アクチン



アクチン (Actin) は螺旋状の多量体を形成してマイクロフィラメントの1種であるアクチンフィラメントを形作る球形のタンパク質である。
この繊維は真核生物の細胞内部で3次元の繊維状構造を作る3つの細胞骨格(アクチンフィラメント、微小管、中間径フィラメント)の中では最も細いものである。アクチンフィラメントは細胞の形を決定している。細胞質流動と、細胞分裂での収縮に関与している。筋細胞ではミオシンと共に筋収縮を担う。また、仮足を介して移動を可能にする。細胞質基質内では、アクチンは種の間での差異が少なく非常に保存されたタンパク質の1つで、藻類とヒトの間で5%しか違わない。恒温脊椎動物では、アクチンには6種類のアイソフォームが存在する[1][2]。
アクチンフィラメント
[編集]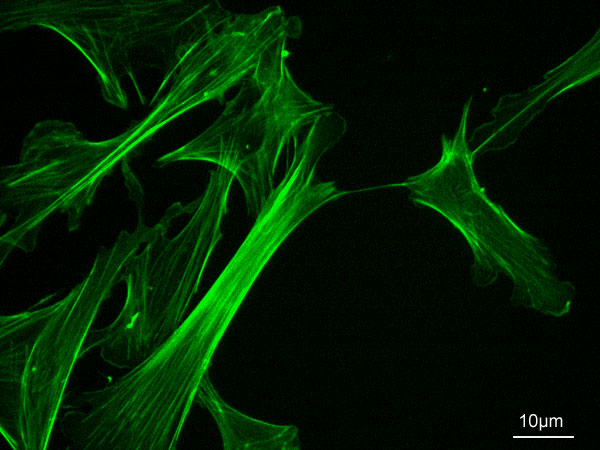
分子量が約42,000の球形 (globular) アクチンはG-アクチンと呼ばれ、G-アクチンが多数重合した糸状 (filamentous) の重合体はF-アクチンと呼ばれ「マイクロフィラメント」になる。マイクロフィラメントは3種の細胞骨格の中でも最も細く、直径6-7nmである。F-アクチンはG-アクチンが右巻き2重螺旋構造でつながり、13個ほどが含まれる36nm程で半周期となる。G-アクチンの重合による成長と脱重合による消失はそれぞれ特定の片側端部で通常行なわれるため、微小管と同様にアクチンフィラメントには方向性があり、成長側を「+端」(プラス端)、消失側を「-端」(マイナス端)と呼ぶ。また、電子顕微鏡写真ではミオシンの結合がやじりのように見えるため、やじりの先端となる-端を「やじり端」「P端」(pointed end) 、その反対側の+端を「反やじり端」「B端」(barbed end) と呼ぶこともある。G-アクチンの細胞内濃度が重合反応と脱重合反応を律速しており、2つの反応が等しくなるG-アクチンの濃度を臨界濃度と呼ぶ。臨界濃度は+端で0.1μM、-端では0.6μMであり、この間の濃度では、+端の方が臨界濃度が低いため重合が進み、-端では脱重合進む。このため定常状態では+端で伸張し-端で短縮している。これはトレッドミリングと呼ばれる[3]。
アクチンは主にATPと結合するがADPとも結合しうる。ATPとアクチンの複合体はADPとアクチンの複合体よりも速く重合し、遅く脱重合する。
このアクチンの重合である核形成の過程は3つのGアクチン単量体が三量体へ合体する事に始まる。この時、ATPの付いたアクチンは+端へ結合し、ATPはその後加水分解され、それによって隣りのユニットとの結合長が短縮して一般に繊維は不安定になる。ADPの付いたアクチンは-端から脱重合し、ADP結合アクチンの増加が結合したADPのATPへの交換を刺激してATP結合アクチンを増やす。この速い代謝回転が細胞の動作に重要である。
コフィリンというタンパク質はADP結合アクチンユニットに結合して、-端からの脱重合を促進し、再構築を妨害する。プロフィリンというタンパク質は、結合するADPとATPの交換を刺激することでこの効果を逆転させる。加えて、プロフィリンと結合したATP結合アクチンユニットはコフィリンと分離して自由に重合できる。他にフィラメントの生成に重要な成分にArp2/3タンパク質があり、これは核形成の場として役に立ち、Gアクチン三量体の形成を刺激する。これら3つのタンパク質は全て細胞シグナルの機構で調節される。
組織化
[編集]繊維束とネットワークは、アクチンフィラメントを構成する2つの一般的な構造のタイプである。アクチン結合タンパク質がアクチンフィラメントと架橋結合をしてどちらの構造を形成するか指示する。
繊維束
[編集]
平行繊維束と収縮性繊維束の2種類のアクチン束がある。平行繊維束では、繊維はアクチン束化タンパク質のフィンブリンから14nm離れて存在している。平行繊維束は細胞の微絨毛の支持をしている。脊椎動物では、アクチン結合タンパク質のビリンが殆どの腸管の細胞にみられる。
筋収縮
[編集]アクチンはミオシン繊維と一緒になって、筋収縮を起こさせるアクトミオシンを形成する。アクチンはATPをエネルギーとして利用する。ATPは加水分解を通してミオシンの頭部を伸ばし、アクチンフィラメントとの結合を可能にする。ADPを使用しての弛緩または収縮運動でアクチンフィラメントが動いてからミオシンの頭部は開放される。
収縮性繊維束では、アクチン繊維束タンパク質のアクチニンはそれぞれ40nmずつ離れている。これだけ離れていることで、モータータンパク質であるミオシンがフィラメントと相互作用できるようにでき、変形と収縮をできる様になっている。この変形ではミオシンの片方の端は原形質膜に結合し、反対側の端はアクチンフィラメントの+端へ向かって移動する。これが細胞の皮質に較べて違う形に膜を押す。収縮に際し、ミオシン分子は通常二つの別のフィラメントに結合し、両端は同時に+端へ向かって、互いのアクチンフィラメントを滑る。これがアクチン繊維束(フィラメントではない)の収縮をもたらす。この機構が筋収縮と細胞分裂の原因となっている。
仮足による運動
[編集]+端から伸長するアクチンフィラメントが細胞膜を力学的に押すことで細胞の端を前に伸ばすことができる。この運動は間期の繊維芽細胞で見られ、長く枝分かれのないアクチンフィラメントが押す糸状仮足 (filopodia) と短く枝分かれしたアクチンフィラメントが押す葉状仮足 (lamellipodia) の形態があるがいずれも、アクチンフィラメントが押し前進した後からストレスファイバー (stress fiber) が形成される。ストレスファイバーはアクチンフィラメントがα-アクチニンによって互いに架橋された構造を持ち、両端はインテグリンによって細胞膜部分につながっている[3]。
収縮環
[編集]細胞分裂の終期に1つの細胞の中央に分裂溝を作って2つの細胞へとくびり切る収縮環 (contractile ring) は、主にアクチンフィラメントとミオシンIIから構成される[3]。
ネットワーク
[編集]多数のアクチン結合タンパク質(フィラミンと呼ばれる)の伴うアクチンネットワークが細胞の皮質に形成される。近年の研究では原形質膜内の物質の拡散の障壁として役に立っている事が示された。
脚注
[編集]出典
[編集]- ^ Gunning PW, Ghoshdastider U, Whitaker S, Popp D, Robinson RC (2015). “The evolution of compositionally and functionally distinct actin filaments”. Journal of Cell Science 128 (11): 2009–19. doi:10.1242/jcs.165563. PMID 25788699.
- ^ Ghoshdastider U, Jiang S, Popp D, Robinson RC (2015). “In search of the primordial actin filament.”. Proc Natl Acad Sci U S A. 112 (30): 9150-1. doi:10.1073/pnas.1511568112. PMID 26178194.
- ^ a b c 田村隆明・山本雅著 『分子生物学イラストレイテッド』 羊土社 2009年3月10日第3版発行 ISBN 978-4-7581-2002-9
外部リンク
[編集]- アクチン - 脳科学辞典
- αアクチニン - 脳科学辞典 アクチン結合タンパク質であるα-アクチニンに関する解説。
- 蛋白質構造データバンク 今月の分子19:アクチン(Actin)