Glukosa 6-fosfatase
| Glukosa 6-fosfatase | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Pengenal pasti | |||||||||
| Nombor EC | 3.1.3.9 | ||||||||
| Nombor CAS | 9001-39-2 | ||||||||
| Pangkalan data | |||||||||
| IntEnz | Lihat IntEnz | ||||||||
| BRENDA | Entri BRENDA | ||||||||
| ExPASy | Lihat NiceZyme | ||||||||
| KEGG | Entri KEGG | ||||||||
| MetaCyc | Laluan metabolik | ||||||||
| PRIAM | Profil | ||||||||
| Struktur PDB | RCSB PDB PDBj PDBe PDBsum | ||||||||
| Ontologi gen | AmiGO / EGO | ||||||||
| |||||||||

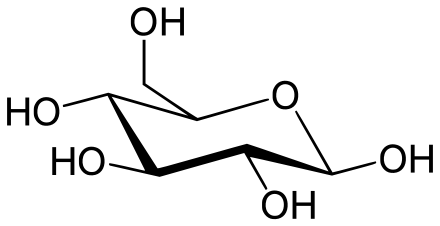
Enzim glukosa 6-fosfatase (EC 3.1.3.9, G6Pase; nama sistematik D-glukosa-6-fosfat fosfohidrolase) memangkinkan hidrolisis glukosa 6-fosfat, menghasilkan penciptaan kumpulan fosfat dan glukosa bebas:
- D-glukosa 6-fosfat + H2O =D-glukosa + fosfat
Glukosa kemudiannya dieksport dari sel melalui protein membran pengangkut glukosa.[1] Pemangkinan ini melengkapkan langkah terakhir dalam glukoneogenesis, dan oleh itu, memainkan peranan penting dalam pengawalan homeostatik paras glukosa darah.[2]
Glukosa 6-fosfatase ialah kompleks protein berbilang komponen, termasuk pengangkut untuk G6P, glukosa dan fosfat. Fungsi fosfatase utama dilakukan oleh subunit pemangkin glukosa 6-fosfatase. Pada manusia, terdapat tiga isozim subunit pemangkin: glukosa 6-fosfatase-α, dikodkan oleh G6PC ; IGRP, dikodkan oleh G6PC2; dan glukosa 6-fosfatase-β, dikodkan oleh G6PC3.[3]
Glukosa 6-fosfatase-α dan glukosa 6-fosfatase-β kedua-dua ialah fosfohidrolase berfungsi, dan mempunyai struktur tapak aktif, topologi, mekanisme tindakan, dan sifat kinetik yang serupa berkenaan dengan hidrolisis G6P.[4] Sebaliknya, IGRP hampir tidak mempunyai aktiviti hidrolase, dan mungkin memainkan peranan yang berbeza dalam merangsang rembesan insulin pankreas.[5]
Struktur dan fungsi
[sunting | sunting sumber]Walaupun persetujuan yang jelas belum dicapai, sebilangan besar saintis mematuhi model pengangkutan substrat untuk mengambil kira sifat pemangkin glukosa 6-fosfatase. Dalam model ini, glukosa 6-fosfatase mempunyai tahap kekhususan yang rendah. Pemindahan glukosa 6-fosfat dijalankan oleh protein pengangkut (T1) dan retikulum endoplasma (ER) mengandungi struktur yang membenarkan keluarnya kumpulan fosfat (T2) dan glukosa (T3).[6]
Glukosa 6-fosfatase terdiri daripada 357 asid amino, dan dipasang pada retikulum endoplasma (ER) oleh sembilan heliks transmembran. Terminal N dan tapak aktifnya terdapat di bahagian lumen ER, dan terminal C-nya mengunjur ke dalam sitoplasma. Oleh kerana hubungannya yang ketat dengan ER, struktur sebenar glukosa 6-fosfatase masih tidak diketahui. Walau bagaimanapun, penjajaran jujukan telah menunjukkan bahawa glukosa 6-fosfatase secara struktur adalah serupa dengan tapak aktif kloroperoksidase berkandungan vanadium yang terdapat dalam Curvularia inaequalis.[7]
Berdasarkan kajian kinetik pH pemangkinan glukosa 6-fosfatase-α, ada cadangan bahawa hidrolisis glukosa 6-fosfat telah diselesaikan melalui perantaraan glukosa 6-fosfat fosfohistidina kovalen. Tapak aktif glukosa 6-fosfatase-α pada mulanya dikenal pasti dengan kehadiran motif tandatangan fosfat terpelihara yang biasanya ditemui dalam fosfatase lipid, fosfatase asid, dan haloperoksidase vanadium.[4]
Sisa-sisa penting dalam tapak aktif haloperoksidases vanadium termasuk Lys353, Arg360, Arg490, His404 dan His496. Sisa yang sepadan dalam tapak aktif glukosa 6-fosfastase-α termasuk Arg170 dan Arg83 yang menderma ion hidrogen kepada fosfat, menstabilkan keadaan peralihan, His119 yang menyediakan proton kepada oksigen terdefosforilasi yang melekat pada glukosa, dan His176 yang melengkapkan serangan nukleofilik ke atas fosfat untuk membentuk perantara enzim fosforil terikat secara kovalen.[1] Dalam kloroperoksidase berkandungan vanadium, Lys353 didapati menstabilkan fosfat dalam keadaan peralihan. Walau bagaimanapun, sisa yang sepadan dalam glukosa 6-fosfatase-α (Lys76) berada dalam membran ER dan fungsinya, jika ada, pada masa ini tidak dapat ditentukan. Dengan pengecualian Lys76, sisa-sisa ini semuanya terletak pada bahagian lumen ER.[4]
Glukosa 6-fosfatase-β ialah protein membran asid 346 amino yang dinyatakan di merata tempat, dan berkongsi 36% identiti jujukan dengan glukosa 6-fosfatase-α. Dalam enzim glukosa 6-fosfatase-β, penjajaran jujukan meramalkan bahawa tapak aktifnya mengandungi His167, His114 dan Arg79. Sama seperti tapak aktif glukosa 6-fosfatase-α, His167 ialah sisa yang menyediakan serangan nukleofilik, dan His114, dan Arg79 ialah penderma hidrogen. Glukosa 6-fosfatase-β juga disetempat dalam membran ER, walaupun orientasinya tidak diketahui.[4]
Mekanisme
[sunting | sunting sumber]Hidrolisis glukosa 6-fosfat bermula dengan serangan nukleofilik di fosfat terikat gula oleh His176, mengakibatkan pembentukan ikatan fosfohistidina dan penguaraian karbonil. Oksigen yang bercas negatif kemudian memindahkan elektronnya membentuk semula karbonil dan memutuskan ikatannya dengan glukosa. Oksigen terikat glukosa bercas negatif kemudiannya diprotonkan oleh His119, membentuk glukosa bebas. Perantara fosfat yang dihasilkan oleh tindak balas antara His176 dan kumpulan fosfat kemudiannya dipecahkan oleh serangan hidrofilik; selepas penambahan hidroksida lain dan penguraian karbonil, karbonil direformasi menendang elektron yang asalnya didermakan oleh sisa His176, dengan itu, mewujudkan kumpulan fosfat bebas, dan melengkapkan hidrolisis.[1]
Ekspresi
[sunting | sunting sumber]Pengekodan gen enzim terutamanya berlaku dalam hati, korteks buah pinggang, dan (sedikit sebanyak) di sel β pulau pankreas dan mukosa usus (terutamanya semasa kelaparan).[6] Menurut Surholt dan Newsholme, glukosa 6-fosfatase terdapat dalam pelbagai jenis otot di seluruh alam haiwan, walaupun dalam kepekatan yang sangat rendah.[8] Oleh itu, glikogen yang disimpan oleh otot biasanya tidak tersedia buat seluruh sel badan kerana glukosa 6-fosfat tidak boleh menyeberangi sarkolema melainkan ia dinyahfosforilkan. Enzim memainkan peranan penting semasa tempoh berpuasa dan apabila tahap glukosa rendah. Ada dapatan bahawa kebuluran dan diabetes mendorong peningkatan dua hingga tiga kali ganda dalam aktiviti glukosa 6-fosfatase dalam hati.[6] Aktiviti Glc 6-Pase juga meningkat secara mendadak semasa lahir apabila organisma menjadi bebas daripada sumber glukosa ibu. Gen Glc 6-Pase manusia mengandungi lima ekson yang merangkumi kira-kira 125.5 kb DNA yang terletak pada kromosom 17q21.[9]
Kepentingan klinikal
[sunting | sunting sumber]Mutasi sistem glukosa 6-fosfatase, khususnya mutasi di subunit glukosa 6-fosfatase-α (glukosa 6-fosfatase-α), pengangkut glukosa 6-fosfat (G6PT) dan subunit glukosa 6-fosfatase-β (G6PC3), membawa kepada kekurangan dalam penyelenggaraan homeostasis glukosa interprandial dan fungsi serta homeostasis neutrofil.[10][11] Mutasi dalam kedua-dua glukosa 6-fosfatase-α dan G6PT membawa kepada penyakit simpanan glikogen jenis I (GSD 1, penyakit von Gierke).[12] Secara khusus, mutasi dalam glukosa-6-fosfatase-α membawa kepada penyakit simpanan glikogen jenis 1a yang dicirikan oleh pengumpulan glikogen dan lemak dalam hati dan buah pinggang, mengakibatkan hepatomegali dan renomegali.[13] GSD-1a membentuk kira-kira 80% daripada kes GSD-1 yang hadir secara klinikal.[14] Ketiadaan G6PT membawa kepada GSD-1b (GSD-1b), yang dicirikan oleh kekurangan G6PT dan mewakili 20% daripada kes yang hadir secara klinikal.[14][15]

Punca khusus GSD-1a berpunca daripada mutasi bukan deria, sisipan/pemadaman dengan atau tanpa anjakan bingkai bacaan atau mutasi tapak sambatan yang berlaku di peringkat genetik.[6] Mutasi sala deria menjejaskan dua gelung lumen besar dan heliks transmembran glukosa 6-fosfatase-α, lalu melenyapkan atau mengurangkan aktiviti enzim secara ketara.[6] Punca khusus GSD-1b berpunca daripada mutasi "teruk" seperti mutasi sambatan, mutasi anjakan bingkai dan penggantian sisa yang sangat terpelihara yang memusnahkan aktiviti G6PT sepenuhnya.[6] Mutasi ini membawa kepada kelaziman GSD-1 dengan menghalang pengangkutan glukosa-6-fosfat (G6P) ke dalam bahagian lumen ER, dan juga menghalang penukaran G6P kepada glukosa untuk digunakan oleh sel.
Jenis ketiga kekurangan glukosa 6-fosfatase, kekurangan glukosa 6-fosfatase-β, dicirikan oleh sindrom neutropenia kongenital, di mana neutrofil menunjukkan tekanan ER yang dipertingkatkan, peningkatan apoptosis, homeostasis tenaga terjejas, dan kefungsian terjejas.[16] Ia juga boleh menyebabkan kecacatan jantung dan urogenital.[17] Kelas kekurangan ketiga ini juga dipengaruhi oleh kekurangan G6PT kerana glukosa-6-fosfatase-β juga terletak dalam lumen ER, dan dengan itu, boleh membawa kepada gejala kekurangan glukosa-6-fosfatase-β yang serupa dikaitkan dengan GSD-1b.[15] Tambahan pula, kajian baru-baru ini telah menjelaskan kawasan persamaan ini antara kedua-dua kekurangan dan telah menunjukkan bahawa penyimpangan Pengglikosilan berlaku dalam kedua-dua jenis kekurangan.[18] Glikosilasi neutrofil mempunyai kesan yang mendalam terhadap aktiviti neutrofil, dan oleh itu, juga boleh diklasifikasikan sebagai gangguan glikosilasi kongenital.[18]
Fungsi utama glukosa 6-fosfatase-β telah ditentukan untuk membekalkan glukosa kitar semula kepada sitoplasma neutrofil untuk mengekalkan fungsi normal. Gangguan nisbah glukosa kepada G6P akibat penurunan ketara paras glukosa intrasel menyebabkan gangguan glikolisis dan laluan pentosa fosfat yang ketara.[11] Melainkan ia diatasi dengan pengambilan glukosa ekstrasel, kekurangan ini membawa kepada penyahfungsian neutrofil.[11]
Sebatian vanadium seperti vanadil sulfate telah ditunjukkan dapat merencat enzim, dan dengan itu meningkatkan kepekaan insulin in vivo dalam pesakit kencing manis, seperti yang dinilai oleh teknik pengapit hiperinsulin yang mungkin mempunyai potensi implikasi terapeutik.[19][20]
Lihat juga
[sunting | sunting sumber]Rujukan
[sunting | sunting sumber]- ^ a b c "The catalytic center of glucose-6-phosphatase. HIS176 is the nucleophile forming the phosphohistidine-enzyme intermediate during catalysis". The Journal of Biological Chemistry. 277 (36): 32837–42. September 2002. doi:10.1074/jbc.M201853200. PMID 12093795.
- ^ Nordlie R, dll. (1985). The Enzymes of biological membranes, 2nd edition. New York: Plenum Press. m/s. 349–398. ISBN 0-306-41453-8.
- ^ "Glucose-6-phosphatase catalytic subunit gene family". The Journal of Biological Chemistry. 284 (43): 29241–5. October 2009. doi:10.1074/jbc.R109.025544. PMC 2785553. PMID 19700406.
- ^ a b c d "Histidine 167 is the phosphate acceptor in glucose-6-phosphatase-β forming a phosphohistidine enzyme intermediate during catalysis". The Journal of Biological Chemistry. 279 (13): 12479–83. March 2004. doi:10.1074/jbc.M313271200. PMID 14718531.
- ^ "In islet-specific glucose-6-phosphatase-related protein, the β cell antigenic sequence that is targeted in diabetes is not responsible for the loss of phosphohydrolase activity". Diabetologia. 48 (9): 1851–9. September 2005. doi:10.1007/s00125-005-1848-6. PMID 16012821.
- ^ a b c d e f "The glucose-6-phosphatase system". The Biochemical Journal. 362 (Pt 3): 513–32. March 2002. doi:10.1042/0264-6021:3620513. PMC 1222414. PMID 11879177.
- ^ "Transmembrane topology of glucose-6-phosphatase". The Journal of Biological Chemistry. 273 (11): 6144–8. March 1998. doi:10.1074/jbc.273.11.6144. PMID 9497333.
- ^ Surholt, B; Newsholme, EA (15 September 1981). "Maximum activities and properties of glucose 6-phosphatase in muscles from vertebrates and invertebrates". The Biochemical Journal. 198 (3): 621–9. doi:10.1042/bj1980621. PMC 1163310. PMID 6275855.
- ^ "Glycogen storage disease type Ia in Argentina: two novel glucose-6-phosphatase mutations affecting protein stability". Molecular Genetics and Metabolism. 83 (3): 276–9. November 2004. doi:10.1016/j.ymgme.2004.06.010. PMID 15542400.
- ^ "Glycogen storage disease type I and glucose-6-phosphatase-β deficiency: etiology and therapy". Nature Reviews. Endocrinology. 6 (12): 676–88. December 2010. doi:10.1038/nrendo.2010.189. PMC 4178929. PMID 20975743.
- ^ a b c "Lack of glucose recycling between endoplasmic reticulum and cytoplasm underlies cellular dysfunction in glucose-6-phosphatase-β-deficient neutrophils in a congenital neutropenia syndrome". Blood. 116 (15): 2783–92. October 2010. doi:10.1182/blood-2009-12-258491. PMC 2974586. PMID 20498302.
- ^ Stryer, Lubert; Berg, Jeremy Mark; Tymoczko, John L. (2007). Biochemistry. San Francisco: W.H. Freeman. ISBN 978-0-7167-8724-2.
- ^ "Glycogen Storage Disease Type I". 1993. PMID 20301489. Cite journal requires
|journal=(bantuan) - ^ a b "Type I glycogen storage diseases: disorders of the glucose-6-phosphatase complex". Current Molecular Medicine. 2 (2): 121–43. March 2002. doi:10.2174/1566524024605798. PMID 11949931.
- ^ a b "Glucose-6-phosphatase deficiency". Orphanet Journal of Rare Diseases. 6: 27. 2011. doi:10.1186/1750-1172-6-27. PMC 3118311. PMID 21599942.
- ^ "G-CSF improves murine G6PC3-deficient neutrophil function by modulating apoptosis and energy homeostasis". Blood. 117 (14): 3881–92. April 2011. doi:10.1182/blood-2010-08-302059. PMC 3083300. PMID 21292774.
- ^ "A syndrome with congenital neutropenia and mutations in G6PC3". The New England Journal of Medicine. 360 (1): 32–43. Jan 2009. doi:10.1056/NEJMoa0805051. PMC 2778311. PMID 19118303.
- ^ a b "G6PC3 mutations are associated with a major defect of glycosylation: a novel mechanism for neutrophil dysfunction". Glycobiology. 21 (7): 914–24. July 2011. doi:10.1093/glycob/cwr023. PMC 3110488. PMID 21385794.
- ^ "Effects of vanadyl sulfate on carbohydrate and lipid metabolism in patients with non—insulin-dependent diabetes mellitus - Metabolism - Clinical and Experimental". www.metabolismjournal.com. Dicapai pada 2015-06-16.
- ^ Shehzad, Saima (2013-01-01). "The potential effect of vanadium compounds on glucose-6-phosphatase". Bioscience Horizons. 6: hzt002. doi:10.1093/biohorizons/hzt002. ISSN 1754-7431.
Pautan luar
[sunting | sunting sumber]- Glucose-6-Phosphatase dalam Tajuk Subjek Perubatan (MeSH) di Perpustakaan Perubatan Negara AS
- G6PC, G6PC2, G6PC3, G6PR
- EC 3.1.3.9