 더 안전하고 효과적으로 이주할 수 있다.
도시 경관의 지속적인 확장으로 인해, 현재의 연구는 녹색 지붕이 서식지 회랑의 가능한 매개체가 될 수 있다고 보고 있다.최근의 연구는 녹색 지붕이 절지동물, 특히 벌과 요충의 [24]서식지를 연결하는 데 도움이 된다는 것을 발견했다.
또 다른 완화 조치는 내부 서식지의 양을 늘리기 위해 작은 잔해를 확대하는 것이다.개발 토지는 종종 더 비싸고 복원하는 데 상당한 시간과 노력이 필요할 수 있기 때문에 이것은 비현실적일 수 있습니다.
최적의 솔루션은 일반적으로 고려 중인 특정 종 또는 생태계에 따라 달라집니다.대부분의 새들처럼, 더 많은 이동성 종들은 연결된 서식지를 필요로 하지 않는 반면, 설치류와 같은 몇몇 작은 동물들은 탁 트인 땅에서 포식자에 더 많이 노출될 수 있습니다.이 질문들은 일반적으로 메타포메이션 섬 생물 지리학의 제목에 포함된다.
유전적 위험
나머지 서식지가 더 작기 때문에, 그들은 더 적은 [25]종의 작은 개체군을 부양하는 경향이 있다.작은 개체군은 그들의 [26]장기 생존에 영향을 미치는 다양한 유전적 결과들의 위험을 증가시킨다.잔류 개체군은 종종 이전에 지속적 서식지에서 발견된 유전적 다양성의 하위 집합만을 포함한다.이러한 경우에, 적응과 같이 근본적인 유전적 다양성에 작용하는 과정들은 환경 변화에 직면하여 살아남기 위해 체력을 유지하는 대립 유전자들의 더 작은 풀을 가지고 있다.그러나 유전적 다양성의 하위 집합이 여러 서식지의 단편들 사이에 분할되는 일부 시나리오에서는 각 개별 단편들이 [27]다양성의 하위 집합을 나타내더라도 거의 모든 원래의 유전적 다양성이 유지될 수 있다.
유전자 흐름과 근친교배
유전자 흐름은 같은 종의 개체들이 번식을 통해 유전 정보를 교환할 때 발생한다.개체군은 이주를 통해 유전적 다양성을 유지할 수 있다.서식지가 조각나고 면적이 줄어들면, 유전자 흐름과 이동은 전형적으로 감소한다.더 적은 수의 개체들이 나머지 조각으로 이동하며, 한때 하나의 큰 개체군의 일부였을 수도 있는 연결되지 않은 작은 개체군은 생식적으로 고립될 것이다.분열로 인해 유전자 흐름이 감소한다는 과학적 증거는 연구 종에 따라 다르다.장기 수분과 분산 메커니즘을 가진 나무들은 [28]분열에 따른 유전자 흐름 감소를 경험하지 못할 수 있지만, 대부분의 종은 서식지 [8]분열에 따른 유전자 흐름 감소의 위험에 처해 있다.
유전자 흐름의 감소와 생식적 격리는 관련된 개체 간의 근친 교배를 초래할 수 있다.근친 교배가 항상 부정적인 건강 결과를 초래하는 것은 아니지만, 근친 교배가 건강 감소와 관련이 있을 때 근친교배 우울증이라고 불립니다.동종 교배는 적합성을 감소시키는 유해 대립 유전자의 발현을 촉진하면서 호모 접합의 수준이 증가함에 따라 더욱 우려의 대상이 된다.서식지 조각화는 유전자 [29][30]흐름의 감소로 인해 많은 종의 근친교배 우울증을 초래할 수 있다.근친교배 우울증은 국지적 [31]멸종과 같은 보존 위험과 관련이 있다.
유전적 표류
작은 집단은 유전적 표류에 더 민감하다.유전적 표류는 모집단의 유전자 구성에 대한 무작위적인 변화이며 유전적 다양성의 감소를 이끈다.인구가 적을수록 유전적 표류는 자연선택보다는 진화의 원동력이 될 가능성이 높다.유전자 표류는 무작위 과정이기 때문에, 그것은 종이 환경에 더 적응하는 것을 허락하지 않는다.서식지 조각화는 작은 [29]개체군의 유전적 다양성에 부정적인 영향을 미칠 수 있는 유전적 표류 증가와 관련이 있다.하지만, 연구는 일부 나무 종들이 개체 수가 10마리 이하로 [27]작을 때까지 유전적 표류의 부정적인 결과에 탄력적일 수 있다는 것을 암시한다.
식물군에 대한 서식지 조각화의 유전적 영향
서식지 조각화는 크기를 줄이고 식물군의 공간적 고립을 증가시킨다.무작위 유전자 이동의 영향 증가로 인한 유전자 변형과 집단 간 유전자 발산 방법 증가로 근친 교배가 증가하고 식물 종 내 유전자 흐름이 감소한다.유전적 변이는 잔류 인구 크기에 따라 감소할 수 있지만, 모든 단편화 이벤트가 유전적 손실과 다른 유형의 유전적 변이로 이어지는 것은 아니다.드물게, 단편화는 또한 지역 유전자 [32]구조를 분해하면서 잔여 개체들 사이의 유전자 흐름을 증가시킬 수 있다.
적응.
개체군이 자연 도태에 반응하여 진화하기 위해서는 자연 도태가 유전적 표류보다 더 강한 진화력이 될 수 있을 만큼 충분히 커야 한다.일부 식물 종에서 서식지의 단편화가 적응에 미치는 영향에 대한 최근 연구는 단편화된 환경의 유기체가 [33][34]단편화에 적응할 수 있다는 것을 시사했다.그러나, 작은 모집단 [35]크기 때문에 단편화가 적응 능력을 감소시키는 경우도 많다.
영향을 받는 종의 예
서식지의 단편화로 인해 유전적 영향을 받은 종은 다음과 같다.

- 맥쿼리아오스트랄라시카[29][36]
- 파구스 실바티카[37]
- 베툴라나나[27]
- 리넬라오르나타[38]
- 오초토나프린셉스[39]
- 우타 스탄스부리아나[40]
- 플레스티오돈스킬토니아누스[40]
- 후두엽충[40]
- 카메아파시아타[40]
동물 행동에 미치는 영향
비록 서식지 조각화가 유전학과 종의 멸종률에 영향을 미치는 방식이 많이 연구되어 왔지만, 조각화는 또한 종의 행동과 문화에도 영향을 미치는 것으로 나타났다.사회적 상호작용이 종의 적합성과 생존을 결정하고 영향을 미치기 때문에 이것은 중요하다.서식지 조각화는 이용 가능한 자원을 변화시키고, 그 결과 서식지의 구조를 변화시키고, 종의 행동과 다른 종들 간의 역학 관계를 변화시킨다.영향을 받는 행동은 번식, 짝짓기, 먹이찾기, 종의 분산, 의사소통 및 이동 패턴과 같은 종 내일 수도 있고 포식자-사료 [41]관계와 같은 종 간의 행동일 수도 있다.게다가, 동물들이 조각난 숲이나 풍경 사이에 있는 미지의 지역으로 모험할 때, 그들은 아마도 인간과 접촉할 수 있고, 이것은 그들을 큰 위험에 처하게 하고 그들의 [5]생존 가능성을 더 감소시킨다.
포식 행동
인위적인 활동으로 인한 서식지 조각화는 종의 수와 그 종의 [41]구성원을 변경함으로써 많은 종의 포식자-사료 역학에 큰 영향을 미치는 것으로 나타났다.이것은 주어진 집단에 있는 동물들 사이의 자연적인 포식자-사료 관계에 영향을 미치고 그들이 그들의 행동과 상호작용을 바꾸도록 강요하며, 따라서 소위 "행동 우주 경쟁"[42]을 재설정합니다.단편화가 이러한 상호작용을 변화시키고 재형성하는 방법은 여러 가지 형태로 발생할 수 있습니다.대부분의 먹잇감 종들은 그들의 포식자들로부터 피난처인 땅을 가지고 있어서, 그들이 새끼를 번식하고 키울 수 있게 해준다.도로와 파이프라인과 같은 인간이 도입한 구조물은 이러한 피난처에서 포식자 활동을 촉진하여 포식자-사료 [42]중첩을 증가시킴으로써 이러한 영역을 변화시킨다.먹이에게 유리한 경우에도 그 반대가 발생할 수 있으며, 먹이 피신처를 증가시키고 포식율을 감소시킨다.또한 조각화는 포식자의 풍부함이나 포식자의 효율성을 증가시켜 포식율을 증가시킨다.[42]포식자의 식단이 얼마나 다양한지, 포식자와 [41]먹잇감의 서식 조건이 얼마나 유연한지를 포함하여, 포식자-사료 역학이 특정 종에 영향을 미치는 정도를 증가시키거나 감소시킬 수 있는 몇 가지 다른 요인들도 있다.어떤 종이 영향을 받고 이러한 다른 요인들에 따라, 포식자-사료 역학에 대한 조각화와 그 영향은 종 [41]멸종의 원인이 될 수 있습니다.이러한 새로운 환경 압력에 대응하여 새로운 적응 행동이 개발될 수 있습니다.먹이 종은 짝짓기 전략을 변경하거나 먹이 및 [41]사료와 관련된 행동과 활동을 변경하는 등의 전략을 통해 포식 위험 증가에 적응할 수 있다.
북방산림순록
브리티시컬럼비아의 한대 삼림 순록에서는 파편화의 영향이 나타난다.종 보호 구역은 도로와 [43]파이프라인과 같은 선형적 특징에 의해 방해된 이탄지 습지입니다.이러한 특징들은 그들의 천적, 늑대, 그리고 흑곰이 더 효율적으로 풍경과 [43]땅 사이를 이동할 수 있게 해주었다.포식자들이 순록의 피난처에 더 쉽게 접근할 수 있기 때문에, 이 종의 암컷들은 이 지역을 피하려고 노력하며, 그들의 번식 행동과 태어난 [43]자손들에게 영향을 미칩니다.
커뮤니케이션 행동
새의 의사소통 행동에 영향을 미치는 단편화는 듀퐁의 종달새에서 잘 연구되어 왔다.종달새는 주로 스페인의 지역에 서식하며,[43] 종 사이의 문화적 전달 수단으로 노래를 사용하는 작은 참새목 새이다.종달새들은 노래와 영토 호출이라는 두 가지 뚜렷한 목소리를 가지고 있다.영역 호출은 수컷이 다른 수컷 종달새로부터 영역을 방어하고 신호를 보내는 데 사용되며 수컷이 라이벌 [44]노래에 응답할 때 이웃 영역 간에 공유됩니다.때때로 그것은 [45]영토에 대한 임박한 공격을 나타내는 위협 신호로 사용됩니다.노래 레퍼토리가 크면 수컷이 다른 수컷으로부터 자신의 영역을 지키는 능력이 커지기 때문에 수컷의 생존과 번식 능력을 높일 수 있고 수컷의 수가 많을수록 더 다양한 노래가 [44]전달된다는 것을 의미한다.농업, 임업 및 도시화로 인한 듀퐁 종달새 지역의 단편화는 이들의 통신 [45]구조에 큰 영향을 미치는 것으로 보인다.수컷은 특정 거리의 영역만을 라이벌로 인식하기 때문에, 분할로 인해 영역을 고립시키는 것은 수컷이 더 이상 그것을 사용할 이유나 그에 [45]걸맞는 노래가 없기 때문에 영역 호출의 감소로 이어집니다.
인간은 또한 생태계에 다양한 영향을 미쳐 동물의 행동과 [46]생성된 반응에 영향을 끼쳤다.비록 펄프나 제지업을 위해 숲에서 나무를 베는 것과 같이 이러한 가혹한 조건에서 살아남을 수 있는 종들이 있지만, 이러한 변화에서 살아남을 수 있는 종들은 있지만, 그렇지 못한 종들도 있다.예를 들어 다양한 수생곤충은 편광에 의해 알을 낳기 위한 적절한 연못을 식별할 수 있지만,[47] 사람에 의한 생태계 변경에 의해 예를 들어 건조한 아스팔트 건조도로에 의해 유도되는 인공광선을 방출하는 인공구조물로 유도된다.
미생물에 미치는 영향
서식지 조각화는 종종 많은 식물과 동물의 개체수와 생물 다양성에 미치는 영향과 관련이 있지만, 생태계의 상호 연결성 때문에 환경의 미생물군에 미치는 영향도 크다.분열의 증가는 부패의 원인이 되는 균류의 개체수 감소와 다양성 그리고 그들이 [48]숙주로 있는 곤충들과 관련이 있다.이것은 오래된 [49]생육림에 비해 고도로 단편화된 지역의 단순화된 먹이사슬과 관련이 있다.또한 가장자리 효과는 빛 가용성, 바람의 존재, 강수량 변화 및 [50]잎 쓰레기의 전체적인 수분 함량 변화로 인해 내부 숲에 비해 상당히 다양한 미세 환경을 초래하는 것으로 나타났다.이러한 미세 환경은 종종 전반적인 [48]산림 건강에 도움이 되지 않습니다. 왜냐하면 이러한 환경은 특정 환경에 의존하는 전문가의 희생을 감수하면서 일반 종들의 번식을 가능하게 하기 때문입니다.
상호적이고 적대적인 관계에 미치는 영향
메타데이터 분석에 따르면 서식지 조각화는 포식이나 초식 등 적대적인 관계에 덜 영향을 [51]미치면서 상호주의적 관계에 큰 영향을 미치는 것으로 나타났다.예를 들어, 메소인 휘장과 메가칠레 사이의 상호주의적 관계입니다.한 연구에 따르면 파편화되지 않은 숲에서 더 많은 수분과 M. 휘장의 과일 생산량이 증가하여 파편화된 [52]숲이 나타났다.둥지 포식 대립관계의 예에 대해서는 단편화된 숲에 대한 둥지 포식 증가가 없다는 연구결과가 나와 가장자리 효과 [53]가설을 뒷받침하지 못하고 있다.
포레스트 플래그멘테이션
숲 조각은 숲이 (자연적 또는 인공적으로) 숲 조각이나 숲 [1]잔재라고 알려진 비교적 작고 고립된 숲으로 축소되는 서식지 조각의 한 형태이다.나머지 삼림지대를 구분하는 간섭 매트릭스는 자연 개방 지역, 농지 또는 개발 지역일 수 있습니다.섬 생물지리학의 원리에 따라 남은 삼림지는 목초지, 들판지, 구획지, 상가 등의 바다에서 숲의 섬처럼 기능한다.이 조각들은 생태계의 붕괴 과정을 거치기 시작할 것이다.
숲의 단편화에는 ROW(유틸리티 Right-of-Ways)와 같이 덜 미묘한 형태의 불연속성도 포함된다.유틸리티 ROW는 미국의 [54]500만 에이커에 달하는 지역에 걸쳐 많은 산림 커뮤니티에 퍼지고 있기 때문에 생태학적 관심이 높다.유틸리티 ROW에는 송전 ROW, 가스관 및 통신 ROW가 포함됩니다.송전 ROW는 송전선로의 식생방해를 방지하기 위해 제작되었습니다.일부 연구에 따르면 복도 및 그 주변의 미세 기후 변화로 인해 송전 ROW가 인접한 삼림 [55]지역보다 더 많은 식물 종을 보유하고 있는 것으로 나타났다.선로권리와 관련된 산림지역의 단종은 선로권이 초기 연속 단계에서 보존되기 때문에 토종 벌과 초원 종의 [56]생물다양성 피난처 역할을 할 수 있다.
숲의 파편화는 동물들의 식량 자원과 서식지를 감소시켜 이 종들을 갈라놓는다.따라서, 이러한 동물들을 포식 효과에 훨씬 더 민감하게 만들고 이종 교배를 덜 하게 만드는 것은 유전적 다양성을 낮춘다.[57]
시사점
숲의 파편화는 숲, 특히 [58]열대지방의 생물다양성에 대한 가장 큰 위협 중 하나이다.애초에 분열을 초래한 서식지 파괴 문제는 다음과 같이 복잡해진다.
- 특히 큰 척추동물의 생존 가능한 집단을 지탱할 수 없는 개별 숲 조각
- 생존 가능한 집단을 지탱할 수 있는 적어도 하나의 파편을 가지고 있지 않은 종의 국지적 멸종
- 조각의 외부 영역의 조건을 변화시켜 진정한 숲 내부 [59]서식지의 양을 크게 감소시키는 가장자리 효과.
숲 패치의 동식물군에 대한 단편화의 영향은 a) 패치의 크기 및 b) [60]분리 정도에 따라 달라진다.분리는 가장 가까운 유사 패치까지의 거리 및 주변 영역과의 대비에 따라 달라집니다.예를 들어, 개간된 지역을 다시 삼림으로 만들거나 재생을 허용하면, 식생의 구조적 다양성이 증가하여 숲 파편의 고립이 감소한다.그러나 이전에 임야였던 땅이 목초지, 농경지 또는 사람이 거주하는 개발 지역으로 영구적으로 전환될 경우, 남아있는 숲 조각과 그 안의 생물들은 종종 매우 고립된다.
더 작거나 더 고립된 삼림 지대는 더 크거나 덜 고립된 삼림 지대에 비해 더 빨리 종을 잃을 것이다.다수의 작은 숲 "섬"은 결합된 면적이 단일 숲보다 훨씬 넓다고 해도 일반적으로 단일 인접 숲이 보유할 수 있는 동일한 생물 다양성을 지원할 수 없다.하지만, 시골 풍경 속의 숲속 섬들은 [61]생물 다양성을 크게 증가시킨다.칠레의 마울리노 숲에서는 파편화가 전체 식물 다양성에 큰 영향을 미치지 않는 것으로 보이며, 실제로 큰 연속 [62][63]숲보다 파편에서 나무의 다양성이 높다.
캐나다 퀘벡 주 몬트리올에 있는 맥길 대학교는 국제 [64]과학자들에 의해 수행된 연구에 따르면, 전 세계에 남아있는 숲의 70%가 숲 가장자리에서 1킬로미터 이내에 위치해 생물 다양성을 엄청난 위험에 처하게 한다는 대학 기반의 신문 성명을 발표했다.
fragment 면적의 감소, 격리, 엣지의 증가로 모든 에코시스템에 퍼지는 변화가 시작됩니다.서식지 조각화는 또한 일부 종의 풍부함이나 많은 강력한 시스템 [5]반응을 식별하기 위해 긴 시간적 척도가 필요한 패턴과 같은 예기치 않은 결과를 형성할 수 있다.
지속가능한 산림관리
숲 파편의 존재는 인접한 농업 분야의 다양한 생태계의 공급에 영향을 미친다(Mitchell et al. 2014).미첼 외 연구진(2014)은 캐나다 퀘벡의 농업 지형에 걸쳐 고립과 크기가 다른 인근 숲 조각별로 분리된 거리를 통해 콩밭의 농작물 생산, 분해, 농약 규제, 탄소 저장, 토양 비옥, 수질 규제 등 6가지 다양한 생태계 요인을 연구했다.da. 지속 가능한 산림 관리는 생태계 서비스를 위한 산림 관리(단순한 프로비저닝을 넘어), 정부 보상 제도, 효과적인 규제 [65]및 법적 프레임워크를 통해 여러 가지 방법으로 달성될 수 있다.숲을 보존하는 유일한 현실적인 방법은 추가적인 손실을 감수하기 위해 지속 가능한 산림 관리를 적용하고 실천하는 것이다.
숲이 제공할 수 있는 목재, 펄프, 종이, 그리고 다른 자원에 대한 산업적 수요가 높기 때문에, 기업들은 그러한 자원을 얻기 위해 숲을 벌채하는 것에 더 많은 접근을 원할 것이다.열대우림연합은 지속 가능한 산림관리에 대한 접근을 효율적으로 할 수 있었고, 1980년대 후반에 이것을 설립했습니다.그들의 보존은 전세계 [66]거의 5억 에이커의 땅을 절약했기 때문에 성공한 것으로 간주되었다.
숲을 보존하기 위해 취할 수 있는 몇 가지 접근과 조치는 침식을 최소화하고, 폐기물을 적절히 처리하며, 유전적 다양성을 유지하기 위해 토종 나무 종을 [66]보존하고, 임야를 보존하는 방법이다.또한 산불도 빈번하게 발생할 수 있으며 산불 발생을 더욱 방지하기 위한 조치도 취할 수 있다.예를 들어 과테말라의 문화적으로나 생태적으로 중요한 페텐 지역에서는 연구원들이 20년 이상의 기간을 발견할 수 있었고, FSC 인증 산림은 인근 보호 지역보다 상당히 낮은 삼림 벌채율을 경험했으며, 산불은 인증 토지의 0.1퍼센트만 영향을 미쳤다.보호 [66]구역의 경우.그러나 산림 부문의 고용과 수확 관행에 관한 단기적인 결정은 생물 [67]다양성에 장기적인 영향을 미칠 수 있다는 점에 유의해야 한다.식림된 숲은 전 세계 산업용 원목 생산량의 약 1/4을 공급하고 20년 이내에 전 세계 생산량의 50%를 차지할 것으로 예상되기 때문에 점점 더 중요해지고 있다(Brown, 1998; Jaakko Poyry, 1999).[68]많은 어려움이 있었지만 산림 인증의 실시는 경제적, 환경적, 사회적 이슈를 수용하고 총체적 개념에 대한 효과적인 인식과 지식을 전 세계에 전파할 수 있다는 점에서 상당히 두드러지고 있다.또한 지속가능성 평가 외에 탄소 [69]흡수원 검증과 같은 다양한 응용 분야를 위한 도구를 제공한다.
서식지의 단편화를 이해하기 위한 접근법
서식지의 단편화와 그 생태학적 영향을 이해하기 위해 일반적으로 두 가지 접근방식이 사용된다.
종지향적 접근법
종지향적 접근법은 특히 개별 종에 초점을 맞추고, 각각의 종들이 환경과 서식지의 변화에 어떻게 반응하는지에 초점을 맞춘다.이 접근법은 개별 종에만 초점을 맞추고 종 [70]간 서식지 조각화의 영향을 폭넓게 볼 수 없기 때문에 제한될 수 있다.
패턴 지향 접근법
패턴지향적 접근법은 종 발생과의 상관관계에 있는 토지 피복과 그 패턴화에 기초한다.경관 패턴화를 위한 연구 모델 중 하나는 Richard Forman에 의해 개발된 패치 매트릭스-코리더 모델이다. 패턴 지향 접근법은 인간의 수단과 활동에 의해 정의된 토지 커버에 초점을 맞춘다.이 모델은 섬의 생물 지리학에서 비롯되었으며 정의된 풍경과 그 안에서 종의 발생 사이의 인과 관계를 추론하려고 합니다.접근법에는 종이나 경관에 걸친 집합적 가정에 한계가 있으며,[71] 이들 사이의 차이를 설명하지 못할 수 있다.
변동 모형
다른 모델은 변동 모델입니다.얼룩덜룩한 풍경은 자연 식생의 대부분을 유지하지만 변경된 서식지의 구배와 혼합된다. 이 서식지 조각화 모델은 일반적으로 농업에 의해 수정된 풍경에 적용된다.부적절한 경관 환경으로 둘러싸인 고립된 서식지 패치로 나타나는 단편화 모델과 대조적으로, 다양화 모델은 남아있는 원래 서식지 근처에 작은 서식지가 남아 있는 농업에 의해 수정된 풍경에 적용된다.그 사이는 원래 서식지를 변형한 초원의 매트릭스입니다.이 지역들은 [73]토종에게 그다지 장벽이 되지 않는다.
「 」를 참조해 주세요.
참고 문헌
- 린덴마이어 D.B & Fischer J (2013) 서식지 단편화 및 경관 변화: 생태와 보존의 합성 (아일랜드 프레스)
레퍼런스
- ^ a b c d e Sahney, S.; Benton, M. J.; Falcon-Lang, H. J. (1 December 2010). "Rainforest collapse triggered Pennsylvanian tetrapod diversification in Euramerica" (PDF). Geology. 38 (12): 1079–1082. Bibcode:2010Geo....38.1079S. doi:10.1130/G31182.1.
- ^ Fahrig, Lenore (2019). "Habitat fragmentation: A long and tangled tale". Global Ecology and Biogeography. 28 (1): 33–41. doi:10.1111/geb.12839. ISSN 1466-8238. S2CID 91260144.
- ^ a b c Fahrig, L (2003). "Effects of habitat fragmentation on biodiversity". Annual Review of Ecology, Evolution, and Systematics. 34: 487–515. doi:10.1146/annurev.ecolsys.34.011802.132419.
- ^ van den Berg LJL, Block JM, Clarke RT, Langsten RHW, Rose RJ. 2001.영국 도싯의 다트포드 워블러(실비아 운다타)에 의한 영역 선택: 식생 유형, 서식지의 단편화, 인구 규모.Biol. 보수당 101:217-28
- ^ a b c Haddad, Nick M.; Brudvig, Lars A.; Clobert, Jean; Davies, Kendi F.; Gonzalez, Andrew; Holt, Robert D.; Lovejoy, Thomas E.; Sexton, Joseph O.; Austin, Mike P.; Collins, Cathy D.; Cook, William M. (2015-03-01). "Habitat fragmentation and its lasting impact on Earth's ecosystems". Science Advances. 1 (2): e1500052. Bibcode:2015SciA....1E0052H. doi:10.1126/sciadv.1500052. ISSN 2375-2548. PMC 4643828. PMID 26601154.
- ^ Fischer, Joern; Lindenmayer, David B. (February 7, 2007). "Landscape Modification and Habitat Fragmentation: A synthesis". Global Ecology and Biogeography. 16 (3): 265–280. doi:10.1111/j.1466-8238.2007.00287.x.
- ^ Fahrig, Lenore (November 2003). "Effects of Habitat Fragmentation on Biodiversity". Annual Review of Ecology, Evolution, and Systematics. 34 (1): 487–515. doi:10.1146/annurev.ecolsys.34.011802.132419.
- ^ a b Lienert, Judit (July 2004). "Habitat fragmentation effects on fitness of plant populations – a review". Journal for Nature Conservation. 12 (1): 53–72. doi:10.1016/j.jnc.2003.07.002.
- ^ Wilcove, David S.; et al. (1998). "Quantifying Threats to Imperiled Species in the United States". BioScience. 48 (8): 607–615. doi:10.2307/1313420. JSTOR 1313420.
- ^ a b Fahrig, L (2013). "Rethinking patch size and isolation effects: the habitat amount hypothesis". J. Biogeogr. 40 (9): 1649–1663. doi:10.1111/jbi.12130.
- ^ a b c Fahrig, L (2017). "Ecological Responses to Habitat Fragmentation Per Se". Annual Review of Ecology, Evolution, and Systematics. 48: 1–23. doi:10.1146/annurev-ecolsys-110316-022612.
- ^ Fahrig, L. (2018) 영국 옥스포드 대학 출판부, Careiva, Marvier, Siliman 편집: 효과적인 보존 과학: Data Not Dogma (Data Not Dogma)
- ^ "Access Electronic Resources". www.library.yorku.ca. doi:10.1016/j.biocon.2018.07.022. S2CID 52839843. Retrieved 2022-03-31.
- ^ a b Rosenzweig, Michael L. (1995). Species diversity in space and time. Cambridge: Cambridge University Press.