Human skeletal system
The skeletal system includes the bones of the skeleton and the cartilages, ligaments, and other connective tissues that stabilize or interconnect them. Bones are the organs of the skeletal system, and they do more than serve as racks that muscles hang from; they support your weight and work together with muscles, producing controlled, precise movements. Without a framework of bones to connect to, contracting muscles would just get shorter and fatter. Our muscles must pull against the skeleton to make you sit, stand, walk, or run.
The bones of the skeleton are complex, dynamic organs that contain osseous tissue, other connective tissues, smooth muscle tissue, and neural tissue.
The skeleton has many vital functions:
- Support: The skeletal system provides structural support for the entire body. Individual bones or groups of bones provide a framework for the attachment of soft tissues and organs and muscles use to cause movement.
- Mineral storage: The calcium salts of bone are a valuable mineral reserve that maintains normal concentrations of calcium and phosphate ions in body fluids. Calcium is the most abundant mineral in the human body. A typical human body contains 1–2 kg (2.2–4.4 lb) of calcium, with more than 99 percent of it in the bones of the skeleton.
- Blood cell production: Within certain bones, a connective tissue called red bone marrow produces red blood cells, white blood cells, and platelets, a process called hemopoiesis. Red bone marrow consists of developing blood cells, adipocytes, fibroblasts, and macrophages within a network of reticular fibers. It is present in developing bones of the fetus and in some adult bones, such as the hip (pelvic) bones, ribs, sternum (breastbone), vertebrae (backbones), skull, and ends of the bones of the humerus (arm bone) and femur (thigh bone). In a newborn, all bone marrow is red and is involved in hemopoiesis. With increasing age, much of the bone marrow changes from red to yellow.
- Triglyceride storage. Yellow bone marrow consists mainly of adipose cells, which store triglycerides. The stored triglycerides are a potential chemical energy reserve.
- Protection: Skeletal system protects and supports body organs and delicate tissues. The ribs protect the heart and lungs, the skull encloses the brain, the vertebrae shield the spinal cord, and the pelvis cradles delicate digestive and reproductive organs.
- Leverage: Many bones of the skeleton function as levers. They change the magnitude and direction of the forces generated by skeletal muscles. The movements produced range from the delicate motion of a fingertip to powerful changes in the position of the entire body.
Bones in human body
The skeleton is divided into two regions:
- The Axial skeleton and
- Appendicular skeleton.
The axial skeleton, which forms the central supporting axis of the body, includes:
- the skull,
- auditory ossicles,
- hyoid bone,
- vertebral column, and
- thoracic cage (ribs and sternum).
The appendicular skeleton includes:
- the bones of the upper limb and
- pectoral girdle and the bones of the lower limb and pelvic girdle.
Bones of the Skeletal System
It is often stated that there are 206 bones in the skeleton, but this is only a typical adult count, not an invariable number. At birth there are about 270, and even more bones form during childhood. With age, however, the number decreases as separate bones gradually fuse. For example, each side of a child’s pelvic girdle has three bones—the ilium, ischium, and pubis—but in adults, these are fused into a single hip bone on each side. The fusion of several bones, completed by late adolescence to the mid-20s, brings about the average adult number of 206.
This number varies even among adults. One reason is the development of sesamoid bones—bones that form within some tendons in response to strain. The patella (kneecap) is the largest of these; most of the others are small, rounded bones in such locations as the hands and feet. Another reason for adult variation is that some people have extra bones in the skull called sutural bones.
Figure 1. Human skeleton
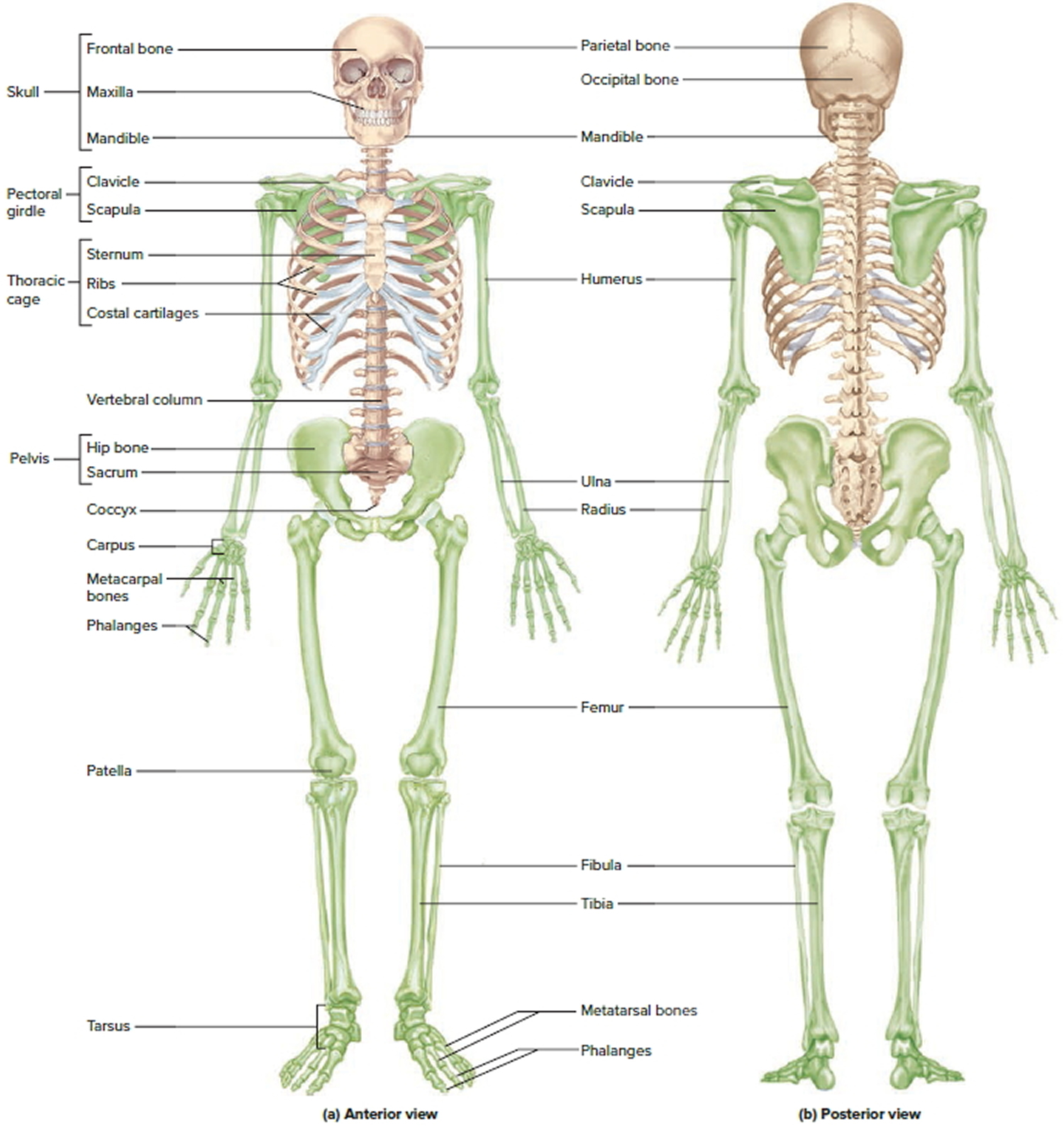
Note: Green colored bones are Appendicular skeleton. The rest are Axial skeleton.
Figure 2. Skull bone


Figure 3. Cervical vertebrae

Figure 4. Vertebral column
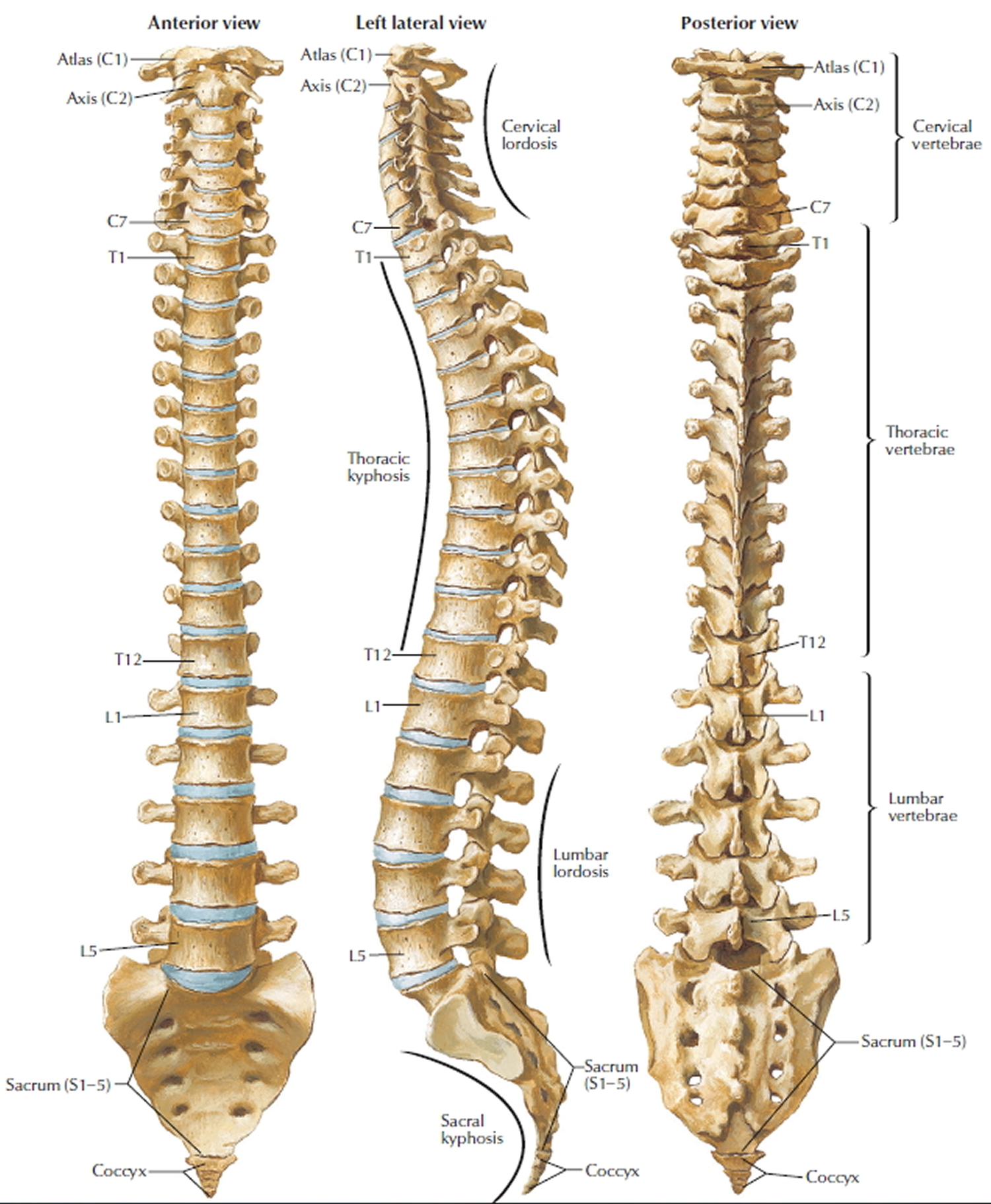
Figure 5. Thoracic bones

Figure 6. Lumbar spine and pelvis
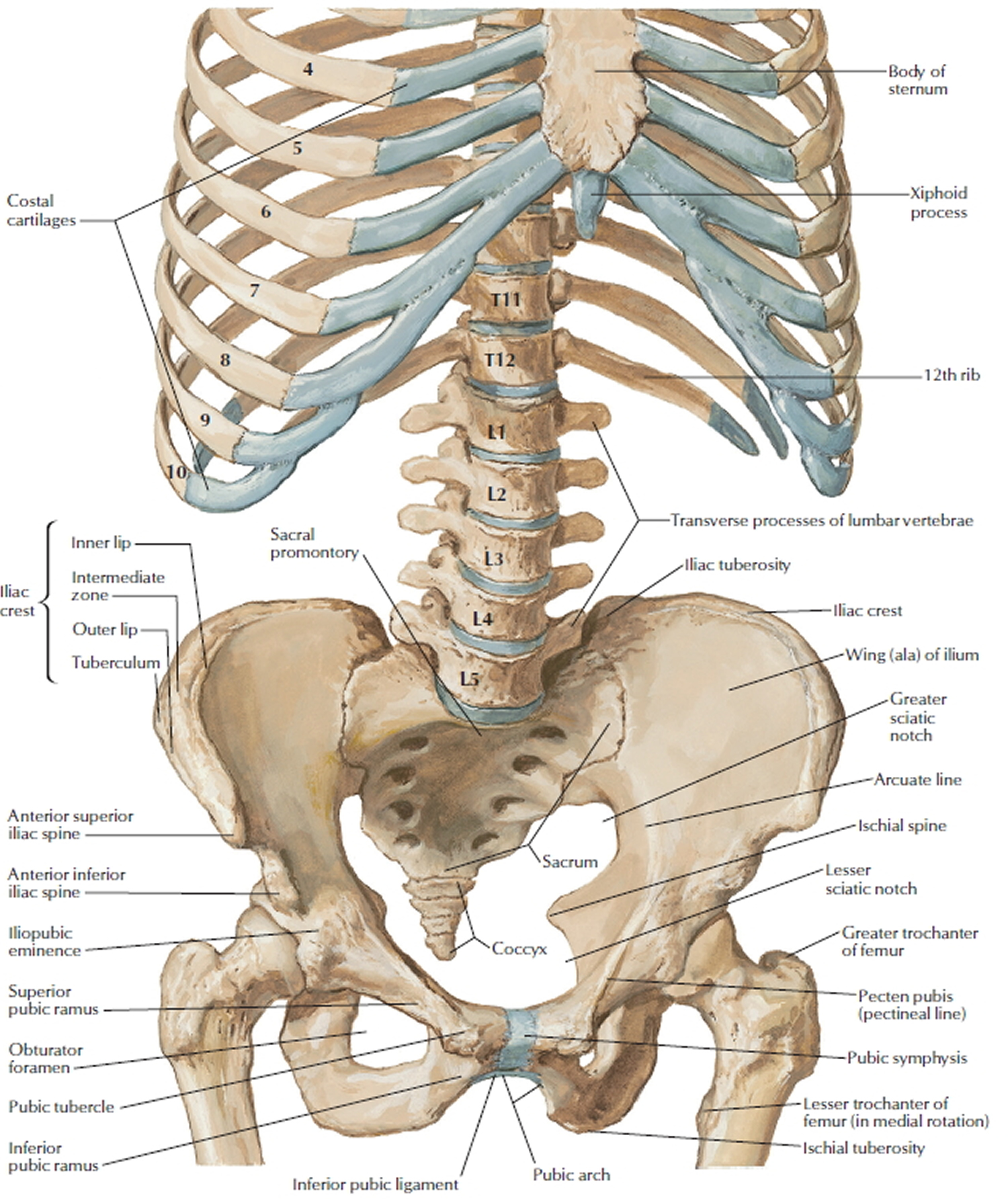
Figure 7. Hand and wrist bones

Figure 8. Foot bones

Structure and Function of Bone
Structure of Bone
A typical long bone consists of the following parts:
- The diaphysis is the bone’s shaft or body—the long, cylindrical, main portion of the bone.
- The epiphyses (singular is epiphysis) are the proximal and distal ends of the bone.
- The metaphyses (singular is metaphysis) are the regions between the diaphysis and the epiphyses. In a growing bone, each metaphysis contains an epiphyseal (growth) plate, a layer of hyaline cartilage that allows the diaphysis of the bone to grow in length. When a bone ceases to grow in length at about ages 14–24, the cartilage in the epiphyseal plate is replaced by bone; the resulting bony structure is known as the epiphyseal line.
- The articular cartilage is a thin layer of hyaline cartilage covering the part of the epiphysis where the bone forms an articulation (joint) with another bone. Articular cartilage reduces friction and absorbs shock at freely movable joints. Because articular cartilage lacks a perichondrium and lacks blood vessels, repair of damage is limited.
- The periosteum is a tough connective tissue sheath and its associated blood supply that surrounds the bone surface wherever it is not covered by articular cartilage. It is composed of an outer fibrous layer of dense irregular connective tissue and an inner osteogenic layer that consists of cells. Some of the cells enable bone to grow in thickness, but not in length. The periosteum also protects the bone, assists in fracture repair, helps nourish bone tissue, and serves as an attachment point for ligaments and tendons. The periosteum is attached to the underlying bone by perforating fibers or Sharpey’s fibers, thick bundles of collagen that extend from the periosteum into the bone extracellular matrix.
- The medullary cavity or marrow cavity, is a hollow, cylindrical space within the diaphysis that contains fatty yellow bone marrow and numerous blood vessels in adults. This cavity minimizes the weight of the bone by reducing the dense bony material where it is least needed. The long bones’ tubular design provides maximum strength with minimum weight.
- The endosteum is a thin membrane that lines the medullary cavity. It contains a single layer of bone-forming cells and a small amount of connective tissue.
Figure 9. Parts of a long bone
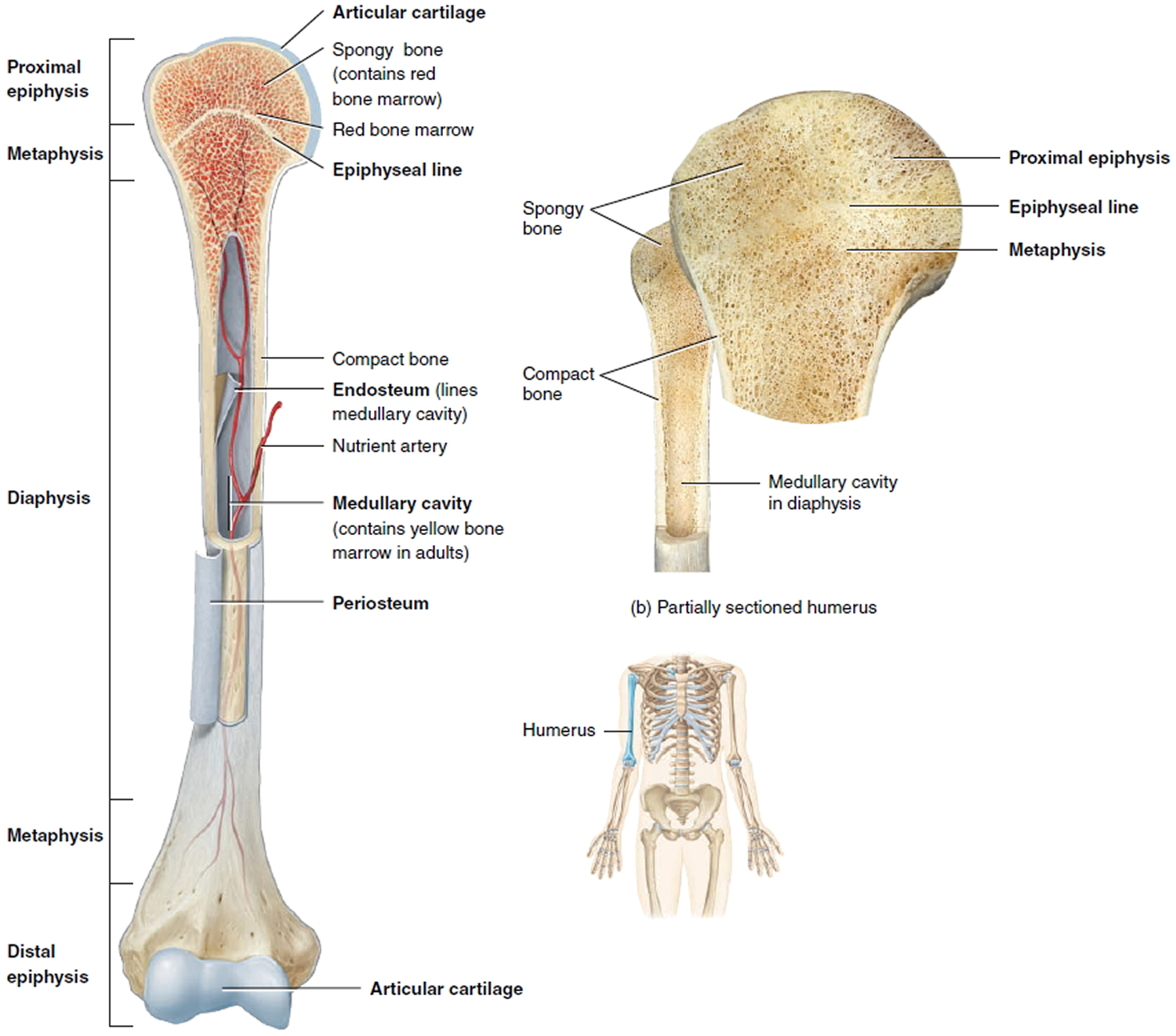
Note: The spongy bone tissue of the epiphyses and metaphyses contains red bone marrow, and the medullary cavity of the diaphysis contains yellow bone marrow (in adults).
The Cells of Mature Bone
Bone contains four cell types: osteoblasts, osteocytes, osteoprogenitor cells, and osteoclasts.
Osteocytes
Mature bone cells are osteocytes. They maintain and monitor the protein and mineral content of the surrounding matrix. The minerals in the matrix are continually recycled. Each osteocyte directs the release of calcium from bone into blood and the deposition of calcium salts into the surrounding matrix. Osteocytes occupy small chambers, called lacunae, that are sandwiched between layers of calcified matrix. These matrix layers are called lamellae (singular, lamella) (Figure 10 b–d). Channels called canaliculi (“little canals”) radiate through the matrix from lacuna to lacuna and toward free surfaces and adjacent blood vessels. The canaliculi connect adjacent lacunae and bring the processes of neighboring osteocytes into close contact. Tight junctions interconnect these processes and provide a route for the diffusion of nutrients and waste products from one osteocyte to another across gap junctions.
Osteoblasts
Cuboidal cells found in a single layer on the inner or outer surfaces of a bone are called osteoblasts (precursor). These cells secrete the organic components of the bone matrix. This material, called osteoid, later becomes mineralized through a complicated, multistep mechanism. Osteoblasts are responsible for the production of new bone—a process called osteogenesis. It is thought that osteoblasts respond to a variety of different stimuli, including mechanical and hormonal, to initiate osteogenesis. If an osteoblast becomes surrounded by matrix, it differentiates into an osteocyte.
Osteoprogenitor Cells
Bone tissue also contains small numbers of stem cells termed osteoprogenitor cells (ancestor). Osteoprogenitor cells differentiate from mesenchyme and are found in numerous locations, including the innermost layer of the periosteum and the endosteum lining the medullary cavities. Osteoprogenitor cells divide to produce daughter cells that differentiate into osteoblasts. The ability to produce additional osteoblasts becomes extremely important after a bone is cracked or broken.
Osteoclasts
Large, multinucleate cells found at sites where bone is being removed are termed osteoclasts. They are derived from the same stem cells that produce monocytes and neutrophils. They secrete acids through a process involving the exocytosis of lysosomes. The acids dissolve the bony matrix and release amino acids and the stored calcium and phosphate. This erosion process, called osteolysis, increases the calcium and phosphate concentrations in body fluids. Osteoclasts are always removing matrix and releasing minerals, and osteoblasts are always producing matrix that quickly binds minerals. The balance between the activities of osteoblasts and osteoclasts is very important; when osteoclasts remove calcium salts faster than osteoblasts deposit them, bones become weaker. When osteoblasts are more active than osteoclasts, bones become stronger and more massive. New research indicates that osteoclasts may also be involved in osteoblast
differentiation, immune system activation, and the proliferation of tumor cells in bone.
Figure 10. Microscopic Structure of a Typical Bone
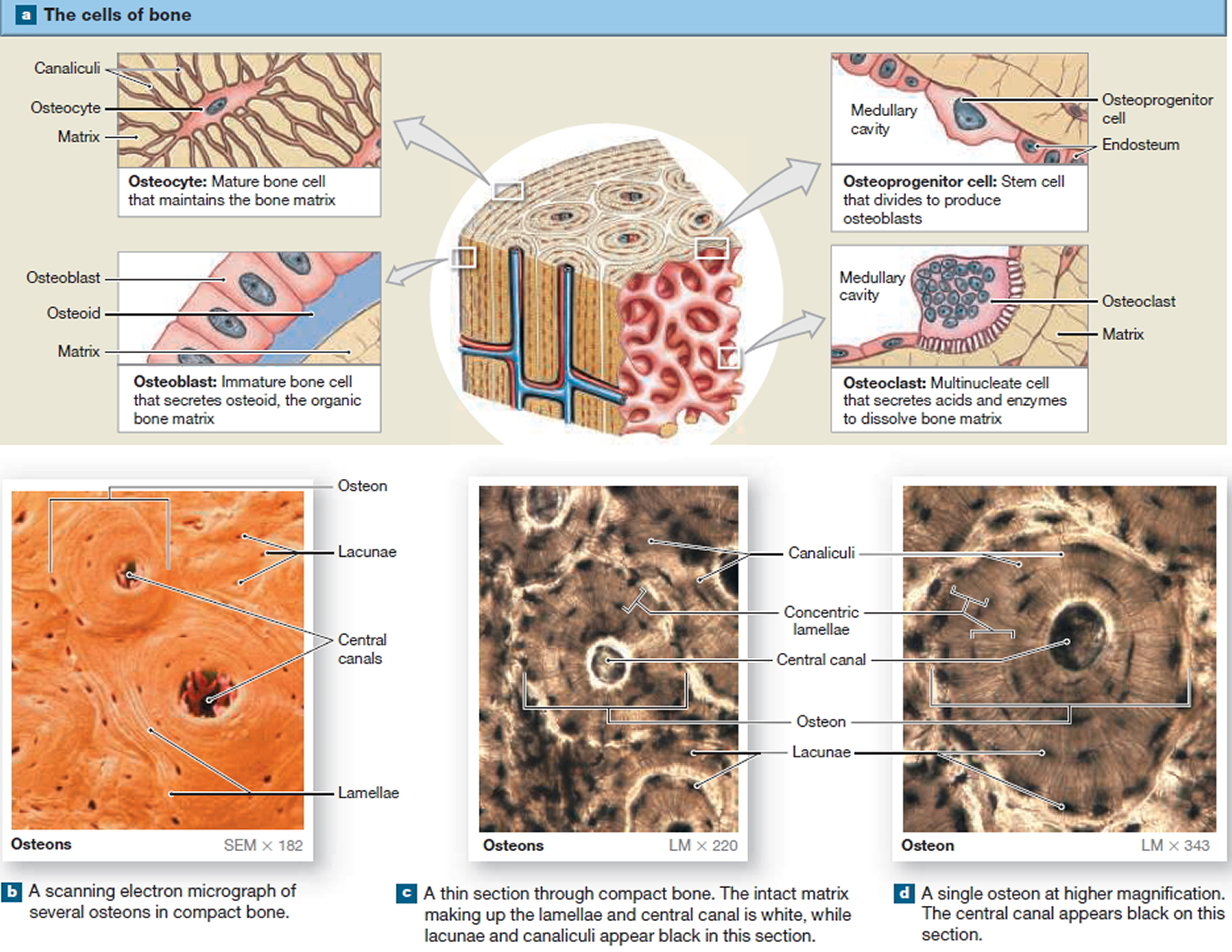
Organization of Mature Bone
The Matrix of Bone
Calcium phosphate, accounts for almost two-thirds of the weight of bone. It interacts with calcium hydroxide to form crystals of hydroxyapatite. As these crystals form, they incorporate other calcium salts, such as calcium carbonate, and ions such as sodium, magnesium, and fluoride. This process, called calcification is initiated by bone-building cells called osteoblasts. Calcification requires the presence of collagen fibers. Mineral salts first begin to crystallize in the microscopic spaces between collagen fibers. After the spaces are filled, mineral crystals accumulate around the collagen fibers. The combination of crystallized salts and collagen fibers is responsible for the characteristics of bone. These inorganic components enable bone to resist compression. Roughly one-third of the weight of bone is from collagen fibers and other noncollagenous proteins, which give bone considerable tensile strength.
Osteocytes and other cell types account for only 2 percent of the mass of a typical bone. Calcium phosphate crystals are very strong, but relatively inflexible. They with stand compression, but the crystals shatter when exposed to bending, twisting, or sudden impacts. Collagen fibers are tough and flexible. They easily tolerate stretching, twisting, and bending but, when compressed, they simply bend out of the way. In bone, the collagen fibers and other noncollagenous proteins provide an organic framework for the formation of mineral crystals. The hydroxyapatite crystals form small plates that lie alongside these ground substance proteins. The result is a protein–crystal combination with properties intermediate between those of collagen and those of pure mineral crystals.
Compact and Spongy Bone
There are two types of osseous tissue: compact bone and spongy bone. Compact bone is relatively dense and solid, whereas spongy bone, also termed trabecular bone or cancellous bone, forms an open network of struts and plates. Both are found in typical bones of the skeleton, such as the humerus, the proximal bone of the upper limb, and the femur, the proximal bone of the lower limb. Compact bone forms the walls, and an internal layer of spongy bone surrounds the medullary
(marrow) cavity. The medullary cavity contains bone marrow, a loose connective tissue that is dominated by either adipocytes (yellow marrow) or a mixture of mature and immature red and white blood cells and the stem cells that produce them (red marrow). Yellow marrow, often found in the medullary cavity of the shaft, is an important energy reserve. Extensive areas of red marrow, such as in the spongy bone of the femur, are important sites of blood cell formation.
Structural Differences between Compact and Spongy Bone
Compact and spongy bone have the same matrix composition, but they differ in the three-dimensional arrangement of the osteocytes, canaliculi, and lamellae.
Figure 11. Internal organization of bones

Compact Bone
Compact bone tissue contains few spaces and is the strongest form of bone tissue. It is found beneath the periosteum of all bones and makes up the bulk of the diaphyses of long bones. Compact bone tissue provides protection and support and resists the stresses produced by weight and movement.
Compact bone tissue is composed of repeating structural units called osteons, or haversian systems. Each osteon consists of concentric lamellae arranged around an osteonic (haversian or central) canal. Resembling the growth rings of a tree, the concentric lamellae are circular plates of mineralized extracellular matrix of increasing diameter, surrounding a small network of blood vessels and nerves located in the central canal. These tubelike units of bone generally form a series of parallel cylinders that, in long bones, tend to run parallel to the long axis of the bone. Between the concentric lamellae are small spaces called lacunae (little lakes; singular is lacuna), which contain osteocytes. Radiating in all directions from the lacunae are tiny canaliculi (small channels), which are filled with extracellular fluid. Inside the canaliculi are slender fingerlike processes of osteocytes.
Neighboring osteocytes communicate via gap junctions. The canaliculi connect lacunae with one another and with the central canals, forming an intricate, miniature system of interconnected canals throughout the bone. This system provides many routes for nutrients and oxygen to reach the osteocytes and for the removal of wastes.
Osteons in compact bone tissue are aligned in the same direction and are parallel to the length of the diaphysis. As a result, the shaft of a long bone resists bending or fracturing even when considerable force is applied from either end. Compact bone tissue tends to be thickest in those parts of a bone where stresses are applied in relatively few directions. The lines of stress in a bone are not static. They change as a person learns to walk and in response to repeated strenuous physical activity, such as weight training. The lines of stress in a bone also can change because of fractures or physical deformity. Thus, the organization of osteons is not static but changes over time in response to the physical demands placed on the skeleton.
The areas between neighboring osteons contain lamellae called interstitial lamellae, which also have lacunae with osteocytes and canaliculi. Interstitial lamellae are fragments of older osteons that have been partially destroyed during bone rebuilding or growth. Blood vessels and nerves from the periosteum penetrate the compact bone through transverse interosteonic (Volkmann’s or perforating) canals. The vessels and nerves of the interosteonic canals connect with those of the medullary cavity, periosteum, and central canals. Arranged around the entire outer and inner circumference of the shaft of a long bone are lamellae called circumferential lamellae. They develop during initial bone formation. The circumferential lamellae directly deep to the periosteum are called external circumferential lamellae. They are connected to the periosteum by perforating (Sharpey’s) fibers. The circumferential lamellae that line the medullary cavity are called internal circumferential lamellae.
Spongy Bone
The major difference between compact and spongy bone is the arrangement of spongy bone into parallel struts or thick, branching plates called trabeculae (also termed spicules). Numerous interconnecting spaces occur between the trabeculae in spongy bone. Spongy bone possesses lamellae, and if the trabeculae are thick enough, osteons will be present. In terms of the associated cells and the structure and composition of the lamellae, spongy bone is no different from compact bone. Spongy bone forms an open framework and as a result it is much lighter than compact bone. However, the branching trabeculae give spongy bone considerable strength despite its relatively light weight. Thus, the presence of spongy bone reduces the weight of the skeleton and makes it easier for muscles to move the bones. Spongy bone is found wherever bones are not stressed heavily or where stresses arrive from many directions.
Functional Differences between Compact and Spongy Bone
A layer of compact bone covers the surface of all bones. The thickness of this layer varies from region to region and from one bone to another, but compact bone is thickest where stresses arrive from a limited number of directions. This superficial layer of compact bone is in turn covered by the periosteum, a connective tissue wrapping that is connected to the deep fascia. The periosteum is complete everywhere except within a joint, where the edges or ends of two bones contact one another.
The shaft of compact bone transfers stresses from one epiphysis to another. For example, when you are standing, the shaft of the femur transfers your body weight from your hip to your knee. The osteons within the shaft are parallel to its long axis, and as a result the femur is very strong when stressed along that axis. Imagine a single osteon as a drinking straw with very thick walls. When you push the ends of a straw together, it seems quite strong. However, when you hold the ends and push the side of the straw, it breaks easily. Similarly, a long bone does not bend when forces are applied to either end, but an impact to the side of the shaft can easily cause a break, or fracture.
Spongy bone is not as massive as compact bone, but it is much more capable of resisting stresses applied from many different directions. The epiphyses of the femur are filled with spongy bone, and the alignment of the trabeculae within the proximal epiphysis. The trabeculae are oriented along the stress lines, but with extensive cross-bracing. At the proximal epiphysis, the trabeculae transfer forces from the hip across the metaphysis to the femoral shaft; at the distal epiphysis, the trabeculae transfer the forces across the knee joint to the leg. In addition to reducing weight and handling stress from many directions, the open trabecular framework supports and protects the cells of the bone marrow.
Blood and Nerve Supply to Bones
Like any living tissue, bones need nourishment. Osseous tissue is very vascular, and the bones of the skeleton have an extensive blood supply. In a long bone such as the humerus, four major sets of blood vessels develop:
- The nutrient artery and nutrient vein: These vessels form as blood vessels invade the cartilage model at the start of endochondral ossification. There is usually only one nutrient artery and one nutrient vein entering the diaphysis through a nutrient foramen. A foramen (plural, foramina) is an opening in a bone. However, a few bones, including the femur, have two or more nutrient arteries. These vessels penetrate the shaft to reach the medullary cavity. The nutrient artery divides into ascending and descending branches, which approach the epiphyses. These vessels then re-enter the compact bone through perforating canals and extend along the central canals to supply the osteons of the compact bone.
- Metaphyseal vessels: These vessels supply blood to the inner (diaphyseal) surface of each epiphyseal cartilage, where bone is replacing cartilage.
- Epiphyseal vessels: The epiphyseal ends of long bones contain numerous smaller foramina. Epiphyseal vessels enter the bone through these foramina to supply the osseous tissue and medullary cavities of the epiphyses.
- Periosteal vessels: Blood vessels from the periosteum are incorporated into the developing bone surface. These vessels provide blood to the superficial osteons of the shaft. During endochondral ossification, branches of periosteal vessels enter the epiphyses, bringing blood to the secondary ossification centers. After the epiphyses close, all of these blood vessels become extensively interconnected. The periosteum also contains an extensive network of lymphatic vessels and sensory nerves. The lymphatic vessels collect lymph (fluid derived from the interstitial fluid) from branches that enter the bone and reach individual osteons through perforating canals. The sensory nerves penetrate the compact bone with the nutrient artery to innervate the endosteum, medullary cavity, and epiphyses. Because of this rich sensory innervation, injuries to bones are usually very painful.
Figure 12. Blood supply to a Mature Bone
