Aves
| Aves | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ocorrência: Jurássico Superior–Recente, 150–0 Ma | |||||||||||||
 Diversidade das aves | |||||||||||||
| Classificação científica | |||||||||||||
| |||||||||||||
| Ordens | |||||||||||||
Aves são uma classe de seres vivos vertebrados endotérmicos caracterizada pela presença de penas, um bico sem dentes, oviparidade de casca rígida, elevado metabolismo, um coração com quatro câmaras e um esqueleto pneumático resistente e leve. As aves estão presentes em todas as regiões do mundo e variam significativamente de tamanho, desde os 5 cm do colibri até aos 2,75 m da avestruz. São a classe de tetrápodes com o maior número de espécies vivas, aproximadamente dez mil, das quais mais de metade são passeriformes. As aves apresentam asas, que são mais ou menos desenvolvidas dependendo da espécie. Os únicos grupos conhecidos sem asas são as moas e as aves-elefante, ambos extintos. As asas, que evoluíram a partir dos membros posteriores, oferecem às aves a capacidade de voar, embora a especiação tenha produzido aves não voadoras, como as avestruzes, pinguins e diversas aves endémicas insulares. Os sistemas digestivo e respiratório das aves estão adaptados ao voo. Algumas espécies de aves que habitam em ecossistemas aquáticos, como os pinguins ou a família dos patos, desenvolveram a capacidade de nadar.
Algumas aves, especialmente os corvos e os papagaios, estão entre os animais mais inteligentes do planeta. Algumas espécies constroem e usam ferramentas e passam o conhecimento entre gerações. Muitas espécies realizam migrações ao longo de grandes distâncias. As aves são animais sociais que comunicam entre si com sinais visuais, chamamentos e cantos, e realizam atividades comunitárias como procriação e caça cooperativa, voo em bando e grupos de defesa contra predadores. A grande maioria das espécies de aves são monogâmicas, geralmente durante uma época de acasalamento e por vezes durante vários anos, mas raramente durante toda a vida. Outras espécies são polígamas ou, mais raramente, poliândricas. As aves reproduzem-se através de ovos, que são fertilizados por reprodução sexual e geralmente colocados num ninho onde são incubados pelos progenitores. A maior parte das aves apresenta um período prolongado de cuidados parentais após a incubação. Algumas aves, como as galinhas, põem ovos mesmo que não sejam fertilizados, embora esses ovos não produzam descendência.
As aves, e em particular os fringilídeos de Darwin, tiveram um papel importante no desenvolvimento da teoria da evolução por seleção natural de Darwin. O registo fóssil indica que as aves são os últimos sobreviventes dos dinossauros, tendo evoluído a partir de dinossauros emplumados dentro do grupo terópode dos saurísquios. As primeiras aves apareceram durante o período cretácico, há cerca de 100 milhões de anos,[1] e estima-se que o último ancestral comum tenha vivido há 95 milhões de anos.[2] As evidências de ADN indicam que as aves se desenvolveram extensivamente durante a extinção do Cretáceo-Paleogeno que matou os dinossauros não avianos. As aves na América do Sul sobreviveram a este evento, tendo depois migrado para as várias partes do mundo através de várias passagens terrestres, ao mesmo tempo que se diversificavam em espécies durante os períodos de arrefecimento global.[3] Algumas aves primitivas dentro do grupo Avialae datam do período Jurássico.[4] Muitos destes ancestrais das aves, como o Archaeopteryx, não tinham plena capacidade de voo e muitos apresentavam ainda características primitivas como mandíbula em vez de bico e cauda vertebrada.[4][5]
Muitas espécies de aves têm importância económica. As aves domesticadas (de capoeira) e não domesticadas (de caça) são fontes importantes de ovos, carne e penas. As aves canoras e os papagaios são animais de estimação populares. O guano é usado como fertilizante. As aves são um elemento de destaque na cultura. No entanto, desde o século XVII que cerca de 120 a 130 espécies foram extintas devido à ação humana e várias centenas foram extintas nos séculos anteriores. Atualmente existem 1 375 espécies de aves ameaçadas de extinção, embora haja esforços no sentido de as conservar. A observação de aves é uma atividade importante no setor do ecoturismo.
Habitat, diversidade e distribuição

A capacidade de voar proporcionou às aves uma diversificação extraordinária, pelo que hoje em dia vivem e reproduzem-se em praticamente todos os habitats terrestres e em todos os sete continentes.[6] O petrel-das-neves nidifica em colónias que já foram observadas a distâncias de 440 km do litoral da Antártida.[7] A maior biodiversidade de aves tem lugar nas regiões tropicais. Anteriormente, pensava-se que esta maior diversidade era o resultado de uma maior velocidade de especiação nos trópicos. No entanto, estudos mais recentes verificaram que a especiação é superior nas latitudes mais elevadas, embora a velocidade de extinção seja também superior à dos trópicos.[8] Várias famílias de aves evoluíram para se adaptar à vida nos oceanos. Algumas espécies de aves marinhas regressam à costa apenas para nidificar[9] e alguns pinguins são capazes de mergulhar até 300 metros de profundidade.[10]
Regra geral, o número de espécies que se reproduz em determinada área é diretamente proporcional ao tamanho dessa área e à diversidade de habitats disponíveis. O número total de espécies está também relacionado com factores como a posição dessa área em relação às rotas de migração e ao número de espécies que aí passam o inverno. Na Europa a oeste dos montes Urais, incluindo grande parte da Turquia, vivem cerca de 540 espécies de aves. Na Ásia vivem 2 700 espécies, o que corresponde a 25% da avifauna mundial, e só na Rússia vivem cerca de 700. Em África vivem cerca de 2 300 espécies. Em todo o continente americano vivem cerca de 4 400 espécies, embora em alguns países da América central e do sul haja mais de mil espécies. A Costa Rica é a região com maior número de espécies em relação ao tamanho, com cerca de 800 conhecidas numa área de apenas 51 000 km2.[6]
Muitas espécies de aves estabeleceram o seu território nas regiões em que foram introduzidas pelo ser humano. A introdução de algumas espécies foi deliberada, como por exemplo o faisão-comum, que foi introduzido em todo o mundo como ave de caça.[11] Outras introduções foram acidentais, como o periquito-monge que atualmente está presente em várias cidades norte-americanas como consequência de fugas de cativeiro.[12] Algumas espécies, como a garça-boieira,[13] o gavião-carrapateiro[14] e a cacatua-galah[15] expandiram-se muito para além do seu território inicial de forma natural, à medida que a agricultura foi criando novos habitats.
Anatomia e fisiologia

Em comparação com outros vertebrados, as aves apresentam um corpo com diversas adaptações invulgares e únicas que lhes permitem voar, mesmo que sejam apenas estruturas vestigiais ou que sejam usadas para deslocação terrestre ou aquática. Embora haja várias particularidades ósseas e anatómicas exclusivas das aves, a presença de penas é a mais proeminente e distintiva característica das aves. As aves possuem também um órgão único entre os animais, a siringe, que lhes permite produzir os cantos e chamamentos. A pele das aves é praticamente ausente de quaisquer glândulas, à exceção da glândula uropigial que produz um óleo que protege e impermeabiliza as penas.[6]
As aves são animais endotérmicos que mantêm uma temperatura de aproximadamente 41 °C, podendo ser ligeiramente inferior durante as horas de sono e ligeiramente superior durante os períodos de atividade intensa. As penas e, em algumas espécies, a gordura subcutânea, oferecem isolamento térmico. Como não possuem glândulas sudoríparas, o excesso de calor é dissipado pela respiração rápida que, em algumas espécies, pode chegar às 300 respirações por minuto. Algumas espécies são capazes de hibernar temporariamente.[6]
Nas aves voadoras de maior dimensão, os ossos são permeados por cavidades de ar e o sistema respiratório é constituído por sacos de ar, o que contribui para diminuir o seu peso. O albatroz é a ave voadora com maior envergadura de asas, que pode atingir os 3,5 metros, enquanto o cisne-trombeteiro pode atingir os 17 kg de peso. A ave mais pequena é o colibri-cubano, que pode medir 5–6 cm de comprimento e pesar apenas 3 gramas. Quando as aves perdem a capacidade de voar deixam de estar limitadas pelo peso, como é o caso da avestruz, que pode atingir os 2,75 metros de altura e pesar 150 kg.[6]
Voo

A maior parte das aves tem a capacidade de voar, o que as distingue de praticamente todas as outras classes de vertebrados. O voo é a principal forma de deslocação para a maioria das espécies de aves e é usado para a reprodução, alimentação e fuga de predadores.[16] No entanto, cerca de 60 espécies vivas de aves são incapazes de voar, assim como muitas das aves extintas.[17] A incapacidade de voo ocorre muitas vezes em ilhas isoladas, provavelmente devido aos recursos limitados e à ausência de predadores terrestres.[18] Embora incapazes de voar ou de percorrer grandes distâncias, algumas aves usam a mesma musculatura e movimentos para nadar na água, como é o caso dos pinguins, tordas ou os melros-d'água.[19]
As aves possuem diversas adaptações evolutivas destinadas ao voo, entre as quais ossos pneumáticos e leves, dois grandes músculos de voo (os peitorais – que correspondem a 15% do peso da ave – e o supracoracoide) e um membro anterior modificado (a asa) que atua simultaneamente como aerofólio (que sustenta a ave no ar) e como elemento de impulso. Muitas aves alternam entre o voo impulsionado pelo bater de asas e o voo planado e usam a cauda para direcionar o voo.[16] A asa é bastante flexível, podendo ser alongada ou recolhida por flexão, as penas das bordas podem ser abertas ou fechadas e o ângulo das asas pode ser ajustado, quer em conjunto, quer individualmente.[6]

O tamanho e forma da asa geralmente está relacionado com o tipo de voo dessa espécie. Por exemplo, os albatrozes, as gaivotas e outras as aves marinhas apresentam asas compridas e estreitas que tiram partido do vento sobre o oceano para planar, enquanto que as aves de rapina têm asas amplas para tirar partido das correntes de ar ascendentes e descendentes de montanha. As aves de asa curta, como os galináceos, fazem apenas voos curtos. As asas pontiagudas dos patos permitem-lhes percorrer rapidamente grandes distâncias. As andorinhas e os estorninhos têm asas compridas e pontiagudas que lhes permitem fazer acrobacias aéreas durante horas seguidas. Enquanto algumas aves de grande dimensão, como os abutres, planam ao longo de horas e só ocasionalmente batem as asas, outras aves, como os colibris, batem as asas várias vezes por segundo.[6]
A velocidade de voo varia imenso entre as espécies. As aves mais pequenas, como os pardais ou as carriças, voam entre 15-30 km/h. As aves de pequena a média dimensão, como os tordos, rabos-de-quilha, estorninhos ou andorinhões, e as aves de grande envergadura de asa, como os pelicanos e as gaivotas, voam entre 30 e 60 km/h. As aves mais rápidas, como os falcões, patos, gansos e pombos voam entre 60 a 100 km/h. A ave que atinge maior velocidade é o falcão-peregrino, que é capaz de realizar voos a pique que atingem os 320 km/h.[6]
Muitas aves com pés pequenos, como os andorinhões, colibris ou calaus, raramente caminham e usam as patas apenas para se empoleirarem, enquanto outras apresentam patas mais robustas e realizam grande parte dos movimentos a pé, como as galinhas de água. Os papagaios caminham frequentemente ao longo dos ramos, enquanto espécies como os melros saltitam no terreno. Muitas espécies usam as asas para se deslocar debaixo de água, como é o caso dos pinguins, embora algumas só nadem à superfície. Algumas aves, como as mobêlhas, estão tão adaptadas à vida no mar que só vão a terra nidificar. Muitas aves, como o pato-real são capazes de levantar voo diretamente a partir da superfície da água, embora outras necessitem de fazer uma pequena corrida para ganhar impulso, como os mergulhões.[6]
Penas, plumagem e escamas

As penas são uma característica proeminente e única das aves.[nota 1] As penas permitem às aves voar, fornecem isolamento que facilita a regulação térmica e são usadas como forma de exibicionismo, camuflagem e comunicação.[16] Existem vários tipos de penas, cada um com finalidades específicas. As penas são desenvolvimentos epidérmicos ligados à pele e crescem apenas em regiões específicas da pele denominadas pterilas. O padrão de distribuição da implantação das penas em tratos denomina-se pterilose. Aptérios são zonas desprovidas de penas. A distribuição e aparência do conjunto das penas no corpo é denominado plumagem e é uma das principais características que permitem identificar a espécie de ave. A plumagem pode variar significativamente dentro da própria espécie de acordo com a idade, estatuto social[20] e sexo.[21] A cor das penas tem origem em pigmentos ou na estrutura. Os castanhos e os pretos são causados por melaninas sintetizadas pela ave e depositados em grânulos, enquanto os amarelos, laranjas e vermelhos são causados por carotenoides com origem na dieta e difundidos pela pele e penas. As coes azuis são estruturais e têm origem numa camada fina e porosa de queratina. Os verdes resultam do acréscimo de pigmento amarelo ao azul estrutural. As cores iridescentes têm origem na estrutura laminada das bárbulas e são realçadas pelos depósitos de melanina.[6]
A plumagem das aves muda regularmente. Geralmente a muda é anual, embora em algumas espécies se observem duas mudas, uma antes da época de reprodução e outra depois, podendo ocorrer variações entre espécies e entre indivíduos da mesma espécie.[22] As maiores aves de rapina podem mudar de plumagem apenas uma vez em vários anos. As características da muda variam entre espécies. Em passariformes, as penas de voo são substituídas uma de cada vez, sendo as primárias da face inferior substituídas primeiro. Após a substituição da quinta ou sexta primária, começam a cair as terciárias exteriores, seguidas pelas secundárias e pelas outras penas. As grandes coberturas primárias são substituídas ao mesmo tempo das primárias que cobrem. Este processo denomina-se muda centrífuga.[23] Por outro lado, um pequeno número de espécies, entre as quais os patos e os gansos, perdem todas as penas de voo de uma única vez, ficando temporariamente incapazes de voar.[24] Regra geral, a muda das penas da cauda segue o mesmo padrão da das asas, do interior para o exterior.[23] No entanto, nos Phasianidae verifica-se muda centrípeta (de fora para o centro),[25] enquanto que na cauda dos pica-paus e das trepadeiras começa a muda começa no segundo par mais interior e acaba no par central de penas, de modo a que a ave possa continuar a trepar.[23][26]
As escamas das aves encontram-se principalmente nos dedos e no metatarso, embora em algumas aves estejam presentes acima do tornozelo. À semelhança dos bicos, garras e espigões, são constituídas por queratina. A maior parte das escamas não se sobrepõe de forma significativa, exceto nos casos do guarda-rios e do pica-pau. Pensa-se que as escamas das aves sejam homólogas às dos répteis e dos mamíferos.[27]
Bico

Embora o tamanho e a forma do bico das aves varie significativamente de espécie para espécie, a sua estrutura é idêntica. Todos os bicos são constituídos por duas mandíbulas, superior e inferior. A mandíbula superior está apoiada num osso denominado intermaxilar, cujas ligações permitem que a mandíbula seja flexível e se mova para cima e para baixo. A mandíbula inferior está apoiada num osso denominado osso maxilar inferior.[28]
A superfície exterior do bico é constituída por uma camada fina de queratina, denominada ranfoteca.[28] Entre a ranfoteca e as camadas mais profundas da derme existe uma camada vascularizada ligada diretamente aos ossos do bico.[29] A ranfoteca forma-se nas células na base de cada mandíbula[30] e cresce continuamente na maior parte das aves, enquanto em algumas espécies muda de cor conforme a estação.[31]
As bordas cortantes do bico são denominadas tomia.[32] Na maior parte das espécies, as bordas variam entre arredondadas e ligeiramente afiadas, embora algumas espécies tenham desenvolvido modificações estruturais que lhes permitem manusear melhor a sua fonte de alimentação.[33] Por exemplo, as aves granívoras têm bordas serrilhadas que lhes permitem quebrar a casca de sementes.[34] A maior parte dos falcões e alguns papagaios omnívoros apresentam uma projeção na mandíbula superior, de modo a atingir mortalmente as vértebras das presas ou a dilacerar insetos.[35] Algumas espécies que se alimentam de peixe, como os mergansos, têm bicos serrilhados que mantêm preso o peixe escorregadio.[36] Cerca de 30 famílias de aves insetívoras apresentam cerdas muito curtas em toda a mandíbula que aumentam a fricção e lhes permitem segurar insetos de carapaça dura.[37]
Visão e audição

A visão das aves é geralmente bastante desenvolvida. As aves marinhas apresentam lentes flexíveis, o que lhes permite ver claramente tanto no ar como dentro de água.[16] Algumas espécies têm dupla fóvea. As aves são tetracromáticas, possuindo cones não só sensíveis à luz verde, vermelha e azul como também à luz ultravioleta.[38] Muitas aves exibem padrões de plumagem refletora ultravioleta que são invisíveis ao olho humano. Algumas aves cujos sexos aparentam ser iguais apresentam, na realidade, padrões diferenciados. Os machos do chapim-azul, por exemplo, têm uma coroa refletora ultravioleta que é exposta durante a corte ao levantar as penas da nuca.[39] A luz ultravioleta é também usada na procura de comida. Alguns falcões procuram presas através da deteção dos rastos de urina dos roedores, que refletem a luz ultravioleta.[40]
As pálpebras das aves não são usadas para pestanejar. Em vez disso, o olho é lubrificado por uma terceira pálpebra que se desloca horizontalmente, denominada membrana nictitante.[41] Esta membrana também cobre o olho e funciona como uma lente de contacto em muitas aves marinhas.[16] A retina das aves tem um sistema irrigatório próprio denominado pécten ocular.[16] A maior parte das aves não consegue mover os olhos, embora haja exceções, como o corvo-marinho-de-faces-brancas.[42] As aves com olhos nas partes laterais da cabeça têm um grande campo visual, enquanto que as aves com olhos na parte da frente da cabeça têm visão binocular e são capazes de estimar a profundidade de campo.[43]
O alcance auditivo das aves é limitado.[6] O ouvido não apresenta pavilhão auditivo externo, embora o orifício seja revestido por penas. Em algumas aves de rapina, como as corujas, mochos ou bufos, estas penas formam tufos que se assemelham a orelhas. O ouvido interno apresenta uma cóclea, mas não em espiral como nos mamíferos.[44]
Sistema respiratório e olfato

As aves têm um dos mais complexos e eficientes sistemas respiratórios de todos os animais.[16] Devido ao elevado ritmo metabólico exigido pelo voo, necessitam de um fornecimento constante de grande quantidade de oxigénio. Embora possuam pulmões, a ventilação é assegurada em grande parte por sacos aéreos ligados aos pulmões, que correspondem a 15% do volume do corpo.[45] Embora as paredes dos sacos aéreos não façam trocas gasosas, têm a função de atuar como foles para fazer circular o ar pelo sistema respiratório.[46] No momento da inalação, apenas 25% do ar inalado vai diretamente para os pulmões; os restantes 75% atravessam os pulmões e passam diretamente para um saco de ar posterior que se estende a partir dos pulmões e está ligado às cavidades de ar nos ossos, as quais preenche com ar. Quando a ave expira, o ar usado sai dos pulmões e o ar fresco do saco de ar posterior é forçado a passar para os pulmões. Desta forma, os pulmões são constantemente fornecidos de ar fresco, quer durante a inalação quer durante a expiração.[47]
Os sons das aves são produzidos na siringe, uma câmara muscular constituída por várias membranas timpânicas que diverge da extremidade inferior da traqueia.[48] Em algumas espécies, a traqueia é alongada, o que aumenta o volume das vocalizações e a perceção do tamanho da ave.[49]
A maior parte das aves tem olfato pouco apurado, embora os quivis,[50] os abutres do Novo Mundo[51] e os procelariformes sejam exceções notáveis.[52] A maior parte das aves apresenta duas narinas externas situadas no bico, geralmente com forma circular, oval ou em corte e ligadas à cavidade nasal no interior do crânio. As espécies da ordem dos procelariformes, como os albatrozes, apresentam narinas encerradas em dois tubos na mandíbula superior. No entanto, outras espécies, como os gansos-patola, não têm narinas externas e respiram pela boca. Embora na maior parte das aves as narinas estejam descobertas, em algumas espécies encontram-se cobertas por penas, como nos corvos e nos pica-paus.[53]
Sistema ósseo
O sistema ósseo das aves é constituído por ossos extremamente leves com cavidades de ar ligadas ao sistema respiratório (ossos pneumáticos).[54] Nos adultos, os ossos do crânio estão fundidos entre si e não apresentam suturas cranianas.[16] As órbitas são de grande dimensão e separadas por um septo nasal. A coluna vertebral divide-se nas vértebras cervicais, torácicas, lombares e caudais. As vértebras cervicais (do pescoço), cujo número varia significativamente entre espécies, são particularmente flexíveis. No entanto, o movimento nas vértebras torácicas é limitado e completamente ausente nas vértebras posteriores.[55] As últimas vértebras encontram-se fundidas com a pelve, formando o sinsacro. As costelas são achatadas. O esterno apresenta uma quilha que sustenta os músculos peitorais envolvidos no voo, embora esteja ausente das aves não voadoras. Os membros anteriores encontram-se modificados na forma de asas.[16][56][57]
Sistema digestivo

O sistema digestivo das aves é constituído por um papo, onde é armazenado o conteúdo digestivo, e uma moela que contém pedras, que as aves engolem para moer os alimentos como forma de compensar a falta de dentes.[58] A maior parte das aves apresenta uma digestão bastante rápida, de modo a não interferir com o voo.[59] Algumas aves migratórias adaptaram-se de modo a usar proteínas de várias partes do corpo como fonte de energia adicional durante a migração.[60] Muitas aves regurgitam egregófitos.[61]
Embora a maior parte das aves necessite de água, as características do seu sistema excretor e a ausência de glândulas sudoríparas fazem com que a quantidade necessária seja reduzida.[62] Algumas aves do deserto conseguem satisfazer a necessidade de água exclusivamente a partir da água contida nos alimentos e são tolerantes ao aumento da temperatura corporal, o que lhes permite evitar despender vapor de água em arrefecimento.[63] As aves marinhas estão adaptadas para beber água salgada do mar, possuindo glândulas de sal na cabeça que eliminam o excesso de sal através das narinas.[64]
A maior parte das aves não consegue realizar movimento de sucção da água, pelo que recolhe a água no bico e inclina a cabeça de modo a permitir que a água escorra pela garganta. No entanto, algumas espécies, principalmente de regiões áridas, conseguem beber água sem necessidade de inclinar a cabeça, como é o caso das famílias dos pombos, estrilídeos, coliiformes, toirões e abetardas.[65] Algumas aves do deserto dependem da presença de fontes de água, como os cortiçois, que se agregam à volta de poços de água durante o dia.[66] Algumas espécies levam água às crias humedecendo as penas, enquanto outras a transportam no papo ou a regurgitam juntamente com a comida. As famílias dos pombos, dos flamingos e dos pinguins produzem para as crias um líquido nutritivo denominado leite de papo.[67]
Sistemas reprodutor e urogenital

Tal como os répteis, as aves são seres uricotélicos; isto é, os seus rins filtram os resíduos nitrogenados da corrente sanguínea e excretam-no na forma de ácido úrico (em vez de ureia ou amoníaco) ao longo dos uréteres até ao intestino. As aves não têm bexiga nem uretra exterior, pelo que (à exceção das avestruzes) o ácido úrico é expelido em conjunto com as fezes em dejetos semi-sólidos.[68][69][70] No entanto, algumas aves, como o colibri, podem ser amoniotélicos, excretando a maior parte dos resíduos nitrogenados na forma de amoníaco.[71] Também excretam creatina, em vez de creatinina como os mamíferos.[16] Este material é expulso através da cloaca, em conjunto com os dejetos intestinais.[72][73] A cloaca é um orifício com diversas finalidades. Não só é por aí que são expulsos os dejetos, como maior parte das aves acasalam juntando as cloacas e é também por aí que as fêmeas depositam os ovos.[61]
Os machos possuem dois testículos internos que durante a época de acasalamento aumentam de tamanho centenas de vezes para produzir esperma.[74] As fêmeas de maior parte das famílias possuem um único ovário funcional (o esquerdo), ligado a um oviduto, embora as de algumas espécies tenham dois ovários funcionais.[75] Os machos dos Palaeognathae (à exceção dos quivis), dos Anseriformes (à exceção dos Anhimidae) e, de forma rudimentar, dos Galliformes (mas totalmente desenvolvido nos Cracidae) possuem um pénis, o qual não se observa nas Neoaves.[76][77] Pensa-se que o comprimento esteja relacionado com a competição espermática.[78] Fora do momento da cópula, encontra-se oculto no interior do proctodeu, um compartimento no interior da cloaca.
Sistema circulatório
O sistema circulatório das aves é impulsionado por um coração miogénico de quatro câmaras protegido por um pericárdio fibroso. O pericárdio encontra-se preenchido com fluido seroso que o lubrifica.[79] O coração em si está dividido em duas metades, direita e esquerda, cada uma constituída por uma aurícula e um ventrículo. As aurículas e vetrículos de cada lado estão separados entre si por válvulas cardíacas que impedem que o sangue passe de uma câmara para a outra durante a contração. Sendo miogénico, o ritmo cardíaco é mantido pelas células marca-passo do nó sinusal situado na aurícula direita. O coração das aves é também apresenta arcos musculares, constituídos por camadas espessas de músculo. De forma semelhante ao coração dos mamíferos, o coração das aves é constituído pelas camadas do endocárdio, miocárdio e pericárdio.[79] As paredes auriculares tendem a ser mais delgadas do que as paredes ventriculares, devido à intensa contração ventricular usada para bombear sangue oxigenado pelo corpo. Em comparação com a massa corporal, o coração das aves é geralmente maior do que o dos mamíferos. Esta adaptação permite que seja bombeada maior quantidade de sangue de modo a responder às necessidades metabólicas associadas ao voo.[80] As principais artérias que transportam o sangue do coração têm origem no arco aórtico direito, ao contrário dos mamíferos, em que é o arco aórtico esquerdo que forma esta parte da aorta.[16] A veia cava inferior recebe o sangue dos membros através do sistema venoso portal. Também ao contrário dos mamíferos, os glóbulos vermelhos das aves mantêm o seu núcleo celular.[81]
As aves têm um sistema bastante eficiente de distribuição de oxigénio pelo corpo, uma vez que têm uma superfície de trocas gasosas dez vezes maior do que os mamíferos. Isto faz com que as aves tenham nos vasos capilares mais sangue por unidade de volume dos pulmões do que um mamífero.[80] As artérias das aves são constituídas por músculos elásticos e espessos para resistir à pressão da forte constrição ventricular, que se tornam mais rígidos à medida que se encontram mais afastados do coração. O sangue passa pelas artérias, que sofrem vasoconstrição, em direção às arteríolas, que distribuiem o oxigénio e os nutrientes a todos os tecidos do corpo. À medida que as arteríolas se encontram mais afastadas do coração e próximas dos órgãos e dos tecidos, são cada vez mais ramificadas de modo a aumentar a área de superfície e diminuir a intensidade da corrente sanguínea. A partir das arteríolas, o sangue desloca-se para os vasos capilares onde ocorre a troca de oxigénio pelos resíduos de dióxido de carbono. Após a troca gasosa, o sangue deoxigenado passa pelas vénulas e é transportado pelas veias de volta ao coração. Ao contrário das artérias, as veias são finas e rígidas, uma vez que não necessitam de resistir a pressões elevadas.[82]
Defesa e combate

Algumas espécies são capazes de usar defesas químicas contra predadores. Alguns Procellariiformes projetam um óleo estomacal desagradável contra agressores,[83] enquanto algumas espécies de pitohuis da Nova Guiné têm a pele e penas revestidas por uma poderosa neurotoxina.[84] A plumagem de algumas espécies envia sinais intimidatórios a potenciais predadores, como o pavão-do-pará, cujas penas criam a ilusão de um grande predador de modo a afugentar falcões e proteger as crias.[85]
Ocasionalmente, os combates entre espécies resultam em ferimentos ou morte.[86] Os anhimídeos, algumas jaçanãs, o pato-ferrão, o pato-das-torrentes e nove espécies de abibes apresentam um espigão afiado na asa que usam como arma. Os patos-vapor, os cisnes e gansos, as pombas do ártico, os motum e os alcaravões apresentam uma protuberância óssea na álula para esmurrar os oponentes.[86] Algumas jaçanãs apresentam um rádio expandido em forma de lâmina. O extinto ibis jamaicano apresentava um membro anterior alongado que provavelmente funcionava em combate de forma semelhante a um malho. Outras aves, como os cisnes, são capazes de morder ao defender os ovos ou as crias.[86]
Comportamento
A maior parte das aves é diurna, embora algumas aves sejam noctívagas ou crepusculares, como as corujas, e muitas aves limícolas se alimentem de acordo com as marés, seja de dia ou de noite.[87] A socialização entre indivíduos varia imenso de espécie para espécie. Muitas aves de rapina são solitárias, enquanto outras espécies são vincadamente gregárias, atingindo comunidades de milhares de indivíduos. Durante a época de reprodução, muitas espécies defendem um território. Muitos comportamentos das aves são universais e ocorrem invariavelmente em praticamente todas as espécies de aves, como é o caso dos cuidados com as penas, os movimentos de distensão das pernas, asas ou caudas, os banhos de água e poeira, ou o movimento que sacode o corpo.[6]
Alimentação


As dietas das aves são bastante variadas, podendo incluir néctar, fruta, plantas, sementes, carcaças e vários animais de menor dimensão, incluindo outras aves.[16] Uma vez que as aves não têm dentes, o seu sistema digestivo encontra-se adaptado de modo a processar alimentos engolidos inteiros sem mastigar. As aves que usam várias estratégias para adquirir comida ou se alimentam de vários alimentos são denominadas generalistas. As que dedicam tempo e esforço na procura de alimentos específicos são denominadas especialistas.[16] As estratégias de alimentação variam de espécie para espécie. Muitas aves simplesmente recolhem insetos, invertebrados, fruta ou sementes. Algumas caçam, surpreendendo insetos com ataques súbitos a partir de ramos de árvores.[88]
As aves que se alimentam do néctar das flores, como os beija-flores, apresentam línguas longas e especialmente adaptadas para a recolha do néctar e, em muitos casos, os próprios bicos foram-se adaptando de forma a encaixar em determinadas flores – fenómeno denominado coevolução.[89] Os quivís e as aves limícolas têm bico longos que lhes permitem vasculhar o terreno à procura de pequenos invertebrados. Os diferentes comprimentos do bico e diferentes métodos de procura fazem com que as espécies limícolas se encontrem muitas vezes em nichos ecológicos distintos.[16][90] Algumas espécies, como as mobelhas, os zarros, os pinguins ou as tordas, perseguem a presa debaixo de água, usando as asas e as patas como meio de propulsão.[9] Os predadores aéreos, como os sulídeos, os guarda-rios ou as andorinhas-do-mar mergulham em queda livre atrás das presas. Os flamingos, três espécies de priões e alguns patos filtram os alimentos da água.[91][92]
Algumas espécies, como as fragatas, gaivotas,[93] ou os moleiros[94] praticam cleptoparasitismo, roubando comida de outras aves. No entanto, pensa-se que o cleptoparasitismo seja apenas um complemento à comida obtida através da caça, e não a principal fonte de alimentação.[95] Outras espécies são necrófagas. Entre estas, algumas, como os abutres, são especializadas no consumo de carcaças, enquanto outras, como as gaivotas, corvídeos ou determinadas aves de rapina, são oportunistas.[96]
Cuidados com as penas

As penas exigem manutenção constante. Para além do desgaste físico, as penas são atacadas por fungos, parasitas e piolhos.[97] As aves conservam as penas com a limpeza, aplicação de secreções protetoras e banhos de água ou pó. Enquanto algumas aves apenas se banham em água rasa, como bebedouros ou fontes, outras mergulham em águas profundas e e algumas espécies arbustivas aproveitam a água da chuva que se acumula nas folhas. As aves de regiões áridas usam o solo para tomar banhos de poeira. Algumas espécies encorajam formigas a percorrer as penas, reduzindo o número de ectoparasitas. Muitas espécies abrem frequentemente as asas expondo-as à luz direta do sol, o que diminui o número de parasitas e previne o aparecimento de fungos. Algumas aves esfregam as formigas nas penas, o que faz com que libertem ácido fórmico que mata alguns parasitas.[98][99][100]
As aves limpam, alisam e tratam das penas todos os dias, despendendo neste processo cerca de 9% das horas de atividade.[101] O bico é usado para escovar partículas estranhas e para aplicar secreções de uma substância oleosa produzida pela glândula uropigial que mantém as penas flexíveis e atua como agente antimicrobiótico que impede o crescimento de bactérias que degradam as penas.[102]
Migração
Muitas espécies de aves migram de forma a tirar partido das diferenças sazonais de temperatura que influenciam a disponibilidade de fontes alimentares e dos habitats de reprodução. Estas migrações variam significativamente entre diferentes grupos. Muitas aves terrestres, aves limícolas e aves marinhas realizam migrações anuais ao longo de grandes distâncias, geralmente iniciadas com a mudança do tempo ou da quantidade das horas de luz. Estas aves geralmente alternam entre uma época de reprodução passada nas regiões de clima temperado ou polar e outra época não reprodutiva passada em regiões tropicais ou no hemisfério oposto. Antes da migração, as aves aumentam substancialmente as reservas de gordura corporal e diminuem o tamanho de alguns dos seus órgãos.[60][103] A migração exige uma elevada quantidade de energia, sobretudo quando as aves atravessam desertos e oceanos sem se poder reabastecer. As aves terrestres têm um alcance de voo de cerca de 2500 km e as aves costeiras até 4000 km,[104] embora o fuselo seja capaz de voar 10 200 km sem paragens.[105] Algumas aves marinhas realizam também grandes migrações, a mais longa das quais é realizada pela pardela-preta, que nidifica na Nova Zelândia e no Chile e durante o verão se alimenta no Pacífico norte ao largo do Japão, do Alasca e da Califórnia, viajando por ano 64 000 km.[106] Outras aves marinhas dispersam-se após a reprodução, viajando significativamente sem uma rota de migração fixa. Por exemplo, os albatrozes que nidificam no Oceano Austral realizam viagens circumpolares entre as épocas de acasalamento.[107]
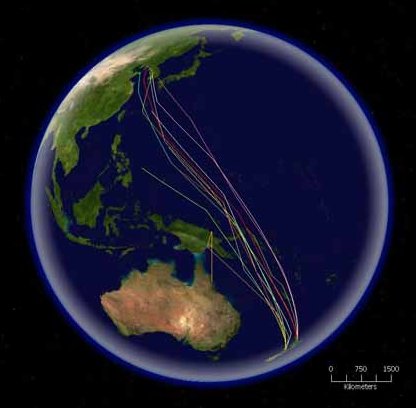
Outras espécies realizam migrações mais curtas, viajando apenas o que for necessário para evitar o mau tempo ou obter comida. As espécies irruptivas, como os fringilídeos boreais, são um desses grupos, sendo possível que num ano permaneçam em determinado local e no seguinte não. Este tipo de migração está normalmente associado à disponibilidade de comida.[108] Algumas espécies podem também viajar apenas dentro da sua área de distribuição, neste caso migrando os indivíduos de maiores latitudes para os locais da sua espécie a menores latitudes. Outras ainda realizam apenas migrações parciais, em que migra apenas uma parte da população, geralmente as fêmeas e os machos subdominantes.[109] Em algumas regiões, as migrações parciais podem representar uma parte significativa dos movimentos migratórios. Na Austrália, por exemplo, 44% dos não passeiriformes e 32% dos passeiriformes realizam apenas migrações parciais.[110] A migração altitudinal é uma forma de migração de curta distância em que as aves passam a época de reprodução nas altitudes mais elevadas e em condições menos favoráveus migram para zonas mais baixas. Este tipo de migração é desencadeado por alterações na temperatura e quando o território normal se torna inóspito devido à falta de comida.[111] Outras espécies são nómadas, não tendo um território definido e deslocando-se de acordo com o tempo e a disponibilidade de comida.[112]
As aves têm a capacidade de regressar exatamente à mesma localização de onde partiram, mesmo após percorrem grandes distâncias.[113] Durante a migração, as aves navegam através de diversos métodos. Os migrantes diurnos orientam-se pelo Sol durante o dia e pelas estrelas durante a noite. As que usam o sol compensam a deslocação do astro usando um relógio interno,[16] enquanto que a orientação pelas estrelas depende da posição das constelações que circundam a estrela polar.[114] Para além disto, algumas espécias têm a capacidade de sentir o geomagnetismo terrestre através de fotorrecetores especiais.[115]
Comunicação
 |
|
| Problemas para escutar este arquivo? Veja a ajuda. | |
As aves comunicam principalmente através de sinais visuais e auditivos. Muitas espécies usam a plumagem para procurar ou reafirmar o estauto social,[116] para mostrar que se encontram na época de acasalamento ou para enviar sinais intimidatórios aos predadores.[117] As variações na plumagem também permitem identificar as aves e diferenciar espécies. Os rituais de exibição fazem também parte da comunicação visual entre as aves, podendo ser usados para demonstrar agressão, submissão ou como forma de criar laços entre indivíduos.[16] Os rituais mais sofisticados ocorrem durante o cortejo sexual e são acompanhados por danças de movimentos complexos.[118] O êxito do macho em acasalar pode depender da qualidade destes rituais.[119]
Os cantos e chamamentos são o principal meio de comunicação sonoro das aves e podem ser bastante complexos. Os sons são produzidos na siringe. Algumas espécies conseguem usar os dois lados da siringe de forma independente, o que lhes permite produzir dois sons diferentes em simultâneo.[48] Os chamamentos são usados para as mais diversas finalidades, entre as quais a atração do macho durante a época de reprodução,[16] a avaliação de potenciais parceiros,[120] a criação de laços afetivos, a reivindicação e manutenção territorial,[16] e identificação de outros indivíduos (como quando os progenitores procuram as crias nas colónias ou quando os parceiros se reúnem no início da época de reprodução),[121] e aviso às outras aves de potenciais predadores, por vezes com informação específica sobre a natureza da ameaça.[122] Algumas espécies também usam sons mecânicos para comunciar. As narcejas neo-zelandesas produzem sons fazendo o ar circular por entre as penas,[123] os pica-paus tamboreiam de forma territorial,[59] e as cacatuas-das-palmeiras usam instrumentos de percussão.[124]
Comportamento de grupo

Enquanto algumas aves são essencialmente territoriais ou vivem em pequenos grupos familiares, outras aves formam bandos de grande dimensão. Os principais benefícios dos bandos são a segurança pelo número e a maior eficácia na procura de comida.[16] A defesa contra eventuais predadores é especialmente relevante em habitats fechados, como nas florestas, em que é a predação por emboscada é comum e um grupo grande aumenta a vigilância. Existem também bandos dedicados à procura de comida que juntam várias espécies que, embora ofereçam segurança, também aumentam a competição pelos recursos.[126]
No entanto, o agrupamento em bando também potencia o assédio, a dominância de algumas aves em relação a outras e, em alguns casos, uma diminuição na eficácia da procura por comida.[127]
As aves por vezes também formam associações com espécies não aviárias. Algumas aves marinhas que caçam por mergulho associam-se a golfinhos e atuns que empurram os cardumes de peixe em direção à superfície.[128] Os calaus têm uma relação mutualista com os mangustos-anão, na qual caçam em conjunto e avisam-se entre si em relação a aves de rapina e outros predadores.[129]
Repouso e empoleiramento

O elevado metabolismo das aves durante a parte ativa do dia é compensado pelo repouso no restante tempo. Ao dormir, as aves incorrem num tipo de sono denominado "sono de vigília", em que os períodos de descanso são intercalados com rápidas "espreitadelas" de olhos abertos, o que lhes permite aperceber de eventuais distúrbios e rapidamente escapar de ameaças.[130] Tem sido sugerido que possa haver determinados tipos de sono que são possíveis até mesmo durante o voo.[131] Por exemplo, acredita-se que os andorinhões sejam capazes de dormir em pleno voo, e as observações de radar sugerem que se orientam na direção do vento durante o sono.[132] Algumas aves têm também demonstrado a capacidade de ter de forma alternada apenas um dos hemisférios cerebrais em sono profundo, o que permite ao olho oposto ao hemisfério que está a dormir continuar vigilante. As aves tendem a exercer esta capacidade quando estão nas orlas dos bandos.[133] O empoleiramento comunitário é comum, uma vez que diminui a perda de calor corporal e diminui os riscos associados com os predadores.[134] Os locais de poleiro são geralmente escolhidos tendo em conta a segurança e regulação de calor.[135]
Durante o sono, muitas aves dobram a cabeça e o pescoço sobre as costas e enfiam o bico nas penas do dorso, enquanto outras o fazem nas penas do peito. Muitas aves descansam apoiadas apenas numa perna, enquanto algumas puxam as pernas para baixo das penas, especialmente no inverno. Os pássaros apresentam um mecanismo que bloqueia o tendão que os ajuda a manterem-se no poleiro enquanto dormem. Muitas aves que vivem junto ao solo, como as codornizes e os faisões, empoleiram-se nas árvores. Alguns papagaios do género Loriculus empoleiram-se de cabeça para baixo.[136] Mais de uma centena de espécies, como o colibri ou noitibó, descansam num estado de torpor acompanhado de uma diminuição do metabolismo.[137] Uma espécie, o noitibó-de-nuttall, chega a entrar num estado de hibernação.[138]
Reprodução
As aves têm dois sexos: macho e fêmea. O sexo das aves é determinado pelos cromossomas Z e W, em vez dos cromossomas X e Y presentes nos mamíferos. Os machos têm dois cromossomas Z (ZZ) e as fêmeas um cromossoma W e um Z (WZ).[16]
Acasalamento


As aves apresentam diversos comportamentos de acasalamento, estando identificados muitos tipos de seleção sexual entre as aves. Na seleção intersexual é a fêmea que escolhe o parceiro, enquanto na competição intrasexual os indivíduos do sexo mais abundante competem entre si pelo privilégio de acasalar,[139] O acasalamento de maior parte das espécies envolve alguma forma de corte sexual, geralmente por parte do macho.[140] A maior parte dos rituais de acasalamento são bastante simples e envolvem apenas algum tipo de canto. No entanto, alguns são coreografias bastante complexas e, dependendo da espécie, podem incluir dança, bater de asas ou cauda, voos aéreos ou lek comunitário.[139] Após a escolha do parceiro, muitas espécies fazem ofertas de objetos entre os parceiros ou juntam os bicos em sinal de afeto.[59]
Os ornamentos de seleção sexual muitas vezes evoluem para se tornar mais pronunciados em situações de competição, até ao ponto em que começam a limitar a agilidade do indivíduo. O elevado custo dos comportamentos e ornamentos sexuais exagerados assegura que apenas os indvíduos de melhor qualidade os conseguem apresentar.[141] A cauda do pavão é provavelmente o exemplo mais conhecido de um ornamento sexual em aves. Os dimorfismos sexuais, como a diferença de tamanho e de cor, são bastante comuns entre as aves e um indicador de forte competição reprodutiva.[142]
Noventa e cinco por cento das espécies de aves são socialmente monógamas. Os casais destas espécies estão juntos pelo menos durante uma época de reprodução ou, em alguns casos, durante vários anos ou até à morte de um dos parceiros.[143] A monogamia permite que o pai também cuide das crias, o que é fundamental nas espécies em que as fêmeas necessitam da assistência dos machos durante a incubação.[144] No entanto, entre as muitas espécies monógamas a infidelidade é comum.[145] Este comportamento verifica-se geralmente entre os machos dominantes e as fêmeas de machos subordinados, embora possa também ser o resultado de cópula forçada em patos e outros anatídeos.[146] As fêmeas das aves têm mecanismos de armazenamento de esperma que permitem que o esperma do macho se mantenha viável durante bastante tempo depois da cópula, chegando a 100 dias em algumas espécies,[147] e que haja competição entre o esperma de vários machos. Para as fêmeas, entre os possíveis benefícios da cópula extra-par está a possibilidade de melhores genes para a prole e garantia contra uma eventual infertilidade do parceiro.[148] Os machos das espécies que praticam cópula extra-par vigiam de perto a companheira de modo a garantir a paternidade da prole que criam.[149] Nos restantes 5% de espécies verificam-se vários sistemas de acasalamento, incluindo poliginia, poliandria, poligamia, poliginandria e promiscuidade. O acasalamento polígamo ocorre quando as fêmeas são capazes de criar a prole sem a ajuda dos machos.[16] Foi também observado comportamento homossexual em machos ou fêmeas de várias espécies de aves, incluindo cópula, formação de pares e nos cuidados com as crias.[150]
Territórios, nidificação e incubação


Durante a época de reprodução, muitas aves defendem ativamente um território de outros indivíduos da mesma espécie, assegurando assim fontes de comida para as suas crias. As espécies que não são territoriais, como as aves marinhas ou os andorinhões, geralmente reproduzem-se em colónias que oferecem proteção contra predadores. As aves coloniais defendem apenas o ninho, embora a competição pelos melhores locais para construir o ninho possa ser intensa.[151]
As aves geralmente põem os ovos num ninho. A maior parte das espécies constrói ninhos bastante elaborados, que podem ser em forma de chávena, de prato, em montículo ou dentro de uma toca.[152] No entanto, alguns ninhos são extremamente primitivos. O ninho do albatroz, por exemplo, pouco mais é do alguma terra e ramos secos no chão. A maior parte das aves constrói ninhos em locais abrigados ou escondidos para evitar predadores, embora as aves coloniais ou de grande dimensão, que têm maior capacidade de defesa, possam construir ninhos abertos. Os materiais de construção do ninho variam imenso, incluindo pequenas pedras, lama, ervas, galhos, folhas, algas e fibras de plantas, sendo usados tanto isoladamente como em conjunto. Algumas aves procuram também materiais de origem animal, como penas, pêlo de cavalos ou pele de cobras. Os materiais podem ser entrançados, cosidos ou unidos com lama.[6] Algumas espécies procuram plantas com determinadas toxinas que impedem a propagação de parasitas, de modo a aumentar a sobrevivência das crias,[153] e penas, que oferecem isolamento térmico.[152] Algumas espécies não fazem ninho; por exemplo, o airo põe os ovos na rocha nua e os pinguins-imperador mantêm os ovos entre o corpo e as patas.[154]
Todas as aves põem ovos amnióticos de casca dura, constituída principalmente por carbonato de cálcio.[16] O número de ovos postos em cada ninhada varia entre 1 e cerca de 20, dependendo da espécie. Algumas espécies põem invariavelmente o mesmo número de ovos por ninhada, embora a maioria das aves ponha um número variável. O número de ovos tende a ser menor nas espécies de regiões tropicais do que nas de regiões mais frias.[6] As espécies que constroem o ninho enterrado ou em buracos tendem a por ovos brancos ou claros, enquanto as espécies de ninho aberto põem ovos camuflados. No entanto, existem muitas exceções a este padrão; por exemplo, os noitibós, que colocam os ovos no chão, têm ovos claros e a camuflagem é oferecida pelas próprias penas. As espécies que são vítimas de parasitas de ninhada têm ovos de várias cores de modo a aumentar a probabilidade de detetar o ovo de um parasita, o que obriga as fêmeas parasitas a fazer corresponder os seus ovos com os dos seus hospedeiros.[155]
Antes da nidificação, as fêmeas de muitas espécies desenvolvem uma placa de incubação, perdendo penas no abdómen. A pele desta região é bastante irrigada com vasos sanguíneos, o que ajuda a ave durante a Incubação.[156] A incubação dos ovos tem início após a postura do último ovo e tem como finalidade normalizar a temperatura de desenvolvimento da cria.[16] Em espécies monogâmicas a tarefa de incubação é geralmente partilhada entre o casal, enquanto que nas espécies polígamas só um dos progenitores é que é responsável pela incubação. O calor dos progenitores passa para o ovo através de placas de incubação – áreas de pele no abdómen ou peito das aves ricas em vasos sanguíneos. A incubação pode ser um processo exigente em termos energéticos; por exemplo, um albatroz adulto chega a perder 83 g de peso por dia durante a incubação.[157] No entanto, os megápodes aquecem os ovos com energia do sol, da decomposição orgânica ou aproveitando a energia geotérmica.[158] Os períodos de incubação variam entre os 10 dias (nos pica-paus, cucos e passeriformes) e os 80 dias (em albatrozes e quivís).[16]
Cuidados parentais e crias


No momento da eclosão dos ovos, dependendo da espécie, as crias apresentam vários estádios de desenvolvimento, desde completamente indefesas até perfeitamente independentes. As crias indefesas são denominadas altriciais e tendem a nascer pequenas, cegas, imóveis e ainda sem penas. As crias que nascem já com autonomia, mobilidade e penas são denominadas precociais ou nidífugas. As crias altriciais necessitam de ajuda para regular a temperatura do corpo e são cuidadas durante mais tempo do que as precociais. As crias que não se enquadram nestes extremos podem ser semi-precociais ou semi-altriciais.[159]
A natureza e duração dos cuidados parentais variam sinificativamente entre as diferentes ordens e espécies. Num dos extremos, os cuidados parentais nos megápodes terminam no momento da eclosão. As crias saem do ovo e escavam sozinhas o caminho para fora do montículo que serve de ninho, começando imediatamente a alimentar-se por si mesmas.[160] No outro extremo, muitas aves marinhas prestam cuidados parentais ao longo de um grande período de tempo. O mais longo é o da fragata-grande, cujas crias levam seis meses até estarem prontas a voar e são alimentadas pelos progenitores ainda por mais 14 meses.[161] O período de guarda é o período imediatamente a seguir à eclosão em que um dos adultos está permanentemente presente no ninho. O propósito principal deste período é ajudar as crias a regular a temperatura e protegê-las dos predadores.[162]
Em muitas espécies, são ambos os progenitores que cuidam das crias. Noutras, os cuidados são da responsabilidade de apenas um dos sexos. Em algumas espécies, outros membros da mesma espécie ajudam a criar a prole, geralmente parentes próximos do casal, como um descendente de uma ninhada anterior.[163] Este comportamento é particularmente comum entre os corvídeos,[164] embora tenha também sido observada em espécies tão diversas como a carriça-da-nova-zelândia ou o milhafre-real. Embora na maioria dos animais sejam raros os cuidados parentais por parte do macho, nas aves são bastante comuns e mais do que em qualquer outra classe de vertebrados.[16] Embora a defesa do território, do ninho, a incubação e a alimentação das crias sejam geralmente tarefas partilhadas entre o casal, por vezes verifica-se uma divisão do trabalho, em que um dos parceiros realiza toda ou grande parte de determinada tarefa.[165]
O momento em que as penas e os músculos das crias estão suficientemente desenvolvidos para permitir voar varia de forma muito significativa. As crias das tordas abandonam o ninho na noite imediatamente a seguir à eclosão dos ovos, seguindo os progenitores em direção ao mar, onde são criados fora do alcance de predadores terrestres.[166] Mas na maior parte das espécies as crias abandonam o ninho imediatamente antes ou pouco depois de serem capazes de voar. O período de cuidados parentais após as crias conseguirem voar varia. Por exemplo, enquanto as crias de albatroz abandonam o ninho por elas próprias e não recebem mais ajuda, outras espécies continuam a alimentar as crias.[167] As crias de algumas espécies migratórias seguem os pais ao longo da sua primeira migração.[168]
Parasitismo de ninhada

O parasitismo de ninhada, ou nidoparasitismo, descreve o comportamento de algumas espécies que põem os ovos nas ninhadas de outras espécies. Este comportamento é mais comum entre aves em comparação com outros animais.[169] Após a ave parasita ter posto os ovos no ninho de outra ave, esses ovos são muitas vezes aceites e cuidados pelo hospedeiro e com prejuízo para a sua própria ninhada. Os nidoparasitas dividem-se entre os que o fazem por necessidade, uma vez que são incapazes de criar as suas próprias crias, e os ocasionais, que apesar de serem capazes de criar a prole põem ovos em ninhos de espécies coespecíficas para aumentar a sua capacidade reprodutora.[170] Entre os parasitas ocasionais estão mais de cem espécies de aves, incluindo espécies das famílias Indicatoridae, Icteridae e o pato-de-cabeça-preta, embora o exemplo mais conhecido sejam os cucos.[169] Algumas crias de nidoparasitas eclodem antes das crias dos hospedeiros, o que lhes permite destruir os ovos, empurrando-os para fora do ninho ou matando as crias. Isto assegura que toda a comida levada para o ninho servirá para alimentar as crias dos nidoparasitas.[171]
Ecologia

As aves ocupam uma ampla variedade de posições ecológicas.[125] Enquanto algumas aves são generalistas, outras são especializadas em relação ao habitat ou à alimentação. Mesmo num único habitat, como por exemplo numa floresta, os nichos ecológicos ocupados pelas diferentes espécies variam. Enquanto algumas espécies vivem no dossel florestal, outras vivem nos estratos intermédios e outras ainda junto ao solo. Cada um destes grupos pode conter aves insetívoras, frugívoras e nectarívoras. As aves aquáticas geralmente alimentam-se da pesca, de plantas ou por cleptoparasitismo. As aves de rapina especializam-se na caça de pequenos mamíferos ou outras aves, enquanto os abutres são necrófagos especializados.[172] Alguns nectarívoros são polinizadores importantes e muitos frugívoros são essenciais para a dispersão de sementes.[173] As plantas e as aves polinizadoras muitas vezes coevoluem,[174] e, em alguns casos, o principal polinizador de determinada flor é a única espécie de ave capaz de alcançar o seu néctar.[175]
As aves são particularmente importantes para a ecologia das ilhas. Tendo sido capazes de migrar para ilhas às quais os mamíferos não conseguiram chegar, as aves podem desempenhar papéis ecológicos que nos continentes são geralmente realizados por animais de maior porte. Por exemplo, as extintas moas tiveram de tal forma impacto no ecossistema da Nova Zelândia, assim como têm o kereru ou o kokako na atualidade,[173] pelo que ainda hoje as plantas da ilha mantêm as adaptações defensivas que as protegiam das moas.[176] Durante a nidação, as aves marinhas também afetam significativamente a ecologia das ilhas e do mar envolvente, principalmente devido à concentração de grandes quantidades de guano que enriquece o solo da região.[177][178]
Relação com o ser humano
Algumas aves têm bastante visibilidade e são animais comuns com os quais o ser humano tem uma estreita relação desde o início da Humanidade.[179] Em alguns casos, estas relações são mutualistas, como na recolha de mel conjunta entre os Indicatoridae e alguns povos africanos, como os Boranas.[180] Noutros casos, as relações podem ser comensais, como o benefício que o pardal-doméstico tira das atividades humanas.[181] No entanto, as aves podem também ser vetores que propagam ao longo de grandes distâncias doenças como a ornitose, salmonelose, campilobacteriose, micobacteriose (tuberculose aviária), gripe das aves, giardiose ou criptosporidíase. Algumas destas são doenças zoonóticas que podem também ser transmitidas para os seres humanos.[182]
Importância económica


As aves de criação são a principal fonte de proteínas animais consumida pelo ser humano. Em 2003, foram consumidos 76 milhões de toneladas de aves de criação e produzidas 61 milhões de toneladas de ovos em todo o mundo.[183] Os frangos representam a maior parte do consumo de aves, embora o peru, o pato e os gansos domesticados sejam também comuns. Muitas espécies de aves são também caçadas para comida. A caça de aves é essencialmente uma atividade recreativa, exceto em regiões bastante subdesenvolvidas. As aves de caça mais comuns são os patos selvagens, faisões, perus selvagens, perdizes, pombos, tetrazes, maçaricos e galinholas.[184] Embora alguma caça possa ser sustentável, a caça no geral tem provocado a extinção ou colocado em risco dezenas de espécies.[185] Entre os outros produtos avícolas com valor comercial estão as penas, especialmente as dos gansos e patos, que são usadas como isolamento em casacos e na roupa de cama, e o guano (fezes de aves marinhas), que é uma fonte valiosa de fósforo e nitrogénio. A Guerra do Pacífico foi travada devido em parte ao controlo dos depósitos de guano.[186]
As espécies que se alimentam de pragas são muitas vezes usadas em programas de controlo biológico.[88] Por outro lado, algumas espécies tornaram-se elas próprias pragas agrícolas de elevado custo económico,[187] enquanto outras constituem riscos para a aviação.[188]
As aves são domesticadas pelo ser humano, não só como animais de estimação mas também para efeitos utilitários. Muitas aves com cores exóticas, como os papagaios ou as araras, são criadas em cativeiro ou vendidas como animais de companhia. No entanto, isto também tem feito crescer o tráfico ilegal de várias espécies ameaçadas.[189] A falcoaria e a pesca com a assistência de corvos marinhos são tradições milenares. Os pombos-correio, usados pelo menos desde o século I, foram essenciais nas comunicações até à II Guerra Mundial, No entanto, hoje em dia estas atividades são mais comuns como passatempo ou atração turística.[190] Existem milhões de entusiastas amadores que apreciam a observação de aves, ou birdwatching, uma vertente significativa do ecoturismo.[191] Muitos proprietários constroem alimentadores de pássaros perto de casa para atrair várias espécies.[192]
Religião, folclore e cultura

As aves têm um papel de relevo no folclore, na religião e na cultura popular e vários dos seus atributos, reais ou imaginários, são usados de forma simbólica na arte e na literatura.[6][193] Desde as gravuras rupestres da época pré-histórica que são um motivo frequente de representação artística e, o longo dos séculos, foram usadas como motivo nas mais diversas formas de arte sacra ou simbólica, como por exemplo o Trono do Pavão dos imperadores mogois e persas.[194] As Fábulas de Esopo contêm várias personificações de aves. Os fisiólogos e os bestiários medievais contêm lições de moral que usam as aves como símbolos para transmitir ideias.[6] O tema central do Conto do Velho Marinheiro, de S. T. Coleridge é a relação entre um albatroz e um marinheiro.[195] Algumas metáforas na linguagem têm origem no comportamento das aves, como a associação de investidores ou fundos predatórios aos abutres necrófagos.[196] As aves são também um tema importante na poesia. Homero escreveu sobre os rouxinóis na Odisseia, enquanto Cátulo usou um pardal como símbolo erótico nos seus poemas.[197]
Na religião, as aves são muitas vezes associadas a mensageiros, sacerdotes ou líderes de determinada divindade. No Cristianismo, as aves simbolizam a transcendência da alma e, na iconografia medieval, uma ave envolta em folhas simbolizava a alma envolvida pelo materialismo do mundo secular.[6] Na mitologia nórdica, Hugin e Munin são dois corvos que deram a volta ao mundo para trazer notícias ao deus Odin. Em várias civilizações da Antiguidade italiana, principalmente na mitologia etrusca e na religião romana, os sacerdotes praticavam auguria, interpretando as palavras das aves enquanto o auspex[nota 2] as usava para fazer previsões sobre o futuro.[198] As aves podem também servir como símbolos religiosos, como no caso de Jonas (hebraico: יוֹנָה, pomba) que simbolizava o medo, passividade, pesar e beleza tradicionalmente associados às pombas.[199] As próprias aves são muitas vezes deificadas, como no caso do pavão-comum, que os Dravidianos vêm como a Terra-Mãe.[200] Algumas aves são também vistas como monstros, incluindo o mitológico Roc e o lendário Poukai, uma ave gigante que para os Maoris é capaz de raptar seres humanos.[201]
Os maias e os Astecas veneravam o quetzal-resplandecente, que hoje em dia dá nome à moeda da Guatemala e é um motivo popular na arte, tecidos e joalharia. As águias são símbolo de poder e prestígio em muitas partes do mundo, incluindo na Europa, onde são frequentemente motivos heráldicos. Os povos nativos norte-americanos usavam frequentemente penas de águia em rituais religiosos, como ornamento pessoal e eram oferecidas aos convidados como símbolo de paz e amizade.[6] No culto do Makemake os tangata manu (homens-pássaro) da ilha de Páscoa eram nomeados líderes.[202] Os povos indígenas dos Andes contam lendas de pássaros que atravessam mundos metafísicos. Em imagens religiosas dos impérios Inca e Tiwanaku, as aves são representadas a transgredir a fronteira entre o reino terrestre e o reino espiritual subterrâneo.[203][204]
As percepções culturais sobre diversas espécies de aves são muitas vezes diferentes de cultura para cultura. Por exemplo, enquanto em África e na mitologia norte-americana as corujas são associadas a má sorte, bruxaria e morte,[205] na Europa são vistas como sábias.[206] As poupas, consideradas sagradas no Antigo Egito e símbolo de virtude na Pérsia, são no entanto vistas como ladras em grande parte da Europa e como presságio de guerra na Escandinávia.[207]
Conservação

Embora a presença humana tenha facilitado a expansão de algumas espécies, como a andorinha-das-chaminés ou o estorninho-comum, foi também a causa da extinção ou diminuição da população de muitas outras espécies. Embora em tempos históricos tenham sido extintas mais de cem espécies de aves,[208] a mais dramática das extinções de aves causada pelo ser humano, que erradicou entre 750 e 1 800 espécies, ocorreu durante a colonização humana das ilhas da Melanésia, Polinésia e Micronésia.[209] Muitas populações de aves em todo o mundo encontram-se em declínio, com 1 375 espécies listadas como ameaçadas pela BirdLife International e pela União Internacional para a Conservação da Natureza e dos Recursos Naturais em 2015.[210]
A ameaça humana às aves mais comum é a destruição dos habitats.[211] Entre as outras ameaças estão a caça em excesso, mortalidade acidental devido a colisões aéreas ou captura acessória na pesca,[212] poluição (incluindo marés negras e uso de pesticidas),[213][214] e competição e predação de espécies invasoras.[215] Os governos e os conservacionistas trabalham em conjunto para proteger as aves, criando legislação que promove a conservação ou o restauro dos habitats ou criando populações de cativeiro para futura reintrodução nos habitats. Um estudo estimou que o esforço de conservação salvou da extinção 16 espécies de aves entre 1994 e 2004, como o condor-da-califórnia ou o periquito-de-Norfolk.[216]
Evolução e classificação

A primeira classificação taxonómica das aves foi publicada por Francis Willughby e John Ray em 1676 no volume Ornithologiae.[217] Em 1758, Carolus Linnaeus modificou essa classificação para o sistema de classificação taxonómica em uso na atualidade.[218] No Sistema de Linné, as aves estão categorizadas na classe homónima das Aves. A classificação filogenética coloca as aves no clado dos dinossauros terópodes.[219] As Aves, bem como o grupo relacionado do clado Crocodilia, são os únicos representantes vivos do clado réptil dos Archosauria. Em termos filogenéticos, as Aves são geralmente definidas como todos os descendentes do ancestral comum mais recente das aves modernas e do Archaeopteryx lithographica.[220] Segundo esta definição, a ave mais antiga que se conhece é o Archaeopteryx, que viveu na idade do Kimmeridgiano do Jurássico Superior, há 155-150 milhões de anos.[221]
No entanto, outros paleontólogos, entre os quais Jacques Gauthier e os aderentes do sistema PhyloCode, definiram Aves de modo a incluir apenas os grupos de aves modernas, excluíndo a maior parte dos grupos conhecidos apenas a partir de fósseis, os quais atribuíram ao grupo Avialae.[221] Segundo esta definição, o Archaeopteryx é um membro dos Avialae, e não das Aves. As propostas de Gauthier têm sido adotadas por muitos investigadores no campo da paleontologia e evolução das aves, embora as definições aplicadas não tenham sido consistentes. O clado Avialae, inicialmente proposto para substituir o conteúdo físsil do grupo das Aves, é muitas vezes usado por estes investigadores como sinónimo do termo vernacular "ave".[222]
Dinossauros e a origem das aves
| ||||||||||||||||||||||||||||||
| Cladograma segundo o estudo filogenético de Godefroit et al., 2013.[222] |
Com base em evidências fósseis e biológicas, a maior parte dos cientistas aceita que as aves são um subgrupo especializado dos dinossauros terópodes,[223] e, mais especificamente, que são membros dos Maniraptora, um grupo de terópodes que inclui os dromeossaurídeos e os oviraptorídeos, entre outros.[224] À medida que os cientistas foram descobrindo mais terópodes estreitamente relacionados com as aves, a distinção anteriormente clara entre aves e não aves foi-se esbatendo. As descobertas recentes na província chinesa de Liaoning de vários dinossauros emplumados terópodes de pequena dimensão vieram contribuir para esta ambiguidade.[225][226][227]
O consenso na paleontologia contemporânea é de que os terópodes voadores, ou Avialae, são os parentes mais próximos dos deinonicossauros, grupo que inclui os dromeossaurídeos e os troodontídeos.[228] Em conjunto, estes formam um grupo designado Paraves. Alguns membros basais deste grupo, como o Microraptor, apresentam características que lhes podem ter permitido planar ou voar. A maior parte dos deinocossauros basais eram muito pequenos. Estas evidências aumentam a possibilidade de que o ancestral de todos os paraves possa ter sido arborícola, ter sido capaz de planar, ou ambos.[229][230] Ao contrário do Archaeopteryx e dos dinossauros emplumados não avianos, que comiam principalmente carne, os estudos mais recentes sugerem que os primeiros avianos eram omnívoros.[231]
O Archaeopteryx, que viveu no Jurássico Superior, foi um dos primeiros fósseis de transição descobertos, tendo sido uma das bases para a teoria da evolução em finais do século XIX. O Archaeopteryx foi o primeiro fóssil a exibir não só características claramente de réptil, como os dentes, garras e uma cauda comprida semelhante a um lagarto, mas também asas com penas adaptadas ao voo semelhantes às das aves modernas. No entanto, não é considerado um ancestral direto das aves, embora esteja possivelmente relacionado com o verdadeiro ancestral.[232]
Evolução inicial
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Cladograma segundo os resultados do estudo filogenético de Lefèvre et al., 2014.[233] |
Os mais antigos fósseis avianos conhecidos são provenientes da Formação de Tiaojishan, na China, e estão datados do estágio Oxfordiano do período Jurássico Superior, há cerca de 160 milhões de anos.[222] As espécies avianas deste período incluem o Anchiornis huxleyi, Xiaotingia zhengi e o Aurornis xui. O aviano mais conhecido, Archaeopteryx, data de rochas jurássicas alemãs ligeiramente posteriores, de há cerca de 155 milhões de anos. Muitos destes primeiros avianos partilhavam entre si características anatómicas invulgares que podem ter sido ancestrais das aves modernas, mas que foram mais tarde perdidas durante a evolução das aves. Estas características incluem uma garra maior no segundo dedo e asas posteriores que cobriam os membros e pés posteriores e que podem ter sido usadas para manobras no ar.[234]
Durante o Cretácico, os avianos diversificaram-se numa ampla variedade de formas.[235] Muitos grupos mantiveram as características primitivas, como as asas com garras e os dentes. Estes últimos acabaram por se perder em vários grupos avianos, incluindo as aves modernas, embora de forma independente entre si. Enquanto os avianos mais antigos, como o Archaeopteryx e o Shenzhouraptor sinensis, mantiveram as caudas ósseas dos seus ancestrais,[235] as caudas de avianos mais avançados foram sendo encurtadas após o aparecimento dos ossos pigóstilos no grupo Pygostylia. No Cretácico superior, há cerca de 95 milhões de anos, o ancestral de todas as aves modernas desenvolveu também o sentido do olfato.[236]
Diversificação inicial dos ancestrais das aves
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Filogenia das aves durante o Mesozoico, simplificada de acordo com a análise filogenética de Wang et al de 2015.[237] |
A primeira grande e diversificada linhagem de avianos de cauda curta a evoluir foram os enantiornithes, ou "opostos às aves", assim designados porque a construção dos ossos dos ombros se apresenta invertida em relação às aves modernas. Os enantiornithes ocupavam um vasto conjunto de nichos ecológicos, desde os que vasculhavam as areias nas zonas costeiras, passando pelos que se alimentavam de peixe, até aos que viviam nas árvores e se alimentavam de sementes. Embora tenham sido o grupo dominante dentro dos avianos durante o período Cretácico, os enantiornithes extinguiram-se no fim do Mesozoico, a par de muitos outros grupos de dinossauros.[235]
A segunda grande linhagem aviana a desenvolver-se foram os Euornithes, ou "aves verdadeiras", assim designados porque incluem os ancestrais das aves modernas. Muitas das espécies deste grupo eram semi-aquáticas e especializadas na captura de peixe e outros organismos marinhos. Ao contrário dos enantiornithes, que dominavam os habitats terrestres e arbustivos, à maior parte dos primeiros euornithes faltavam adaptações que lhes permitissem empoleirar, pelo que neste grupo se observam sobretudo espécies adaptadas às regiões costeiras ou capazes de nadar e mergulhar. Entre as que possuíam esta última característica estão os hesperornithiformes, que se tornaram de tal forma especializados na pesca de peixe em ambientes marinhos que perderam a capacidade de voar e se tornaram aquáticos.[235] Os primeiros euornithes também assistiram ao desenvolvimento de várias características associadas às aves modernas, como osso esterno com quilha, ausência de dentes e partes da mandíbula em forma de bico. No entanto, a maior parte dos euornithes manteve dentes noutras partes da mandíbula.[238] O grupo dos euornithes também inclui os primeiros avianos a desenvolver um verdadeiro pigóstilo e um conjunto completo de penas de cauda,[239] o qual pode ter substituído as asas posteriores como principal meio de manobrabilidade e desaceleração em voo.[234]
Diversificação das aves modernas
| |||||||||||||||||||||||||||
| Divergências basais das aves modernas com base na Taxonomia de Sibley-Ahlquist |
Todas as aves modernas pertencem ao grupo coroa das Aves (ou, em alternativa, Neornithes), o qual tem duas subdivisões: os Paleognathae, em que se inclui os ratitas não voadores (como a avestruz), os tinamiformes e o extremamente diversificado grupo dos Neognathae, que contém todas as outras aves.[219] A estas duas subdivisões dá-se muitas vezes a categoria taxonómica de superordem.[219][240] Dependendo do ponto de vista taxonómico, o número de espécies de aves vivas conhecidas varia entre 9 800[241] e 10 050.[242] Devido em grande parte à descoberta do Vegavis, um membro neognato da linhagem dos patos, sabe-se que o grupo Aves se dividiu em várias linhagens modernas por volta do fim da era do Mesozoico.[243] Alguns estudos estimaram que a origem efetiva das aves modernas terá provavelmente ocorrido mais cedo do que os fósseis conhecidos mais antigos, provavelmente a meio do período Cretácico.[4]
Há um consenso de que as Aves se desenvolveram durante o Cretácico e que a divisão dos Galloanseri dos outros neognatos ocorreu antes da extinção do Cretáceo-Paleogeno, embora haja diferentes opiniões sobre se a radiação evolutiva dos restantes neognatos ocorreu antes ou depois da extinção dos restantes dinossauros.[244] Esta discordância deve-se em parte a uma divergência nas evidências. Os dados moleculares sugerem uma radiação durante o Cretácico, enquanto as evidências fósseis sugerem uma radiação durante o Cenozoico. As tentativas para reconciliar as evidências moleculares e fósseis têm-se revelado controversas,[244][245] embora resultados recentes mostrem que todos os grupos existentes de aves tenham tido origem num pequeno grupo de espécies que sobreviveu à extinção do Cretáceo-Paleogeno.[246]
Classificação das ordens das aves
Cladograma das relações entre aves modernas com base em Jarvis, E.D. et al. (2014),[247] com alguns nomes de clados segundo Yury, T. et al. (2013).[248]















A classificação das aves é um tema controverso. A publicação Phylogeny and Classification of Birds (1990), da autoria de Charles Sibley e Jon Ahlquist, é uma das principais obras de referência na classificação das aves,[249] embora seja frequentementemente discutida e continuamente revista. A maior parte das evidências sugere que a distribuição das ordens é correta,[250] embora os cientistas discordem entre si quanto às relações entre as ordens. Apesar de a discussão contar com evidências da anatomia das aves modernas, ADN e registos fósseis, ainda não há um consenso determinante. Recentemente, a descoberta de novos fósseis e a obtenção de evidências moleculares está a contribuir para uma panorâmica cada vez mais clara da evolução das ordens de aves modernas. A investigação mais recente tem-se focado na sequenciação de genomas completos de 48 espécies representativas.[247]
Ver também
- ↑ Embora também tenham estado presentes em alguns dinossauros, hoje em dia não são considerados verdadeiras Aves.
- ↑ De onde deriva a palavra "auspício"
- ↑ Brown, J.W. & Van Tuinen, M. (2011). «Evolving Perceptions on the Antiquity of the Modern Avian Tree, in Living Dinosaurs». John Wiley & Sons LtD. The Evolutionary History of Modern Birds: 306–324. doi:10.1002/9781119990475.ch12
- ↑ «Asteroid impact helped create the birds we know today». Science. Consultado em 12 de fevereiro de 2016
- ↑ «Influence of Earth's history on the dawn of modern birds». www.sciencedaily.com. American Museum of Natural History. 11 de dezembro de 2015. Consultado em 11 de dezembro de 2015
- ↑ a b c Lee, Michael SY; Cau, Andrea; Darren, Naish; Gareth J., Dyke (2013). «Morphological Clocks in Paleontology, and a Mid-Cretaceous Origin of Crown Aves». Oxford Journals. Systematic Biology. 63: 442–449. PMID 24449041. doi:10.1093/sysbio/syt110
- ↑ Alonso, P. D.; Milner, A. C.; Ketcham, R. A.; Cookson, M. J.; Rowe, T. B. (2004). «The avian nature of the brain and inner ear of Archaeopteryx» (PDF). Nature. 430 (7000): 666–669. PMID 15295597. doi:10.1038/nature02706
- ↑ a b c d e f g h i j k l m n o p q r Storer, Robert W. (2016). «Bird». Enciclopédia Britannica. Consultado em 6 de março de 2016
- ↑ Brooke, Michael (2004). Albatrosses And Petrels Across The World. Oxford: Oxford University Press. ISBN 0-19-850125-0
- ↑ Weir, Jason T.; Schluter, D (2007). «The Latitudinal Gradient in Recent Speciation and Extinction Rates of Birds and Mammals». Science. 315 (5818): 1574–76. PMID 17363673. doi:10.1126/science.1135590
- ↑ a b Schreiber, Elizabeth Anne; Joanna Burger (2001). Biology of Marine Birds. Boca Raton: CRC Press. ISBN 0-8493-9882-7
- ↑ Sato, Katsufumi; Y Naito, A Kato, Y Niizuma, Y Watanuki, JB Charrassin, CA Bost, Y Handrich, Y Le Maho (1 de maio de 2002). «Buoyancy and maximal diving depth in penguins: do they control inhaling air volume?». Journal of Experimental Biology. 205 (9): 1189–1197. PMID 11948196
- ↑ Hill, David; Peter Robertson (1988). The Pheasant: Ecology, Management, and Conservation. Oxford: BSP Professional. ISBN 0-632-02011-3
- ↑ Spreyer, Mark F.; Enrique H. Bucher (1998). «Monk Parakeet (Myiopsitta monachus), The Birds of North America». Cornell Lab of Ornithology. doi:10.2173/bna.322. Consultado em 13 de dezembro de 2015
- ↑ Arendt, Wayne J. (1 de janeiro de 1988). «Range Expansion of the Cattle Egret, (Bubulcus ibis) in the Greater Caribbean Basin». Colonial Waterbirds. 11 (2): 252–62. JSTOR 1521007. doi:10.2307/1521007
- ↑ Bierregaard, R.O. (1994). «Yellow-headed Caracara». In: Josep del Hoyo, Andrew Elliott and Jordi Sargatal (eds.). Handbook of the Birds of the World. Volume 2; New World Vultures to Guineafowl. Barcelona: Lynx Edicions. ISBN 84-87334-15-6
- ↑ Juniper, Tony; Mike Parr (1998). Parrots: A Guide to the Parrots of the World. London: Christopher Helm. ISBN 0-7136-6933-0
- ↑ a b c d e f g h i j k l m n o p q r s t u v w x y Gill, Frank (1995). Ornithology. Nova Iorque: WH Freeman and Co. ISBN 0-7167-2415-4
- ↑ Roots, Clive (2006). Flightless Birds. Westport: Greenwood Press. ISBN 978-0-313-33545-7
- ↑ McNab, Brian K. (Outubro de 1994). «Energy Conservation and the Evolution of Flightlessness in Birds». The American Naturalist. 144 (4): 628–42. JSTOR 2462941. doi:10.1086/285697
- ↑ Kovacs, Christopher E.; Meyers, RA (2000). «Anatomy and histochemistry of flight muscles in a wing-propelled diving bird, the Atlantic Puffin, Fratercula arctica». Journal of Morphology. 244 (2): 109–25. PMID 10761049. doi:10.1002/(SICI)1097-4687(200005)244:2<109::AID-JMOR2>3.0.CO;2-0
- ↑ Belthoff, James R.; Dufty,; Gauthreaux, (1 de agosto de 1994). «Plumage Variation, Plasma Steroids and Social Dominance in Male House Finches». The Condor. 96 (3): 614–25. doi:10.2307/1369464
- ↑ Guthrie, R. Dale. «How We Use and Show Our Social Organs». Body Hot Spots: The Anatomy of Human Social Organs and Behavior. Consultado em 19 de outubro de 2007. Cópia arquivada em 21 de junho de 2007
- ↑ Ginn, HB; Melville, DS (1983). Moult in birds. [S.l.]: British Trusl for Ornilology, Tring Field Guide. pp. 16–21. ISBN 978-0903793025
- ↑ a b c Pettingill Jr. OS (1970). Ornithology in Laboratory and Field. [S.l.]: Burgess Publishing Co. ISBN 0-12-552455-2
- ↑ de Beer, SJ; Lockwood GM, Raijmakers JHFS, Raijmakers JMH, Scott WA, Oschadleus HD, Underhill LG (2001). SAFRING Bird Ringing Manual (PDF). Col: ADU Guide. [S.l.]: Universidade da Cidade do Cabo
- ↑ Gargallo, Gabriel (1 de junho de 1994). «Flight Feather Moult in the Red-Necked Nightjar Caprimulgus ruficollis». Journal of Avian Biology. 25 (2): 119–24. JSTOR 3677029. doi:10.2307/3677029
- ↑ Mayr, Ernst (1954). «The tail molt of small owls» (PDF). The Auk. 71 (2): 172–78. doi:10.2307/4081571
- ↑ Lucas, Alfred M. (1972). Avian Anatomy—integument. East Lansing, Michigan, US: USDA Avian Anatomy Project, Michigan State University. pp. 67, 344, 394–601
- ↑ a b Proctor, Noble S.; Lynch, Patrick J. (1998). Manual of Ornithology: Avian Structure and Function. New Haven: Yale University Press. p. 66. ISBN 0-300-07619-3
- ↑ Samour (2000), p. 296.
- ↑ Girling (2003), p. 4.
- ↑ Bonser RHC & Mark S Witter (1993). «Indentation hardness of the bill keratin of the European Starling» (PDF). The Condor. 95: 736–738. doi:10.2307/1369622
- ↑ Campbell and Lack (1985), p. 598.
- ↑ Stettenheim, Peter R. «The Integumentary Morphology of Modern Birds—An Overview» (PDF). Integrative and Comparative Biology. 40 (4): 461–477. doi:10.1093/icb/40.4.461
- ↑ Klasing, Kirk C. (1999). «Avian gastrointestinal anatomy and physiology». Seminars in Avian and Exotic Pet Medicine. 8 (2): 42–50. doi:10.1016/S1055-937X(99)80036-X
- ↑ Ferguson-Lees, James; Christie, David A. Raptors of the World. Londres: Christopher Helm. p. 66. ISBN 0-7136-8026-1
- ↑ Campbell and Lack (1985), p. 48.
- ↑ Gosner, Kenneth L. (Junho de 1993). «Scopate Tomia: An Adaptation for Handling Hard-shelled Prey?» (PDF). The Wilson Bulletin. 105 (2): 316–324
- ↑ Wilkie, Susan E.; Vissers, PM; Das, D; Degrip, WJ; Bowmaker, JK; Hunt, DM (Fevereiro de 1998). «The molecular basis for UV vision in birds: spectral characteristics, cDNA sequence and retinal localization of the UV-sensitive visual pigment of the budgerigar (Melopsittacus undulatus)». Biochemical Journal. 330 (Pt 1): 541–47. PMC 1219171
. PMID 9461554
- ↑ Andersson, S.; J. Ornborg, M. Andersson (1998). «Ultraviolet sexual dimorphism and assortative mating in blue tits». Proceedings of the Royal Society B. 265 (1395): 445–50. doi:10.1098/rspb.1998.0315
- ↑ Viitala, Jussi; Korplmäki, Erkki; Palokangas, Pälvl; Koivula, Minna (1995). «Attraction of kestrels to vole scent marks visible in ultraviolet light». Nature. 373 (6513): 425–27. doi:10.1038/373425a0
- ↑ Williams, David L.; Flach, E (Março de 2003). «Symblepharon with aberrant protrusion of the nictitating membrane in the snowy owl (Nyctea scandiaca)». Veterinary Ophthalmology. 6 (1): 11–13. PMID 12641836. doi:10.1046/j.1463-5224.2003.00250.x
- ↑ White, Craig R.; Day, N; Butler, PJ; Martin, GR; Bennett, Peter (Julho 2007). Bennett, Peter, ed. «Vision and Foraging in Cormorants: More like Herons than Hawks?». PLoS ONE. 2 (7): e639. PMC 1919429
- ↑ Martin, Graham R.; Katzir, G (1999). «Visual fields in Short-toed Eagles, Circaetus gallicus (Accipitridae), and the function of binocularity in birds». Brain, Behaviour and Evolution. 53 (2): 55–66. PMID 9933782. doi:10.1159/000006582
- ↑ Saito, Nozomu (1978). «Physiology and anatomy of avian ear». The Journal of the Acoustical Society of America. 64 (S1): S3. doi:10.1121/1.2004193
- ↑ Whittow, G. Causey (2000). Sturkie's Avian Physiology. [S.l.]: Academic Press. pp. 233–241. ISBN 978-0-12-747605-6
- ↑ Calder, William A. (1996). Size, Function, and Life History. Mineola, Nova Iorque: Courier Dove Publications. p. 91. ISBN 978-0-486-69191-6
- ↑ Maina, John N. (Novembro de 2006). «Development, structure, and function of a novel respiratory organ, the lung-air sac system of birds: to go where no other vertebrate has gone». Biological Reviews. 81 (4): 545–79. PMID 17038201. doi:10.1017/S1464793106007111
- ↑ a b Suthers, Roderick A.; Sue Anne Zollinger (2004). «Producing song: the vocal apparatus». In: H. Philip Zeigler and Peter Marler (eds.). Behavioral Neurobiology of Birdsong. Col: Annals of the New York Academy of Sciences 1016. Nova Iorque: New York Academy of Sciences. pp. 109–129. ISBN 1-57331-473-0. PMID 15313772. doi:10.1196/annals.1298.041
- ↑ Fitch, W. T. (1999). «Acoustic exaggeration of size in birds via tracheal elongation: comparative and theoretical analyses». Journal of Zoology. 248: 31–48. doi:10.1017/S095283699900504X
- ↑ Sales, James (2005). «The endangered kiwi: a review» (PDF). Folia Zoologica. 54 (1–2): 1–20
- ↑ Ehrlich, Paul R.; David S. Dobkin; Darryl Wheye (1988). «The Avian Sense of Smell». Birds of Stanford. Stanford University. Consultado em 13 de dezembro de 2007
- ↑ Lequette, Benoit; Verheyden, Jouventin (1 de agosto 1989). «Olfaction in Subantarctic seabirds: Its phylogenetic and ecological significance» (PDF). The Condor. 91 (3): 732–35. doi:10.2307/1368131. Cópia arquivada em 23 de fevereiro de 2013
- ↑ Campbell, Bruce; Lack, Elizabeth (1985). A Dictionary of Birds. Carlton: T and A D Poyser. pp. 47, 375. ISBN 0-85661-039-9
- ↑ Ehrlich, Paul R.; David S. Dobkin; Darryl Wheye (1988). «Adaptations for Flight». Birds of Stanford. Stanford University. Consultado em 13 de dezembro de 2007 com base em: Paul Ehrlich, David Dobkin, e Darryl Wheye (1988). The Birder's Handbook. Nova Iorque: Simon and Schuster
- ↑ Noll, Paul. «The Avian Skeleton». paulnoll.com. Consultado em 13 de dezembro de 2007
- ↑ «Skeleton of a typical bird». Fernbank Science Center's Ornithology Web. Consultado em 13 de dezembro de 2007
- ↑ Romão, Ricardo. «Osteologia das Aves» (PDF). Universidade de Évora – Departamento de Zoologia. Consultado em 15 de fevereiro de 2016
- ↑ Gionfriddo, James P. (1 Fevereiro de 1995). «Grit Use by House Sparrows: Effects of Diet and Grit Size» (PDF). Condor. 97 (1): 57–67. doi:10.2307/1368983
- ↑ a b c Attenborough, David (1998). The Life of Birds. Princeton: Princeton University Press. ISBN 0-691-01633-X
- ↑ a b Battley, Phil F.; Piersma,T; Dietz, MW; Tang, S; Dekinga, A; Hulsman, K (Janeiro de 2000). «Empirical evidence for differential organ reductions during trans-oceanic bird flight». Proceedings of the Royal Society B. 267 (1439): 191–5. PMC 1690512
- ↑ a b Balgooyen, Thomas G. (1 de outubro de 1971). «Pellet Regurgitation by Captive Sparrow Hawks (Falco sparverius)» (PDF). Condor. 73 (3): 382–85. JSTOR 1365774. doi:10.2307/1365774. Cópia arquivada em 24 de maio de 2013
- ↑ Engel, Sophia Barbara (2005). Racing the wind: Water economy and energy expenditure in avian endurance flight. [S.l.]: University of Groningen. ISBN 90-367-2378-7
- ↑ Tieleman, B.I.; Williams, JB (1999). «The role of hyperthermia in the water economy of desert birds». Physiol. Biochem. Zool. 72 (1): 87–100. PMID 9882607. doi:10.1086/316640
- ↑ Schmidt-Nielsen, Knut (1 de maio de 1960). «The Salt-Secreting Gland of Marine Birds». Circulation. 21 (5): 955–967. doi:10.1161/01.CIR.21.5.955
- ↑ Hallager, Sara L. (1994). «Drinking methods in two species of bustards». Wilson Bull. 106 (4): 763–764
- ↑ MacLean, Gordon L. (1 de junho de 1983). «Water Transport by Sandgrouse». BioScience. 33 (6): 365–369. JSTOR 1309104. doi:10.2307/1309104
- ↑ Eraud C; Dorie A, Jacquet A, Faivre B (2008). «The crop milk: a potential new route for carotenoid-mediated parental effects». Journal of Avian Biology. 39 (2): 247–251. doi:10.1111/j.0908-8857.2008.04053.x
- ↑ Ehrlich, Paul R.; David S. Dobkin; Darryl Wheye (1988). «Drinking». Birds of Stanford. Stanford University. Consultado em 13 de dezembro de 2007
- ↑ Tsahar, Ella; Martínez Del Rio, C; Izhaki, I; Arad, Z (2005). «Can birds be ammonotelic? Nitrogen balance and excretion in two frugivores». Journal of Experimental Biology. 208 (6): 1025–34. PMID 15767304. doi:10.1242/jeb.01495
- ↑ Skadhauge, E; Erlwanger, KH; Ruziwa, SD; Dantzer, V; Elbrønd, VS; Chamunorwa, JP (2003). «Does the ostrich (Struthio camelus) coprodeum have the electrophysiological properties and microstructure of other birds?». Comparative biochemistry and physiology. Part A, Molecular & integrative physiology. 134 (4): 749–755. PMID 12814783. doi:10.1016/S1095-6433(03)00006-0
- ↑ Preest, Marion R.; Beuchat, Carol A. (Abril 1997). «Ammonia excretion by hummingbirds». Nature. 386 (6625): 561–62. doi:10.1038/386561a0
- ↑ Mora, J.; Martuscelli, J; Ortiz Pineda, J; Soberon, G (1965). «The regulation of urea-biosynthesis enzymes in vertebrates» (PDF). en:Biochemical Journal. 96 (1): 28–35. PMC 1206904
- ↑ Packard, Gary C. (1966). «The Influence of Ambient Temperature and Aridity on Modes of Reproduction and Excretion of Amniote Vertebrates». en:The American Naturalist. 100 (916): 667–82. JSTOR 2459303. doi:10.1086/282459
- ↑ Deviche, Pierre; Hurley, Laura L; Fokidis, H Bobby (2010). «Chapter 2 - Avian Testicular Structure, Function, and Regulation». In: David Norris, Kristin Lopez. Hormones and Reproduction of Vertebrates. [S.l.]: Academic Press. pp. 27–70. ISBN 9780123749321. doi:10.1016/B978-0-12-374929-1.10002-2
- ↑ Kinsky, FC (1971). «The consistent presence of paired ovaries in the Kiwi(Apteryx) with some discussion of this condition in other birds». Journal of Ornithology. 112 (3): 334–357. doi:10.1007/BF01640692
- ↑ Yong, Ed. «Phenomena: Not Exactly Rocket Science How Chickens Lost Their Penises (And Ducks Kept Theirs)». Phenomena.nationalgeographic.com. Consultado em 3 de outubro de 2013
- ↑ «Ornithology, 3rd Edition - Waterfowl: Order Anseriformes». Consultado em 3 de outubro de 2013
- ↑ McCracken, KG (2000). «The 20-cm Spiny Penis of the Argentine Lake Duck (Oxyura vittata)» (PDF). The Auk. 117 (3): 820–825. doi:10.1642/0004-8038(2000)117[0820:TCSPOT]2.0.CO;2. Cópia arquivada em 24 de maio de 2013
- ↑ a b Whittow, G. (2000). G. Causey Whittow, ed. Sturkie's Avian Physiology. San Diego: Academic Press
- ↑ a b Hoagstrom, C.W. (2002). Vertebrate Circulation. Magill's Encyclopedia of Science: Animal Life. 1. Pasadena, Califórnia: Salem Press. pp. 217–219
- ↑ Scott, Robert B. (Março de 1966). «Comparative hematology: The phylogeny of the erythrocyte». Annals of Hematology. 12 (6): 340–51. PMID 5325853. doi:10.1007/BF01632827
- ↑ Hill, Richard W.; Gordon A. Wyse, Margaret Anderson. (2012). Animal Physiology 3ª ed. Sunderland: Sinauer Associates. pp. 647–678
- ↑ Warham, John (1 de maio de 1977). «The incidence, function and ecological significance of petrel stomach oils» (PDF). Proceedings of the New Zealand Ecological Society. 24 (3): 84–93
- ↑ Dumbacher, J.P.; Beehler, BM; Spande, TF; Garraffo, HM; Daly, JW (Outubro de 1992). «Homobatrachotoxin in the genus Pitohui: chemical defense in birds?». Science. 258 (5083): 799–801. PMID 1439786. doi:10.1126/science.1439786
- ↑ Thomas, Betsy Trent; Strahl (1 de agosto de 1990). «Nesting Behavior of Sunbitterns (Eurypyga helias) in Venezuela» (PDF). The Condor. 92 (3): 576–81. doi:10.2307/1368675. Cópia arquivada em 24 de maio de 2013
- ↑ a b c Longrich, N. R.; Olson, S. L. (5 de janeiro de 2011). «The bizarre wing of the Jamaican flightless ibis Xenicibis xympithecus: a unique vertebrate adaptation». Proceedings of the Royal Society B: Biological Sciences. 278 (1716): 2333–2337. doi:10.1098/rspb.2010.2117
- ↑ Robert, Michel; McNeil, Raymond; Leduc, Alain (Janeiro de 1989). «Conditions and significance of night feeding in shorebirds and other water birds in a tropical lagoon» (PDF). The Auk. 106 (1): 94–101. doi:10.2307/4087761. Cópia arquivada em 24 de maio de 2013
- ↑ a b N Reid (2006). «Birds on New England wool properties – A woolgrower guide» (PDF). Land, Water & Wool Northern Tablelands Property Fact Sheet. Australian Government – Land and Water Australia. Consultado em 17 de julho de 2010. Cópia arquivada (PDF) em 15 de março de 2011
- ↑ Paton, D. C.; Collins, B. G. (1 Abril 1989). «Bills and tongues of nectar-feeding birds: A review of morphology, function, and performance, with intercontinental comparisons». Australian Journal of Ecology. 14 (4): 473–506. doi:10.1111/j.1442-9993.1989.tb01457.x
- ↑ Baker, Myron Charles; Baker, Ann Eileen Miller (1 de abril de 1973). «Niche Relationships Among Six Species of Shorebirds on Their Wintering and Breeding Ranges». Ecological Monographs. 43 (2): 193–212. JSTOR 1942194. doi:10.2307/1942194
- ↑ Cherel, Yves; Bocher, P; De Broyer, C; Hobson, KA (2002). «Food and feeding ecology of the sympatric thin-billed Pachyptila belcheri and Antarctic P. desolata prions at Iles Kerguelen, Southern Indian Ocean». Marine Ecology Progress Series. 228: 263–81. doi:10.3354/meps228263
- ↑ Jenkin, Penelope M. (1957). «The Filter-Feeding and Food of Flamingoes (Phoenicopteri)». Philosophical Transactions of the Royal Society B. 240 (674): 401–93. JSTOR 92549. doi:10.1098/rstb.1957.0004
- ↑ Miyazaki, Masamine; Kuroki, M; Niizuma, Y; Watanuki, Y. (1 Julho 1996). «Vegetation cover, kleptoparasitism by diurnal gulls and timing of arrival of nocturnal Rhinoceros Auklets» (PDF). The Auk. 113 (3): 698–702. JSTOR 3677021. doi:10.2307/3677021. Cópia arquivada em 24 de maio de 2013
- ↑ Bélisle, Marc; Giroux (1 de agosto de 1995). «Predation and kleptoparasitism by migrating Parasitic Jaegers» (PDF). The Condor. 97 (3): 771–781. doi:10.2307/1369185
- ↑ Vickery, J. A.; Brooke, M. De L. (1 de maio de 1994). «The Kleptoparasitic Interactions between Great Frigatebirds and Masked Boobies on Henderson Island, South Pacific» (PDF). The Condor. 96 (2): 331–40. JSTOR 1369318. doi:10.2307/1369318. Cópia arquivada em 24 de maio de 2013
- ↑ Hiraldo, F.C. (1991). «Unspecialized exploitation of small carcasses by birds». Bird Studies. 38 (3): 200–07. doi:10.1080/00063659109477089 Parâmetro desconhecido
|Coautor=ignorado (|coautor=) sugerido (ajuda) - ↑ Mario, Principato; Lisi Federica, Moretta Iolanda, Samra Nada, Puccetti Francesco (2005). «The alterations of plumage of parasitic origin». Italian Journal of Animal Science. 4: 296–299
- ↑ Ehrlich, Paul R. (1986). «The Adaptive Significance of Anting» (PDF). The Auk. 103 (4). 835 páginas. Cópia arquivada em 24 de maio de 2013
- ↑ Revis, Hannah C.; Waller, Deborah A. (2004). «Bactericidal and fungicidal activity of ant chemicals on feather parasites: an evaluation of anting behavior as a method of self-medication in songbirds». The Auk. 121 (4): 1262–1268. doi:10.1642/0004-8038(2004)121[1262:BAFAOA]2.0.CO;2
- ↑ Clayton, Dale H.; Koop, Jennifer A. H.; Harbison, Christopher W.; Moyer, Brett R.; Bush, Sarah E. (2010). «How Birds Combat Ectoparasites» (PDF). The Open Ornithology Journal. 3: 41–71. doi:10.2174/1874453201003010041
- ↑ Walther, Bruno A. (2005). «Elaborate ornaments are costly to maintain: evidence for high maintenance handicaps». Behavioural Ecology. 16 (1): 89–95. doi:10.1093/beheco/arh135
- ↑ Shawkey, Matthew D.; Pillai, Shreekumar R.; Hill, Geoffrey E. (2003). «Chemical warfare? Effects of uropygial oil on feather-degrading bacteria». jornal of Avian Biology. 34 (4): 345–49. doi:10.1111/j.0908-8857.2003.03193.x
- ↑ Klaassen, Marc (1 Janeiro 1996). «Metabolic constraints on long-distance migration in birds». Journal of Experimental Biology. 199 (1): 57–64. PMID 9317335
- ↑ Gill, Frank (1995). Ornithology 2ª ed. Nova Iorque: W.H. Freeman. ISBN 0-7167-2415-4
- ↑ «Long-distance Godwit sets new record». BirdLife International. 4 de maio de 2007. Consultado em 13 de dezembro de 2007
- ↑ Shaffer, Scott A.; Tremblay, Y; Weimerskirch, H; Scott, D; Thompson, DR; Sagar, PM; Moller, H; Taylor, GA; Foley, DG (2006). «Migratory shearwaters integrate oceanic resources across the Pacific Ocean in an endless summer». Proceedings of the National Academy of Sciences of the United States of America. 103 (34): 12799–802. PMC 1568927
- ↑ Croxall, John P.; Silk, JR; Phillips, RA; Afanasyev, V; Briggs, DR (2005). «Global Circumnavigations: Tracking year-round ranges of nonbreeding Albatrosses». Science. 307 (5707): 249–50. PMID 15653503. doi:10.1126/science.1106042
- ↑ Wilson, W. Herbert, Jr. (1999). «Bird feeding and irruptions of northern finches:are migrations short stopped?» (PDF). North America Bird Bander. 24 (4): 113–21. Cópia arquivada em 24 de maio de 2013
- ↑ Nilsson, Anna L. K.; Alerstam, Thomas; Nilsson, Jan-Åke (2006). «Do partial and regular migrants differ in their responses to weather?». The Auk. 123 (2): 537–47. doi:10.1642/0004-8038(2006)123[537:DPARMD]2.0.CO;2
- ↑ Chan, Ken (2001). «Partial migration in Australian landbirds: a review». Emu. 101 (4): 281–92. doi:10.1071/MU00034
- ↑ Rabenold, Kerry N. (1985). «Variation in Altitudinal Migration, Winter Segregation, and Site Tenacity in two subspecies of Dark-eyed Juncos in the southern Appalachians» (PDF). The Auk. 102 (4): 805–19
- ↑ Collar, Nigel J. (1997). «Family Psittacidae (Parrots)». In: Josep del Hoyo, Andrew Elliott, Jordi Sargatal. Handbook of the Birds of the World, Volume 4: Sandgrouse to Cuckoos. Barcelona: Lynx Edicions. ISBN 84-87334-22-9
- ↑ Matthews, G. V. T. (1 setembro 1953). «Navigation in the Manx Shearwater». Journal of Experimental Biology. 30 (2): 370–96
- ↑ Mouritsen, Henrik (15 de novembro de 2001). «Migrating songbirds tested in computer-controlled Emlen funnels use stellar cues for a time-independent compass». Journal of Experimental Biology. 204 (8): 3855–65. PMID 11807103
- ↑ Deutschlander, Mark E. (15 de abril de 1999). «The case for light-dependent magnetic orientation in animals». Journal of Experimental Biology. 202 (8): 891–908. PMID 10085262
- ↑ Möller, Anders Pape (1988). «Badge size in the house sparrow Passer domesticus». Behavioral Ecology and Sociobiology. 22 (5): 373–78. doi:10.1007/BF00295107
- ↑ Thomas, Betsy Trent; Strahl (1 de agosto de 1990). «Nesting Behavior of Sunbitterns (Eurypyga helias) in Venezuela» (PDF). The Condor. 92 (3): 576–81. doi:10.2307/1368675. Cópia arquivada em 24 de maio de 2013
- ↑ Pickering, S. P. C. (2001). «Courtship behaviour of the Wandering Albatross Diomedea exulans at Bird Island, South Georgia» (PDF). Marine Ornithology. 29 (1): 29–37
- ↑ Pruett-Jones, S. G.; Pruett-Jones (1 de maio de 1990). «Sexual Selection Through Female Choice in Lawes' Parotia, A Lek-Mating Bird of Paradise». Evolution. 44 (3): 486–501. doi:10.2307/2409431
- ↑ Genevois, F.; Bretagnolle, V. (1994). «Male Blue Petrels reveal their body mass when calling». Ethology Ecology and Evolution. 6 (3): 377–83. doi:10.1080/08927014.1994.9522988
- ↑ Jouventin, Pierre; Aubin, T; Lengagne, T (Junho de 1999). «Finding a parent in a king penguin colony: the acoustic system of individual recognition». Animal Behaviour. 57 (6): 1175–83. PMID 10373249. doi:10.1006/anbe.1999.1086
- ↑ Templeton, Christopher N.; Greene, E; Davis, K (2005). «Allometry of Alarm Calls: Black-Capped Chickadees Encode Information About Predator Size». Science. 308 (5730): 1934–37. PMID 15976305. doi:10.1126/science.1108841
- ↑ Miskelly, C. M. (Julho de 1987). «The identity of the hakawai». Notornis. 34 (2): 95–116
- ↑ Murphy, Stephen; Legge, Sarah; Heinsohn, Robert (2003). «The breeding biology of palm cockatoos (Probosciger aterrimus): a case of a slow life history». Journal of Zoology. 261 (4): 327–39. doi:10.1017/S0952836903004175
- ↑ a b Sekercioglu, Cagan Hakki (2006). «Foreword». In: Josep del Hoyo, Andrew Elliott and David Christie. Handbook of the Birds of the World. 11: Old World Flycatchers to Old World Warblers. Barcelona: Lynx Edicions. 48 páginas. ISBN 84-96553-06-X
- ↑ Terborgh, John (2005). «Mixed flocks and polyspecific associations: Costs and benefits of mixed groups to birds and monkeys». American Journal of Primatology. 21 (2): 87–100. doi:10.1002/ajp.1350210203
- ↑ Hutto, Richard L. (Janeiro de 1988). «Foraging Behavior Patterns Suggest a Possible Cost Associated with Participation in Mixed-Species Bird Flocks». Oikos. 51 (1): 79–83. JSTOR 3565809. doi:10.2307/3565809
- ↑ Au, David W. K. (1 de agosto de 1986). «Seabird interactions with Dolphins and Tuna in the Eastern Tropical Pacific» (PDF). The Condor. 88 (3): 304–17. doi:10.2307/1368877
- ↑ Anne, O.; Rasa, E. (Junho de 1983). «Dwarf mongoose and hornbill mutualism in the Taru desert, Kenya». Behavioral Ecology and Sociobiology. 12 (3): 181–90. doi:10.1007/BF00290770
- ↑ Gauthier-Clerc, Michael; Tamisier, Alain; Cézilly, Frank (2000). «Sleep-Vigilance Trade-off in Gadwall during the Winter Period» (PDF). The Condor. 102 (2): 307–13. JSTOR 1369642. doi:10.1650/0010-5422(2000)102[0307:SVTOIG]2.0.CO;2. Cópia arquivada (PDF) em 27 de dezembro de 2004
- ↑ Rattenborg, Niels C. (2006). «Do birds sleep in flight?». Die Naturwissenschaften. 93 (9): 413–25. PMID 16688436. doi:10.1007/s00114-006-0120-3
- ↑ Bäckman, Johan; A (1 de abril de 2002). «Harmonic oscillatory orientation relative to the wind in nocturnal roosting flights of the swift Apus apus». The Journal of Experimental Biology. 205 (7): 905–910. PMID 11916987
- ↑ Milius, S. (6 de fevereiro de 1999). «Half-asleep birds choose which half dozes». Science News Online. 155 (6). 86 páginas. JSTOR 4011301. doi:10.2307/4011301
- ↑ Beauchamp, Guy (1999). «The evolution of communal roosting in birds: origin and secondary losses». Behavioural Ecology. 10 (6): 675–87. doi:10.1093/beheco/10.6.675
- ↑ Buttemer, William A. (1985). «Energy relations of winter roost-site utilization by American goldfinches (Carduelis tristis)» (PDF). Oecologia. 68 (1): 126–32. doi:10.1007/BF00379484
- ↑ Buckley, F. G. (1 Janeiro 1968). «Upside-down Resting by Young Green-Rumped Parrotlets (Forpus passerinus)». The Condor. 70 (1). 89 páginas. doi:10.2307/1366517
- ↑ Carpenter, F. Lynn (1974). «Torpor in an Andean Hummingbird: Its Ecological Significance». Science. 183 (4124): 545–47. PMID 17773043. doi:10.1126/science.183.4124.545
- ↑ McKechnie, Andrew E.; Ashdown, Robert A. M.; Christian, Murray B.; Brigham, R. Mark (2007). «Torpor in an African caprimulgid, the freckled nightjar Caprimulgus tristigma». Journal of Avian Biology. 38 (3): 261–66. doi:10.1111/j.2007.0908-8857.04116.x
- ↑ a b Burton, R (1985). Bird Behavior. [S.l.]: Alfred A. Knopf, Inc. ISBN 0-394-53957-5
- ↑ Short, Lester L. (1993). Birds of the World and their Behavior. Nova Iorque: Henry Holt and Co. ISBN 0-8050-1952-9
- ↑ Doutrelant, C; Grégoire, A; Midamegbe, A; Lambrechts, M; Perret, P; Female plumage coloration is sensitive to the cost of reproduction. An experiment in blue tits. (Janeiro de 2012). Journal of Animal Ecology. 81 (1): 87–96. PMID 21819397. doi:10.1111/j.1365-2656.2011.01889.x
- ↑ Edwards, DB (2012). «Immune investment is explained by sexual selection and pace-of-life, but not longevity in parrots (Psittaciformes).». PLOS ONE. 7 (12): e53066. PMID 23300862. doi:10.1371/journal.pone.0053066
- ↑ Freed, Leonard A. (1987). «The Long-Term Pair Bond of Tropical House Wrens: Advantage or Constraint?». The American Naturalist. 130 (4): 507–25. doi:10.1086/284728
- ↑ Gowaty, Patricia A. (1983). «Male Parental Care and Apparent Monogamy among Eastern Bluebirds (Sialia sialis)». The American Naturalist. 121 (2): 149–60. doi:10.1086/284047
- ↑ Westneat, David F.; Stewart, Ian R.K. (2003). «Extra-pair paternity in birds: Causes, correlates, and conflict». Annual Review of Ecology, Evolution, and Systematics. 34: 365–96. doi:10.1146/annurev.ecolsys.34.011802.132439
- ↑ Gowaty, Patricia A.; Buschhaus, Nancy (1998). «Ultimate causation of aggressive and forced copulation in birds: Female resistance, the CODE hypothesis, and social monogamy». American Zoologist. 38 (1): 207–25. doi:10.1093/icb/38.1.207
- ↑ Birkhead, T.R.; Møller, P. (1993). «Sexual selection and the temporal separation of reproductive events: sperm storage data from reptiles, birds and mammals». Biological Journal of the Linnean Society. 50: 295–311. doi:10.1111/j.1095-8312.1993.tb00933.x
- ↑ Sheldon, B (1994). «Male Phenotype, Fertility, and the Pursuit of Extra-Pair Copulations by Female Birds». Proceedings of the Royal Society B. 257 (1348): 25–30. doi:10.1098/rspb.1994.0089
- ↑ Wei, G; Zuo-Hua (2005). «Copulations and mate guarding of the Chinese Egret». Waterbirds. 28 (4): 527–30. doi:10.1675/1524-4695(2005)28[527:CAMGOT]2.0.CO;2 Texto ", Yin; Fu-Min, Lei" ignorado (ajuda);
- ↑ Bagemihl, Bruce (1999). Biological exuberance: Animal homosexuality and natural diversity. Nova Iorque: St. Martin's. pp. 479–655
- ↑ Kokko, H; Harris, M; Wanless, S (2004). «Competition for breeding sites and site-dependent population regulation in a highly colonial seabird, the common guillemot Uria aalge». Journal of Animal Ecology. 73 (2): 367–76. doi:10.1111/j.0021-8790.2004.00813.x
- ↑ a b Hansell, M (2000). Bird Nests and Construction Behaviour. [S.l.]: University of Cambridge Press. ISBN ISBN 0-521-46038-7 Verifique
|isbn=(ajuda) - ↑ Lafuma, L; Lambrechts, M; Raymond, M (2001). «Aromatic plants in bird nests as a protection against Hematophagy». Behavioural Processes. 56 (2): 113–20. doi:10.1016/S0376-6357(01)00191-7 Texto "blood-sucking flying insects? " ignorado (ajuda);
- ↑ Williams, Tony D. (1995). The Penguins. Oxford: Oxford University Press. p. 157. ISBN 0-19-854667-X
- ↑ Booker, L; Booker, M (1991). «Why Are Cuckoos Host Specific?». Oikos. 57 (3): 301–09. JSTOR 3565958. doi:10.2307/3565958
- ↑ Turner, J. Scott (1997). «On the thermal capacity of a bird's egg warmed by a brood patch». Physiological Zoology. 70 (4): 470–80. PMID 9237308. doi:10.1086/515854
- ↑ Warham, J. (1990). The Petrels: Their Ecology and Breeding Systems. [S.l.]: Academic Press. ISBN 0-12-735420-4 Parâmetro desconhecido
|locla=ignorado (ajuda) - ↑ Jones, DN; Dekker, René WRJ, Roselaar, Cees S (1995). The Megapodes. Col: Bird Families of the World. 3. Oxford: Oxford University Press. ISBN ISBN 0-19-854651-3 Verifique
|isbn=(ajuda) - ↑ Ehrlich, Paul R.; Dobkin, David S.; Wheye, Darryl. «Precocial and Altricial Young». Stanford.edu. Consultado em 25 de fevereiro de 2016
- ↑ Elliot, A (1994). «Family Megapodiidae (Megapodes)». In: del Hoyo J, Elliott A, Sargatal J. Handbook of the Birds of the World. 2; New World Vultures to Guineafowl. Barcelona: Lynx Edicions. ISBN 84-87334-15-6
- ↑ Metz, VG; Schreiber, EA (2002). «Great Frigatebird (Fregata minor)». In: Poole, A.; Gill, F. The Birds of North America, No 681. Philadelphia: The Birds of North America Inc
- ↑ Young, Euan (1994). Skua and Penguin. Predator and Prey. [S.l.]: Cambridge University Press. p. 453
- ↑ Ekman, J (2006). «Family living amongst birds». Journal of Avian Biology. 37 (4): 289–98. doi:10.1111/j.2006.0908-8857.03666.x
- ↑ Cockburn A (1996). «Why do so many Australian birds cooperate? Social evolution in the Corvida». In: Floyd R, Sheppard A, de Barro P. Frontiers in Population Ecology. Melbourne: CSIRO. pp. 21–42
- ↑ Cockburn, Andrew (2006). «Prevalence of different modes of parental care in birds». Proceedings of the Royal Society B. 273 (1592): 1375–83. PMC 1560291
- ↑ Gaston, AJ (1994). «Ancient Murrelet (Synthliboramphus antiquus).». In: A. Poole, F. Gill. The Birds of North America. Philadelphia: The Academy of Natural Sciences
- ↑ Schaefer, HC; Eshiamwata, GW; Munyekenye, FB; Böhning-Gaese, K (2004). «Life-history of two African Sylvia warblers: low annual fecundity and long post-fledging care». Ibis. 146 (3): 427–37. doi:10.1111/j.1474-919X.2004.00276.x
- ↑ Alonso, JC; Bautista, LM; Alonso, JA (2004). «Family-based territoriality vs flocking in wintering common cranes Grus grus». Journal of Avian Biology. 35 (5): 434–44. doi:10.1111/j.0908-8857.2004.03290.x
- ↑ a b Davies, N (2000). Cuckoos, Cowbirds and other Cheats. [S.l.]: T. & A. D. Poyser. ISBN 0-85661-135-2 Parâmetro desconhecido
|locla=ignorado (ajuda) - ↑ Sorenson, M (1997). «Effects of intra- and interspecific brood parasitism on a precocial host, the canvasback, Aythya valisineria» (PDF). Behavioral Ecology. 8 (2): 153–61. doi:10.1093/beheco/8.2.153
- ↑ Spottiswoode, C. N.; Colebrook-Robjent, J. F.R. (2007). «Egg puncturing by the brood parasitic Greater Honeyguide and potential host counteradaptations». Behavioral Ecology. 18 (4): 792–799. doi:10.1093/beheco/arm025
- ↑ Burton, Philip (1989). Birds of Prey. [S.l.]: Gallery Books. p. 8. ISBN 0-8317-6381-7
- ↑ a b Clout, M; Hay, J (1989). «The importance of birds as browsers, pollinators and seed dispersers in New Zealand forests» (PDF). New Zealand Journal of Ecology. 12: 27–33
- ↑ Gary Stiles, F. (1981). «Geographical Aspects of Bird-Flower Coevolution, with Particular Reference to Central America». Annals of the Missouri Botanical Garden. 68 (2): 323–51. JSTOR 2398801. doi:10.2307/2398801
- ↑ Temeles, E; Linhart, Y; Masonjones, M; Masonjones, H (2002). «The Role of Flower Width in Hummingbird Bill Length–Flower Length Relationships» (PDF). Biotropica. 34 (1): 68–80. doi:10.1111/j.1744-7429.2002.tb00243.x
- ↑ Bond, William J.; Lee, William G.; Craine, Joseph M. (2004). «Plant structural defences against browsing birds: a legacy of New Zealand's extinct moas». Oikos. 104 (3): 500–08. doi:10.1111/j.0030-1299.2004.12720.x
- ↑ Wainright, S; Haney, J; Kerr, C; Golovkin, A; Flint, M (1998). «Utilization of nitrogen derived from seabird guano by terrestrial and marine plants at St. Paul, Pribilof Islands, Bering Sea, Alaska» (PDF). Marine Ecology. 131 (1): 63–71. doi:10.1007/s002270050297
- ↑ Bosman, A; Hockey, A (1986). «Seabird guano as a determinant of rocky intertidal community structure» (PDF). Marine Ecology Progress Series. 32: 247–57. doi:10.3354/meps032247
- ↑ Bonney, Rick; Rohrbaugh, Ronald (2004). Handbook of Bird Biology 2ª ed. Princeton, NJ: Princeton University Press. ISBN 0-938027-62-X
- ↑ Dean, W; Siegfried R, MacDonald I (1990). «The Fallacy, Fact, and Fate of Guiding Behavior in the Greater Honeyguide». Conservation Biology. 4 (1): 99–101
- ↑ Singer, R.; Yom-Tov, Y. (1988). «The Breeding Biology of the House Sparrow Passer domesticus in Israel». Ornis Scandinavica. 19 (2): 139–44. JSTOR 3676463. doi:10.2307/3676463
- ↑ Reed, KD; Meece, JK; Henkel, JS; Shukla, SK (2003). «Birds, Migration and Emerging Zoonoses: West Nile Virus, Lyme Disease, Influenza A and Enteropathogens». Clinical medicine & research. 1 (1): 5–12. PMC 1069015
- ↑ Brown, Lester (2005). «3: Moving Up the Food Chain Efficiently.». Outgrowing the Earth: The Food Security Challenge in an Age of Falling Water Tables and Rising Temperatures. [S.l.]: earthscan. ISBN 978-1-84407-185-2
- ↑ Simeone, A; Navarro, X (2002). «Human exploitation of seabirds in coastal southern Chile during the mid-Holocene». Rev. Chil. Hist. Nat. 75 (2): 423–31. doi:10.4067/S0716-078X2002000200012
- ↑ Keane, Aidan; Brooke, M.de L.; McGowan, P.J.K. (2005). «Correlates of extinction risk and hunting pressure in gamebirds (Galliformes)». Biological Conservation. 126 (2): 216–33. doi:10.1016/j.biocon.2005.05.011
- ↑ «The Guano War of 1865–1866». World History at KMLA. Consultado em 18 de dezembro de 2007
- ↑ Dolbeer, R (1990). «Ornithology and integrated pest management: Red-winged blackbirds Agleaius phoeniceus and corn». Ibis. 132 (2): 309–22. doi:10.1111/j.1474-919X.1990.tb01048.x
- ↑ Dolbeer, R; Belant, J; Sillings, J (1993). «Shooting Gulls Reduces Strikes with Aircraft at John F. Kennedy International Airport». Wildlife Society Bulletin. 21: 442–50
- ↑ Cooney, R; Jepson, P (2006). «The international wild bird trade: what's wrong with blanket bans?». Oryx. 40 (1): 18–23. doi:10.1017/S0030605306000056
- ↑ Manzi, M; Coomes, O. T. (2002). «Cormorant fishing in Southwestern China: a Traditional Fishery under Siege. (Geographical Field Note)». Geographic Review. 92 (4): 597–603. JSTOR 4140937. doi:10.2307/4140937
- ↑ Pullis La Rouche, G. (2006). «Birding in the United States: a demographic and economic analysis.». In: G.C. Boere, C.A. Galbraith, D.A. Stroud. JNCC.gov.uk Waterbirds around the world Verifique valor
|url=(ajuda) (PDF). Edimburgo: The Stationery Office. pp. 841–46. - ↑ Chamberlain, DE; Vickery, JA; Glue, DE; Robinson, RA; Conway, GJ; Woodburn, RJW; Cannon, AR (2005). «Annual and seasonal trends in the use of garden feeders by birds in winter». Ibis. 147 (3): 563–75. doi:10.1111/j.1474-919x.2005.00430.x
- ↑ Meighan, C. W. (1966). «Prehistoric Rock Paintings in Baja California». American Antiquity. 31 (3): 372–92. JSTOR 2694739. doi:10.2307/2694739
- ↑ Clarke, CP (1908). «A Pedestal of the Platform of the Peacock Throne». The Metropolitan Museum of Art Bulletin. 3 (10): 182–83. JSTOR 3252550. doi:10.2307/3252550
- ↑ Lasky, E. D. (1992). «A Modern Day Albatross: The Valdez and Some of Life's Other Spills». The English Journal. 81 (3): 44–46. JSTOR 820195. doi:10.2307/820195
- ↑ Carson, A (1998). «Vulture Investors, Predators of the 90s: An Ethical Examination» (PDF). Journal of Business Ethics. 17 (5): 543–55. doi:10.1023/A:1017974505642
- ↑ Chandler, A (1934). «The Nightingale in Greek and Latin Poetry». The Classical Journal. 30 (2): 78–84. JSTOR 3289944
- ↑ Ingersoll, Ernest (1923). Birds in legend, fable and folklore. Longmans: Green and co. p. 214
- ↑ Hauser, A. J. (1985). «Jonah: In Pursuit of the Dove». Journal of Biblical Literature. 104 (1): 21–37. JSTOR 3260591. doi:10.2307/3260591
- ↑ Thankappan Nair, P. (1974). «The Peacock Cult in Asia». Asian Folklore Studies. 33 (2): 93–170. JSTOR 1177550. doi:10.2307/1177550
- ↑ Tennyson, A; Martinson P (2006). Extinct Birds of New Zealand. Wellington: Te Papa Press. ISBN 978-0-909010-21-8
- ↑ Routledge, S; Routledge, K (1917). «The Bird Cult of Easter Island». Folklore. 28 (4): 337–55. doi:10.1080/0015587X.1917.9719006
- ↑ Smith, S. (2011). «Generative landscapes: the step mountain motif in Tiwanaku iconography.» (Automatic PDF download). Ancient America. 12: 1–69
- ↑ Dransart, P. (2002). Earth, Water, Fleece and Fabric: An Ethnography and Archaeology of Andean Camelid Herding. [S.l.]: Routledge. pp. 64, 79
- ↑ Enriquez, PL; Mikkola H (1997). «Comparative study of general public owl knowledge in Costa Rica, Central America and Malawi, Africa». In: J.R. Duncan, D.H. Johnson, T.H. Nicholls. Biology and conservation of owls of the Northern Hemisphere. General Technical Report NC-190. Minnesota: USDA Forest Service. pp. 160–66
- ↑ Lewis DP (2005). «Owls in Mythology and Culture». Owlpages.com. Consultado em 15 de setembro de 2007
- ↑ Dupree, N (1974). «An Interpretation of the Role of the Hoopoe in Afghan Folklore and Magic». Folklore. 85 (3): 173–93. JSTOR 1260073. doi:10.1080/0015587X.1974.9716553
- ↑ Fuller, E (2000). Extinct Birds. [S.l.]: Oxford University Press. ISBN 0-19-850837-9 Parâmetro desconhecido
|locla=ignorado (ajuda) - ↑ Steadman, D (2006). Extinction and Biogeography in Tropical Pacific Birds. [S.l.]: University of Chicago Press. ISBN 978-0-226-77142-7
- ↑ «Spotlight on threatened birds». BirdLife International. 2015. Consultado em 6 de março de 2015
- ↑ Norris, K; Pain D (2002). Conserving Bird Biodiversity: General Principles and their Application. [S.l.]: Cambridge University Press. ISBN 978-0-521-78949-3
- ↑ Brothers, NP (1991). «Albatross mortality and associated bait loss in the Japanese longline fishery in the southern ocean». Biological Conservation. 55 (3): 255–68. doi:10.1016/0006-3207(91)90031-4
- ↑ Wurster, D; Wurster, C; Strickland, W (1965). «Bird Mortality Following DDT Spray for Dutch Elm Disease». Ecology. 46 (4): 488–99. doi:10.2307/1934880
- ↑ Wurster, C. F.; Wurster, D. H.; Strickland, W. N. (1965). «Bird Mortality after Spraying for Dutch Elm Disease with DDT». Science. 148 (3666): 90–91. PMID 14258730. doi:10.1126/science.148.3666.90
- ↑ Blackburn, T; Cassey, P; Duncan, R; Evans, K; Gaston, K (2004). «Avian Extinction and Mammalian Introductions on Oceanic Islands». Science. 305 (5692): 1955–58. PMID 15448269. doi:10.1126/science.1101617
- ↑ Butchart, S; Stattersfield, A; Collar, N (2006). «How many bird extinctions have we prevented?» (PDF). Oryx. 40 (3): 266–79. doi:10.1017/S0030605306000950
- ↑ del Hoyo, Josep; Andy Elliott; Jordi Sargatal (1992). Handbook of Birds of the World. 1: Ostrich to Ducks. Barcelona: Lynx Edicions. ISBN 84-87334-10-5
- ↑ Linnaeus, Carolus (1758). Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tomus I. Editio decima, reformata. [S.l.]: Holmiae. (Laurentii Salvii). 824 páginas
- ↑ a b c Livezey, Bradley C.; Zusi, RL (Janeiro 2007). «Higher-order phylogeny of modern birds (Theropoda, Aves: Neornithes) based on comparative anatomy. II. Analysis and discussion». en:Zoological Journal of the Linnean SocietyZoological Journal of the Linnean Society. 149 (1): 1–95. PMC 2517308
- ↑ Padian, Kevin; L.M. Chiappe Chiappe LM (1997). «Bird Origins». In: en:Philip J. Currie, Kevin Padian. Encyclopedia of Dinosaurs. San Diego: Academic Press. pp. 41–96. ISBN 0-12-226810-5
- ↑ a b Gauthier, Jacques (1986). «Saurischian Monophyly and the origin of birds». In: Kevin Padian. The Origin of Birds and the Evolution of Flight. Col: Memoirs of the California Academy of Science 8. [S.l.: s.n.] pp. 1–55. ISBN 0-940228-14-9
- ↑ a b c Pascal Godefroit, Andrea Cau, Hu Dong-Yu, François Escuillié, Wu Wenhao and Gareth Dyke (2013). «A Jurassic avialan dinosaur from China resolves the early phylogenetic history of birds». Nature. in press (7454): 359–62. PMID 23719374. doi:10.1038/nature12168
- ↑ Prum, Richard O. Prum (2008). «Who's Your Daddy». Science. 322 (5909): 1799–1800. PMID 19095929. doi:10.1126/science.1168808
- ↑ Paul, Gregory S. (2002). «Looking for the True Bird Ancestor». Dinosaurs of the Air: The Evolution and Loss of Flight in Dinosaurs and Birds. Baltimore: Johns Hopkins University Press. pp. 171–224. ISBN 0-8018-6763-0
- ↑ Norell, Mark; Mick Ellison (2005). Unearthing the Dragon: The Great Feathered Dinosaur Discovery. New York: Pi Press. ISBN 0-13-186266-9
- ↑ Borenstein, Seth (31 de julho de 2014). «Study traces dinosaur evolution into early birds». Associated Press. Consultado em 3 de agosto de 2014
- ↑ Lee, Michael S. Y.; Cau, Andrea; Naish, Darren; Dyke, Gareth J. (1 de agosto de 2014). «Sustained miniaturization and anatomical innovation in the dinosaurian ancestors of birds». Science. 345 (6196): 562–566. doi:10.1126/science.1252243. Consultado em 2 de agosto de 2014
- ↑ Xing Xu, Hailu You, Kai Du and Fenglu Han (28 Julho 2011). «An Archaeopteryx-like theropod from China and the origin of Avialae». Nature. 475 (7357): 465–470. PMID 21796204. doi:10.1038/nature10288
- ↑ Turner, Alan H.; Pol, D; Clarke, JA; Erickson, GM; Norell, MA (2007). «A basal dromaeosaurid and size evolution preceding avian flight» (PDF). Science. 317 (5843): 1378–1381. PMID 17823350. doi:10.1126/science.1144066
- ↑ Xu, X; Zhou, Z; Wang, X; Kuang, X; Zhang, F; Du, X (2003). «Four-winged dinosaurs from China». Nature. 421 (6921): 335–340. PMID 12540892. doi:10.1038/nature01342
- ↑ Luiggi, Christina (Julho de 2011). «On the Origin of Birds». The Scientist. Consultado em 11 de junho de 2012
- ↑ Mayr, G.; Pohl, B.; Hartman, S.; Peters, D.S. (2007). «The tenth skeletal specimen of Archaeopteryx». Zoological Journal of the Linnean Society. 149: 97–116. doi:10.1111/j.1096-3642.2006.00245.x
- ↑ Lefèvre, U.; Hu, D.; Escuillié, F. O.; Dyke, G.; Godefroit, P. (2014). «A new long-tailed basal bird from the Lower Cretaceous of north-eastern China». Biological Journal of the Linnean Society. 113 (3): 790–804. doi:10.1111/bij.12343
- ↑ a b Zheng, X.; Zhou, Z.; Wang, X.; Zhang, F.; Zhang, X.; Wang, Y.; Wei, G.; Wang, S.; Xu, X. (2013). «Hind Wings in Basal Birds and the Evolution of Leg Feathers». Science. 339 (6125): 1309–1312. PMID 23493711. doi:10.1126/science.1228753
- ↑ a b c d Chiappe, Luis M. (2007). Glorified Dinosaurs: The Origin and Early Evolution of Birds. Sydney: University of New South Wales Press. ISBN 978-0-86840-413-4
- ↑ Agency France-Presse (Abril 2011). «Birds survived dino extinction with keen senses». Cosmos Magazine. Consultado em 11 Junho de 2012
- ↑ Wang, M.; Zheng, X.; o’Connor, J. K.; Lloyd, G. T.; Wang, X.; Wang, Y.; Zhang, X.; Zhou, Z. (2015). «The oldest record of ornithuromorpha from the early cretaceous of China». Nature Communications. 6. 6987 páginas. PMID 25942493. doi:10.1038/ncomms7987
- ↑ Louchart, A.; Viriot, L. (2011). «From snout to beak: the loss of teeth in birds». Trends In Ecology & Evolution. 26 (12): 663–673. doi:10.1016/j.tree.2011.09.004
- ↑ Clarke, J. A.; Zhou, Z.; Zhang, F. (2006). «Insight into the evolution of avian flight from a new clade of Early Cretaceous ornithurines from China and the morphology of Yixianornis grabaui». Journal of Anatomy. 208 (3): 287–308. PMC 2100246
- ↑ Ritchison, Gary. «Bird biogeography». Avian Biology. Eastern Kentucky University. Consultado em 10 de Abril de 2008
- ↑ Clements, James F. (2007). he Clements Checklist of Birds of the World 6ª ed. Ithaca: Cornell University Press. ISBN 978-0-8014-4501-9
- ↑ Gill, Frank (2006). Birds of the World: Recommended English Names. Princeton: Princeton University Press. ISBN 978-0-691-12827-6
- ↑ Clarke, Julia A.; Tambussi, CP; Noriega, JI; Erickson, GM; Ketcham, RA (2005). «Definitive fossil evidence for the extant avian radiation in the Cretaceous» (PDF). Nature. 433 (7023): 305–308. PMID 15662422. doi:10.1038/nature03150 Nature.com, Informação adicional]
- ↑ a b Ericson, Per G.P.; Anderson, CL; Britton, T; Elzanowski, A; Johansson, US; Källersjö, M; Ohlson, J; Parsons, TJ; Zuccon, D (2006). «Diversification of Neoaves: integration of molecular sequence data and fossils» (PDF). Biology Letters. 2 (4): 543–547. PMC 1834003
|formaot=ignorado (ajuda); - ↑ Brown, Joseph W.; Payne, RB; Mindell, DP (Junho de 2007). «Nuclear DNA does not reconcile 'rocks' and 'clocks' in Neoaves: a comment on Ericson et al.». Biology Letters. 3 (3): 257–259. PMC 2464679
- ↑ «Ornithologists Publish Most Comprehensive Avian Tree of Life». Sci-news.com. 9 de outubro de 2015. Consultado em 14 de fevereiro de 2016
- ↑ a b Jarvis, E.D.; et al (2014). «Whole-genome analyses resolve early branches in the tree of life of modern birds». Science. 346 (6215): 1320–1331. PMID 25504713. doi:10.1126/science.1253451
- ↑ Yuri, T.; et al (2013). «Parsimony and Model-Based Analyses of Indels in Avian Nuclear Genes Reveal Congruent and Incongruent Phylogenetic Signals». Biology. 2 (1): 419–444. doi:10.3390/biology2010419
- ↑ Sibley, Charles; Jon Edward Ahlquist (1990). Phylogeny and classification of birds. New Haven: Yale University Press. ISBN 0-300-04085-7
- ↑ Mayr, Ernst; Short, Lester L. (1970). Species Taxa of North American Birds: A Contribution to Comparative Systematics. Col: Publications of the Nuttall Ornithological Club, no. 9. Cambridge, Mass.: Nuttall Ornithological Club. OCLC 517185
Ligações externas
- «SPEA: Sociedade Portuguesa para o Estudo das Aves»
- «Aves de Portugal»
- «Aves do Brasil»
- «Avibase - the world bird database» (em inglês)
- «Aves, na Encyclopedia of Life» (em inglês)
- «SORA: Searchable Ornithological Research Archive» (em inglês)
- «Essays on bird biology, Universidade de Stanford» (em inglês)
- «BirdLife International» (em inglês)
- «Ornithology.com» (em inglês)
- «Dinossauros com penas em www.dinosaur-world.com» (em inglês)