Fagocito

Os fagocitos son células que protexen o corpo ao inxeriren partículas estrañas daniñas, bacterias e células mortas ou moribundas. O seu nome vén do grego phagein, 'comer', e "-cito", o sufixo que en bioloxía significa 'célula', procedente do grego kutos, 'recipiente oco'.[1] Son esenciais para loitar contra as infeccións e ter inmunidade.[2] Os fagocitos son importantes en todos os grupos do reino animal[3] e están moi desenvolvidos nos vertebrados.[4] Un litro de sangue humano contén uns seis mil millóns de fagocitos.[5] Descubriunos en 1882 Ilya Ilyich Mechnikov mentres estaba estudando larvas de estrela de mar.[6] Mechnikov recibiu en 1908 o Premio Nobel de Medicina ou Fisioloxía polo seu descubrimento.[7] Os fagocitos aparecen en moitas especies, incluso en microorganismos; algunhas amebas compórtanse como fagocitos macrófagos, o cal suxire que os fagocitos apareceron moi cedo na evolución da vida.[8]
Os fagocitos humanos e doutros animais denomínanse "profesionais" ou "non profesionais" dependendo da súa efectividade á hora de realizar a fagocitose.[9] Os fagocitos profesionais inclúen moitos tipos de leucocitos, como neutrófilos, monocitos, macrófagos, mastocitos e células dendríticas.[10] A principal diferenza entre os fagocitos profesionais e non profesionais é que os fagocitos profesionais teñen na súa superficie moléculas chamadas receptores que poden detectar obxectos daniños, como as bacterias, que non se encontran normalmente no corpo.[11] Os fagocitos son cruciais na loita contra as infeccións e para manter os tecidos sans ao eliminaren as células mortas e moribundas que chegaron ao final da súa vida.[12]
Durante unha infección, os sinais químicos atraen os fagocitos a lugares onde os patóxenos invadiron o corpo. Estes compostos químicos poden proceder das bacterias invasoras ou doutros fagocitos xa presentes. Os fagocitos móvense por medio da quimiotaxe. Cando os fagocitos entran en contacto cunha bacteria, os receptores da superficie dos fagocitos únense a ela. Esta unión causará a fagocitación da bacteria polo fagocito.[13] Algúns fagocitos matan os patóxenos inxeridos con oxidantes e óxido nítrico.[14] Despois da fagocitose, os macrófagos e células dendríticas poden tamén participar na presentación de antíxenos, un proceso no cal o fagocito despraza fragmentos do material inxerido de novo á súa superficie. Este material funciona como un antíxeno e é mostrado a outras células do sistema inmunitario. Algúns fagocitos viaxan despois aos ganglios linfáticos do corpo e mostran este material a un tipo de glóbulos brancos chamados linfocitos. Este proceso é importante para construír a inmunidade,[15] e en moitos patóxenos evolucionaron métodos para evadirse do ataque dos fagocitos.[2]
Historia
[editar | editar a fonte]
O zoólogo ruso Ilya Ilyich Mechnikov (1845–1916) foi o primeiro que recoñeceu que células especializadas estaban implicadas na defensa contra infeccións microbianas.[16] En 1882, estudou as células móbiles das larvas de estrela de mar, crendo que eran importantes para as defensas inmunes do animal. Para comprobar esta idea, inseriu nas larvas pequenas espiñas da árbore das mandarinas. Pasadas unhas horas decatouse de que as células móbiles rodearan as espiñas.[16] Mechnikov viaxou a Viena e compartiu as súas ideas con Carl Friedrich Claus, que propuxo o nome de "fagocito" (do grego 'célula que come')[1] para as células observadas por Mechnikov.[17]
Un ano despois, Mechnikov estudou o crustáceo de auga doce Daphnia, un diminuto animal transparente que podía ser examinado directamente baixo o microscopio. Descubriu que as esporas fúnxicas que atacaban o animal eran destruídas por fagocitos. Continuou ampliando as súas observacións co estudo dos glóbulos brancos dos animais e descubriu que a bacteria Bacillus anthracis podía ser englobada e matada polos fagocitos, un proceso que el chamou fagocitose.[18] Mechnikov propuxo que os fagocitos eran unha defensa primaria contra organismos invasores.[16]
En 1903, Almroth Wright descubriu que a fagocitose era reforzada por anticorpos específicos que chamou opsoninas, do grego opson, 'aderezo, prebe'.[19] Mechnikov foi galardoado (conxuntamente con Paul Ehrlich) co Premio Nobel de Medicina ou Fisioloxía de 1908 polo seu traballo sobre os fagocitos e a fagocitose.[7]
Aínda que a importancia destes descubrimentos foi lentamente recoñecida durante os inicios do século XX, as intricadas relacións entre os fagocitos e os demais compoñentes do sistema inmunitario non se coñeceron ata a década de 1980.[20]
Fagocitose
[editar | editar a fonte]- Artigo principal: Fagocitose.

A fagocitose é o proceso de captación e introdución na célula de partículas como bacterias, parasitos, células mortas do hóspede e residuos celulares e alleos realizado por unha célula.[21] Implica unha cadea de procesos moleculares.[22] A fogacitose ocorre unha vez que un corpo estraño, unha célula bacteriana, por exemplo, se uniu a moléculas chamadas "receptores" que están na superficie dos fagocitos. O fagocito despois esténdese arredor da bacteria e englóbaa. A fagocitose de bacteria polos neutrófilos humanos tarda como media uns nove minutos.[23] Unha vez dentro do fagocito, a bacteria queda atrapada nun compartimento chamado fagosoma. En aproximadamente un minuto o fagosoma fusiónase cun lisosoma ou un gránulo para formar un fagolisosoma. A bacteria é despois sometida a un poderoso conxunto de mecanismos que a matan[24] en poucos minutos.[23] As células dendríticas e os macrófagos non son tan rápidos e neles a fagocitose pode tardar moitas horas. Os macrófagos son comedores lentos e sucios; engloban grands cantidades de material e liberan frecuentemente algo de material indixerido nos tecidos. Os residuos serven como sinal para recrutar máis fagocitos que chegan desde o sangue.[25] Os fagocitos teñen un apetito voraz; en experimentos alimentáronse macrófagos con limaduras de ferro e despois utilizouse un pequeno imán para separalos doutras células.[26]

Un fagocito ten moitos tipos de receptores sobre a súa superficie que se utilizan para a unión dos materiais.[2] Inclúen receptores de opsoninas, receptor limpador (scavenger) e receptores de tipo Toll. Os receptores de opsoninas incrementan a fagocitose de bacterias que foron cubertas con anticorpos inmunoglobulina G (IgG) ou con moléculas do sistema de complemento. O "complemento" é o nome que se lle dá a unha complexa serie de moléculas proteicas que se encontran no sangue e que destrúen células ou márcanas para a súa destrución.[27] Os receptores limpadores únense a unha ampla variedade de moléculas da superficie das céulas bacterianas, e os receptores de tipo Toll, así chamados pola súa semellanza cos receptores ben estudados das moscas do vinagre codificados polo xene Toll, únense a moléculas máis específicas. A unión a receptores de tipo Toll incrementa a fagocitose e causa a liberación por parte do fagocito dun grupo de hormonas que causa inflamación.[2]
Métodos de matar os microbios
[editar | editar a fonte]
Matar os microbios é unha función esencial dos fagocitos[28] que se realiza no interior do fagocito (morte intracelular) ou no exterior (extracelular).
Intracelular dependente do oxíxeno
[editar | editar a fonte]Cando un fagocito inxire unha bacteria (ou calquera outro material), o seu consumo de oxíxeno increméntase. O incremento no consumo de oxíxeno, chamado explosión respiratoria, produce moléculas que conteñen oxíxeno que son antimicrobianas.[29] Estes compostos de oxíxeno (especies reactivas do oxíxeno) son tóxicas tanto para o invasor coma para a propia célula fagocítica, polo que son mantidas dentro de compartimentos ben illados dentro da célula. Este método de matar microbios invasores usando moléculas reactivas do oxíxeno denomínase morte intracelular dependente do oxíxeno, da cal hai dous tipos.[14]
O primeiro tipo é a produción dependente do oxíxeno de superóxidos,[2] os cales son substancias ricas en oxíxeno letais para as bacterias.[30] O superóxido convértese en peróxido de hidróxeno e oxíxeno singulete por acción dun encima chamado superóxido dismutase. Os superóxidos tamén reaccionan co peróxido de hidróxeno para producir radicais hidroxilo, os cales tamén axudan a matar os microbios invasores.[2]
O segundo tipo implica o uso do encima mieloperoxidase contido nos gránulos dos neutrófilos.[31] Cando os gránulos se fusionan cun fagosoma, libérase a mieloperoxidase no fagolisosoma, e este encima utiliza peróxido de hidróxeno e cloro para crear hipoclorito, unha substancia usada na lixivia doméstica. O hipoclorito é extremadamente tóxico para as bacterias.[2] A mieloperoxidase contén un pigmento hemo, que é o que determina que as secrecións ricas en neutrófilos teñan cor verde, tales como a pus e os esputos infectados.[32]
Intracelular independente do oxíxeno
[editar | editar a fonte]
Os fagocitos poden tamén matar microbios por métodos independentes do oxíxeno, mais estes non son tan efectivos coma os dependentes do oxíxeno. Hai catro tipos principais. O primeiro usa proteínas cargadas electricamente que danan a membrana plasmática da bacteria. O segundo tipo usa lisocimas; estes encimas degradan a parede celular bacteriana. O terceiro tipo usa lactoferrinas, que están presentes en gránulos de neutrófilos e eliminan ferro esencial para as bacterias.[33] O cuarto tipo utiliza proteases e encimas hidrolíticas; estes encimas utilízanse para dixerir as proteínas das bacterias destruídas.[34]
Extracelular
[editar | editar a fonte]O interferón-gamma, que era chamado antes factor activador de macrófagos, estimula os macrófagos para que produzan óxido nítrico. A fonte de interferón-gamma poden ser células T CD4+, células T CD8+, céllas asasinas naturais, células B, células T asasinas naturais, monocitos, macrófagos ou células dendríticas.[35] Entón libérase o óxido nítrico dos macrófagos e, debido á súa toxicidade, mata os microbios que están próximos ao macrófago.[2] Os macrófagos activados producen e segregan factor de necrose tumoral. Esta citocina (unha clase de molécula de sinalización)[36] mata células canceríxenas e células infectadas por virus, e axuda a activar outras células do sistema inmunitario.[37]
Nalgunhas doenzas, por exemplo a pouco frecuente enfermidade granulomatosa crónica, a eficiencia dos fagocitos está moi diminuída e as infeccións bacterianas recorrentes son un constante problema.[38] Nesta doenza preséntanse anormalidades que afectan a diferentes elementos do mecanismo de morte dependente do oxíxeno. Outras anormalidades conxénitas raras, como a síndrome de Chédiak–Higashi, están tamén asociadas con defectos no mecanismo para matar os microbios inxeridos.[39]
Virus
[editar | editar a fonte]Os virus só poden reproducirse dentro das células, e poden conseguir entrar nelas utilizando moitos dos receptores que interveñen na inmunidade. Unha vez dentro da célula, os virus usan a maquinaria biolóxica da célula no seu propio beneficio, forzando a célula a que faga centos de copias idénticas deles. Aínda que os fagocitos e outros compoñentes do sistema inmunitario innato poden, ata certo punto, controlar os virus, unha vez que o virus está dentro da célula as respostas inmunitarias adaptativas, especialmente a dos linfocitos, son máis importantes para a defensa.[40] No lugar onde hai infeccións virais, os linfocitos a miúdo superan amplamente en número a outras células do sistema inmunitario; isto é común, por exemplo, na meninxite viral.[41] As células infectadas por virus que os linfocitos mataron son limpadas do lugar polos fagocitos.[42]
Papel na apoptose
[editar | editar a fonte]- Artigo principal: Apoptose.

Nun anmal as células están morrendo constantemente. Hai un equilibrio entre a división celular e a morte celular que mantén relativamente constante o número de células no organismo adulto.[12] Unha célula pode morrer de dúas maneiras: por necrose ou por apoptose. A diferenza da necrose, que adoita ser o resultado dunha enfermidade ou trauma, a apoptose (ou a morte celular programada) é unha función normal e sá das céllas. O corpo debe eliminar millóns de células mortas ou moribundas cada día e os fagocitos desempeñan un papel crucial neste proceso.[43]
As células que están morrendo que pasan polas fases finais da apoptose[44] exhiben moléculas, como a fosfatidilserina, na súa superficie celular que atraen os fagocitos.[45] A fosfatidilserina atópase normalmente na superficie citosólica da membrana plasmática, pero é redistribuída durante a apoptose por unha proteína chamada escramblase (scramblase) e aparece na superficie extracelular.[46][47] Estas moléculas marcan a célula para a fagocitose que realizan as células que posúan os receptores axeitados, como os macrófagos.[48] A eliminación de células moribundas que fan os fagocitos ocorre de maneira ordenada sen causar unha resposta inflamatoria e é unha función importante dos fagocitos.[49]
Interaccións con outras células
[editar | editar a fonte]Os fagocitos non están normalmente asociados a ningún órgano do corpo en particular, senón que se moven polo corpo interaccionando con outros fagocitos e con células non fagocíticas do sistema inmunitario. Poden comunicarse con outras células producindo substancias químicas chamadas citocinas, que serven para recrutar outros fagocitos no sitio da infección ou estimular linfocitos dormentes.[50] Os fagocitos forman parte do sistema inmunitario innato, que posúen os animais, incluídos os humanos. A inmunidade innata é moi efectiva pero non específica, xa que non discrimina entre diferentes tipos de invasores. Por outra parte, o sistema inmunitario adaptativo dos vertebrados con mandíbula (no que se basea a inmunidade adquirida) está moi especializado e pode protexer contra case calquera tipo de invasor.[51] O sistema inmunitario adaptativo non depende dos fagocitos senón dos linfocitos, os cales producen proteínas protectoras chamadas anticorpos, que etiquetan os invasores para a súa destrución e impiden que os virus infecten as células.[52] Os fagocitos, concretamente as células dendríticas e macrófagos, estimulan os linfocitos para que produzan anticorpos por un importante proceso chamado presentación de antíxenos.[53]
Presentación de antíxenos
[editar | editar a fonte]- Artigo principal: Presentación de antíxenos.

A presentación de antíxenos é un proceso no cal algúns fagocitos volven levar partes do material que fagocitaron de novo á superficie celular e "preséntanas" a outras células do sistema inmunitario.[54] Hai dúas células presentadoras de antíxenos "profesionais": macrófagos e células dendríticas.[55] Despois de que os materiais son englobados e introducidos na célula, as proteínas estrañas (os antíxenos) son cortadas en péptidos dentro das células dendríticas e macrófagos. Estes péptidos únense despois a glicoproteínas do complexo maior de histocompatibilidade (MHC) da célula, as cales transportan os péptidos á superficie celular do fagocito, onde poden ser mostradas ("presentadas") aos linfocitos.[15] Os macrófagos maduros non viaxan moi lonxe do sitio de infección, pero as células dendríticas poden chegar aos ganglios linfáticos do corpo, onde hai millóns de linfocitos.[56] Isto potencia a inmunidade porque os linfocitos responden aos antíxenos presentados por ditas células igual que se estivesen no sitio da infección inicial.[57] Pero as células dendríticas poden tamén destruír ou pacificar os linfocitos se eles recoñecen compoñentes do corpo do hóspede; isto é necesario para impedir as reaccións autoinmunes. Este proceso denomínase tolerancia.[58]
Tolerancia inmunolóxica
[editar | editar a fonte]- Artigo principal: Tolerancia inmunolóxica.
As células dendríticas tamén promoven a tolerancia inmunolóxica,[59] que impide que o corpo se ataque a si mesmo. O primeiro tipo de tolerancia é a tolerancia central, que ocorre no timo. As células T que se unen (por medio do seu receptor de células T) demasiado fortemente a antíxenos propios (presentados por células dendríticas sobre moléculas MHC) son inducidos á morte. O segundo tipo de tolerancia inmunolóxica é a tolerancia periférica. Algunhas células T autorreactivas escapan do timo sen ser destruídas por diferentes razóns, principalmente debido á falta de expresión dalgúns antíxenos propios no timo. Outro tipo de célula T, as células T reguladoras poden regular á baixa as células T autorreactivas na periferia.[60] Cando falla a tolerancia inmunolóxica, poden orixinarse enfermidades autoinmunes.[61]
Fagocitos profesionais
[editar | editar a fonte]
Os fagocitgos humanos e doutros vertebrados mandibulados divídense en dous grupos: "profesionais" e "non profesionais" en canto á súa eficiencia á hora de realizar a fagocitose.[9] Os fagocitos profesionais son os monocitos, macrófagos, neutrófilos, células dendríticas dos tecidos e mastocitos.[10] Un litro de sangue humano contén uns seis mil millóns de fagocitos.[5]
Activación
[editar | editar a fonte]Todos os fagocitos, e moi especialmente os macrófagos, poden presentar distintos graos de rapidez para actuar. Os macrófagos adoitan estar relativamente dormentes nos tecidos e proliferan lentamente. Neste estado de semi-repouso, poden eliminar as células mortas do hóspede e outros residuos non infecciosos e raramente toman parte na presentación de antíxenos. Mais, durante unha infección, reciben sinais químicos, xeralmente o interferón-gamma, que fan que incrementen a súa produción de moléculas de MHC II e que os preparan para a presentación de antíxenos. Neste estado, os macrófagos son bos presentadores de antíxenos e matadores de células. Porén, se reciben un sinal directamente dun invasor, quedan "hiperactivados", deixan de proliferar e concéntranse en matar células. O seu tamaño e taxa de fagocitose increméntase, e algúns fanse o suficientemente grandes como para fagocitar protozoos invasores.[62]
No sangue, os neutrófilos está inactivos, pero son arrastrados a gran velocidade. Cando reciben sinais dos macrófagos situados no sitio de inflamación, van máis lentos e abandonan o torrente sanguíneo. Unha vez que pasan aos tecidos, son activados por citocinas e chegan ao campo de batalla listos para matar.[63]
Migración
[editar | editar a fonte]
Cando ocorre unha infección, emítese un sinal químico de "SOS" para atraer fagocitos a ese lugar.[64] Estes sinais químicos poden incluír proteínas de bacterias invasoras, péptidos do sistema de coagulación, produtos do complemento e citocinas que foron liberadas polos macrófagos localizados no tecido preto do lugar da infección.[2] Outro grupo de atractores químicos son as citocinas que recrutan neutrófilos e monocitos que se encontraban no sangue.[13]
Para chegaren ao sitio de infección, os fagocitos abandonan a circulación sanguínea e entran nos tecidos afectados. Sinais procedentes da infección causan que as células endoteliais que tapizan os vasos sanguíneos fabriquen unha proteína chamada selectina, á cal se unen os neutrófilos que pasan. Outros sinais chamados vasodilatadores afrouxan as unións que conectan as células endoteliais, o que permite que os fagocitos pasen a través da parede do capilar. A quimiotaxe é o proceso polo cal os fagocitos seguen o "rastro" de citocinas ata o lugar da infección.[2] Os neutrófilos viaxan a través de órganos tapizados por células epiteliais aos sitios de infección, e, aínda que este é un compoñente importante da loita contra a infección, a propia migración pode orixinar síntomas similares aos dunha enfermidade.[65] Durante unha infección, recrútanse millóns de neutrófilos desde o sangue, pero morren en poucos días.[66]
Monocitos
[editar | editar a fonte]- Artigo principal: Monocito.

Os monocitos desenvóvlense na medula ósea e chegan á madureza no sangue. Os monocitos maduros teñen núcleos grandes, lisos e lobados e abundante citoplasma que contén gránulos. Os monocitos inxiren substancias alleas ou perigosas e presentan antíxenos a outras células inmunitarias. Forman dous grupos: monocitos circulantes e monocitos marxinais, que permanecen noutros tecidos (os marxinais son aproximadamente o 70% de todos os monocitos). A maioría dos monocitos abandonan a circulación sanguínea despois de 20–40 horas para viaxar aos tecidos e órganos e ao facelo transfórmanse en macrófagos[67] ou en células dendríticas, dependendo dos sinais que reciban.[68] Hai uns 500 millóns de monocitos nun litro de sangue humano.[5]
Macrófagos
[editar | editar a fonte]- Artigo principal: Macrófago.
Os macrófagos maduros non viaxan moi lonxe senón que permanecen facendo garda naquelas áreas do corpo que están expostas ao mundo exterior. Alí actúan como colectores de lixo, células presentadoras do antíxeno ou matadores feroces de células, dependendo dos sinais que reciban.[69] Derivan de monocitos, de células nais de granulocitos ou da división de macrófagos preexistentes.[70] Os macrófagos humanos teñen uns 21 micrómetros de diámetro.[71]

Este tipo de fagocitos non ten gránulos, pero contén moitos lisosomas. Os macrófagos encóntranse por todo o corpo en case todos os tecidos e órganos (por exemplo, as células microgliais do cerebro e os macrófagos alveolares dos pulmóns), onde se encontran esperando sixilosamente. A localización dun macrófago pode determinar o seu tamaño e aparencia. Os macrófagos causan inflamación por medio da produción de interleucina-1, interleucina-6 e TNF-alfa.[72] Os macrófagos adoitan encontrarse só nos tecidos e raramente se ven no sangue circulante. A duración da vida dun macrófago dos tecidos estimouse en de catro a quince días.[73]
Os macrófagos poden ser activados para que realicen funcións que os monocitos non poden realizar.[72] As células T axudantes (tamén chamadas células T efectoras ou células Th), un subgrupo de linfocitos, son responsables da activación dos macrófagos. As células Th1 activan os macrófagos ao enviar sinais con IFN-gamma e expoñer na súa superficie a proteína ligando de CD40.[74] Outros sinais son o TNF-alfa e os lipopolisacáridos bacterianos.[72] As células Th1 poden recrutar outros fagocitos ao sitio de infección de diversos xeitos. Segregan citocinas que actúan sobre a medula ósea para estimular a produción de monocitos e neutrófilos, e segregan algunhas das citocinas que son responsables da migración de monocitos e neutrófilos fóra da corrente sanguínea.[75] As células Th1 proceden da diferenciación de células T CD4+ unha vez que estas responderon ao antíxeno nos tecidos linfoides secundarios.[72] Os macrófagos activados teñen un potente papel na destrución de tumores ao produciren TNF-alfa, IFN-gamma, óxido nítrico, compostos reactivos do oxíxeno, proteínas catiónicas e encimas hidrolíticos.[72]
Neutrófilos
[editar | editar a fonte]- Artigo principal: Neutrófilo.

Os neutrófilos encóntranse normalmente na corrente sanguínea e son o tipo máis abundante de fagocitos, constituíndo do 50% ao 60% do total dos glóbulos brancos circulantes no sangue.[76] Un litro de sangue humano contén uns cinco mil millóns de neutrófilos,[5] os cales teñen uns 10 micrómetros de diámetro[77] e viven só uns cinco días.[37] Unha vez que reciben os sinais apropiados, tardan uns trinta minutos en abandonar o sangue e chegar ao sitio da infección.[78] Son comedores feroces e fagocitan rapidamente os invasores que están cubertos de anticorpos e moléculas do complemento, e células danadas ou residuos celulares. Os neutrófilos non regresan ao sangue; morren e convértense en células do pus.[78] Os neutrófilos maduros son menores en tamaño que os monocitos e teñen un núcleo segmentado con varios lobos ou seccións en número de 2 a 5; cada unha destas seccións está conectada por filamentos de cromatina. Os neutrófilos non se encontran normalmente na medula ósea ata a madureza, pero durante unha infección libéranse os precursores de neutrófilos chamados promielocitos, mielocitos e metamielocitos.[79]
Os gránulos intracelulares dos neutrófilos humanos foron desde hai longo tempo recoñecidos polas súas propiedades de destruír proteínas e bactericidas.[80] Os neutrófilos poden segregar produtos que estimulan os monocitos e macrófagos. As secrecións dos neutrófilos incrementan a fagocitose e a formación de compostos reactivos do oxíxeno que interveñen na matanza intracelular de células.[81] As secrecións dos gránulos primarios dos neutrófilos estimulan a fagocitose de bacterias cubertas con anticorpos IgG.[82]
Células dendríticas
[editar | editar a fonte]- Artigo principal: Célula dendrítica.

As células dendríticas son células presentadoras de antíxenos especializadas que posúen longas prolongacións celulares chamadas dendritas,[83] que lles axudan a fagocitar os microbios e outros invasores.[84][85] Están presentes nos tecidos que están en contacto co ambiente externo, principalmente na pel, na cuberta interna das fosas nasais, os pulmóns, o estómago e os intestinos.[86] Unha vez activadas, maduran e migran aos tecidos linfoides, onde interaccionan coas células T e B para iniciar e orquestrar a resposta inmunitaria adaptativa.[87] As células dendríticas maduras activan as células T axudantes e as células T citotóxicas.[88] As células T axudantes activadas, á súa vez, interaccionan con macrófagos e células B para activalas. Ademais, as células dendríticas poden influír no tipo de resposta inmune xerada; cando viaxan ás áreas linfoides onde se encontran as células T poden activar as células T, as cales despois diferéncianse en células T citotóxicas ou axudantes.[84]
Mastocitos
[editar | editar a fonte]- Artigo principal: Mastocito.
Os mastocitos posúen receptores de tipo Toll e interaccionan coas células dendríticas, células B e T para axudar na mediación das funcións inmunitarias adaptativas.[89] Os mastocitos expresan moléculas do MHC de clase II e poden participar na presentación de antíxenos; porén, o papel dos mastocitos na presentación de antíxenos non se comprende moi ben.[90] Os mastocitos poden fagocitar e matar bacterias gramnegativas (por exemplo, salmonela), e procesar os seus antíxenos.[91] Especialízanse en procesar as proteínas das fimbrias que se encontan na superficie da bacteria, as cales están implicadas na adhesión aos tecidos.[92][93] Ademais destas funcións, os mastocitos producen citocinas que inducen unha resposta inflamatoria.[94] Isto é unha parte vital da destrución dos microbios porque as citocinas atraen máis fagocitos cara ao sitio de infección.[91][95]
| Localización principal | Variedade de fenotipos |
|---|---|
| Sangue | neutrófilos, monocitos |
| Medula ósea | macrófagos, monocitos, células sinusoidais, células que tapizan cavidades internas |
| Tecido óseo | osteoclastos |
| Placas de Peyer do tracto gastrointestinal | macrófagos |
| Tecido conectivo | histiocitos, macrófagos, monocitos, células dendríticas |
| Fígado | células de Kupffer, monocitos |
| Pulmóns | macrófagos autorreplicantes, monocitos, mastocitos, células dendríticas |
| Tecido linfoide | macrófagos libres e fixos e monocitos, células dendríticas |
| Tecido nervioso | células microgliais (CD4+) |
| Bazo | macrófagos libres e fixos, monocitos, células sinusoidais |
| Timo | macrófagos libres ou fixos e monocitos |
| Pel | células de Langerhans residentes, outras células dendríticas, macrófagos convencionais, mastocitos |
Fagocitos non profesionais
[editar | editar a fonte]As células moribundas e os microorganismos estraños son fagocitados por outras células distintas dos "fagocitos profesionais".[97] Estas células inclúen as células epiteliais, células endoteliais, fibroblastos e células mesenquimáticas. Denomínanse fagocitos non profesionais para salientar que nelas, a diferenza dos fagocitos profesionais, a fagocitose non é a súa principal función.[98] Os fibroblastos, por exemplo, que poden fagocitar coláxeno durante o proceso de remodelación de cicatrices, tamén fan algún intento de inxerir partículas estrañas.[99]
Os fagocitos non profesionais están máis limitados que os fagocitos profesionais en canto ao tipo de partículas que poden captar. Isto débese a que carecen de receptores fagocíticos eficientes, en concreto de opsoninas, que son moléculas de anticorpos e do complemento creadas polo sistema inmunitario adheridas aos invasores.[11] Ademais, a maioría dos fagocitos non profesionais non producen moléculas que conteñan oxíxeno reactivo en resposta á fagocitose.[100]
| Localización principal | Variedade de fenotipos |
|---|---|
| Sangue, linfa e ganglios linfáticos | linfocitos |
| Sangue, linfa e ganglios linfáticos | células NK e linfocitos granulares grandes (LGL do inglés large granular lymphocytes) |
| Sangue | eosinófilos e basófilos[101] |
| Pel | células epiteliais |
| Vasos sanguíneos | células endoteliais |
| Tecido conectivo | fibroblastos |
Evasión e resistencia dos patóxenos
[editar | editar a fonte]
Un patóxeno só ten éxito á hora de infectar un organismo se pode superar as súas defensas. As bacterias patóxenas e os protozoos desenvolveron diversos métodos para resistir os ataques dos fagocitos e moitos mesmo poden sobrevivir e replicarse dentro das células fagocíticas.[102][103]
Evitar o contacto
[editar | editar a fonte]Hai varios xeitos en que unha bacteria pode evitar o contacto cos fagocitos. Primeiro, pode crecer en sitios a onde os fagocitos non poden viaxar (por exemplo, a superficie da pel sá, sen feridas abertas). Segundo, as bacterias poden superar a resposta inflamatoria; sen esta resposta á infección os fagocitos non son quen de responder adecuadamente. Terceiro, algunhas especies de bacterias poden inhibir a capacidade dos fagocitos de viaxar ao sitio da infección ao interferiren coa quimiotaxe.[102] Cuarto, algunhas bacterias poden evitar o contacto cos fagocitos ao enganar ao sistema inmune para que "pense" que a bacteria é algo "propio". O Treponema pallidum, a bacteria causante da sífilis, evita a detección polos fagocitos ao cubrir a súa superficie con fibronectina,[104] a cal é unha proteína que produce naturalmente o corpo e ten unha función esencial na curación de feridas.[105]
Evitar a fagocitose
[editar | editar a fonte]As bacterias a miúdo producen cápsulas feitas de proteínas ou carbohidratos que cobren as súas células e interfiren coa fagocitose.[102] Algúns exemplos son a cápsula K5 e o antíxeno O O75 que se encontran na superficie da bacteria Escherichia coli,[106] e as cápsulas de exopolisacárido de Staphylococcus epidermidis.[107] Streptococcus pneumoniae produce varios tipos de cápsula que proporcionan diferentes niveis de protección,[108] e os estreptococos do grupo A producen proteínas como a proteína M e proteínas das fimbrias para bloquear a fagocitose. Algunhas proteínas dificultan a inxestión relacionada con opsoninas; Staphylococcus aureus produce a proteína A para bloquear receptores de anticorpos, o cal fai diminuír a efectividade das opsoninas.[109] As especies enteropatóxenas do xénero Yersinia únense utilizando o factor de virulencia YopH a receptores de fagocitos desde os cales inflúen na capacidade das células para exercer a fagocitose.[110]
Supervivencia dentro do fagocito
[editar | editar a fonte]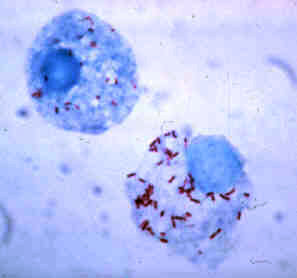
As bacterias desenvolveron estratexias para sobrevivir dentro dos fagocitos, onde continúan evadíndose do sistema inmunitario.[111] Para estaren a salvo dentro do fagocito expresan proteínas chamadas invasinas. Cando está dentro da célula permanecen no citoplasma e evitan as substancias químicas tóxicas contidas nos fagolisosomas.[112] Algunhas bacterias impiden a fusión dun fagosoma e un lisosoma, para formar o fagolisosoma.[102] Outros patóxenos, como Leishmania, crean un vacúolo moi modificado dentro do fagocito, que lle axuda a persistir e replicarse.[113] Algunhas bacterias poden vivir dentro do fagolisosoma. Staphylococcus aureus, por exemplo, produce os encimas catalase e superóxido dismutase, que degradan substancias químicas, como o peróxido de hidróxeno, producidas por fagocitos para matar bacterias.[114] As bacterias poden escapar do fagosoma antes da formación do fagolisosoma: Listeria monocytogenes pode facer un burato na parede do fagosoma usando uns encimas chamados listeriolisina O e fosfolipase C.[115]
Matanza de células
[editar | editar a fonte]As bacterias desenvolveron varios xeitos de matar os fagocitos.[109] Estes inclúen o uso de citolisinas, que forman poros nas membranas celulares do fagocito, estreptolisinas e leucocidinas, que causan a rotura dos gránulos dos neutrófilos, e a liberación de substancias tóxicas,[116][117] e exotoxinas que reducen a dispoñibilidade de ATP do fagocito, necesaria para a fagocitose. Despois de que se inxire unha bacteria, pode matar o fagocito ao liberar toxinas que viaxan a través da membrana do fagosoma ou fagolisosoma e teñen como dianas outras partes da célula.[102]
Alteración da sinalización celular
[editar | editar a fonte]Algunhas estratexias de supervivencia a miúdo implican a alteración das citocinas e outros métodos de sinalización celular para impedir a resposta dos fagocitos á invasión.[118] Os protozoos parasitos Toxoplasma gondii, Trypanosoma cruzi e Leishmania infectan macrófagos e cada un ten un método particular para amansalos.[118] Algunhas especies de Leishmania alteran a sinalización dos macrófagos infectados, reprimen a produción de citocinas e moléculas microbianas (óxido nítrico e especies reactivas do oxíxeno) e comprometen a presentación de antíxenos.[119]
Dano aos hóspedes feito polos fagocitos
[editar | editar a fonte]Os macrfófagos e neutrófilos xogan un papel central no proceso inflamatorio ao liberar proteínas e pequenas moléculas mediadoras da inflamación que controlan a infección, pero que poden tamén danar os tecidos do hóspede. En xeral, os fagocitos pretenden destruír os patóxenos englobándoos dentro da célula e someténdoos a unha batería de substancias químicas tóxicas dentro dun fagolisosoma. Se un fagocito non consegue fagocitar a súa diana, estes axentes tóxicos poden ser liberados no ambiente que rodea a célula (o que se denomina "fagocitose frustrada"). Como estes axentes son tamén tóxicos para a célula hóspede, poden causar un amplo dano a células e tecidos sans.[120]
Cando os neutrófilos liberan o contido dos seus gránulos nos riles, dito contido (compostos reactivos do oxíxeno e proteases) degradan a matriz extracelular das células hóspede e poden danar as células glomerulares, afectando a súa capacidade de filtrar o sangue e causando cambios de forma. Ademais, os produtos da fosfolipase (por exemplo, leucotrienos) intensifican os danos. Esta liberación de substancias promove a quimiotaxe de máis neutrófilos ao sitio de infección, e as células glomerulares poden sufrir máis danos polas moléculas de adhesión durante a migración dos neutrófilos. Os danos causados ás células glomerulares poden orixinar unha insuficiencia renal.[121]
Os neutrófilos tamén xogan un papel clave no desenvolvemento da maioría das formas de lesión pulmonar aguda.[122] Neste caso, os neutrófilos activados liberan os contidos dos seus gránulos tóxicos no ambiente pulmonar.[123] Realizáronse experimentos que mostran que unha redución no número de neutrófilos diminúe os efectos das lesións pulmonares agudas,[124] pero o tratamento de inhibición dos neutrófilos non é realista clinicamente, xa que deixa o hóspede en estado vulnerable ante a infección.[123] No fígado, dos danos causados por neutrófilos poden contribuír á disfunción e a lesións en resposta á liberación de endotoxinas producida por bacterias, sepse, trauma, hepatite alcohólica, isquemia e choque hipovolémico resultado dunha hemorraxia aguda.[125]
As substancias químicas liberadas polos macrófagos poden tamén danar os tecidos do hóspede. O TNF-α é un importante composto químico que liberan os macrófagos que causan que o sangue dos vasos sanguíneos pequenos coagule para impedir o espallamento dunha infección.[126] Porén, se unha infección bacteriana se espalla polo sangue, o TNF-α libérase en órganos vitais, o cal causa vasodilatación e un descenso no volume de plasma sanguíneo; isto pode ir seguido dun choque séptico. Durante o choque séptico, a liberación de TNF-α causa un bloqueo dos pequenos vasos sanguíneos que regan os órganos vitais e os órganos poden fallar. O choque séptico pode orixinar a morte.[13]
Orixes evolutivas
[editar | editar a fonte]A fagocitose é algo común na natureza e probablemente apareceu moi cedo na evolución,[127] evolucionando primeiro en eucariotas unicelulares.[128] As amebas son protistas unicellares que se separaron da póla evolutiva que levaba aos metazoos pouco despois da diverxencia das plantas, e comparten moitas funcións específicas coas células fagocíticas de mamíferos.[128] Dictyostelium discoideum, por exemplo, é unha ameba que vive no solo e aliméntase de bacterias. Igual que os fagocitos animais, fagocita as bacterias principalmente por medio de receptores de tipo Toll e ten outras funcións biolóxicas en común cos macrófagos.[129] Dictyostelium discoideum é un morganismo social; agrégase cando ten escaseza de alimentos e forma un pseudoplasmodio migrador. Este organismo multicelular produce finalmente un corpo frutífero con esporas que son resistentes aos perigos ambientais. Antes da formación dos corpos frutíferos, as células migran como un organismo parecido a unha masa reptante durante varios días. Durante este período a exposición a toxinas ou patóxenos bacterianos podería poñer en perigo a supervivencia da especie ao limitar a produción de esporas. Non obstante, algunhas das amebas fagocitan as bacterias e absorben as toxinas que circulan dentro da masa reptante e estas amebas finalmente morren. Son xeneticamente idénticas ás outras amebas da masa reptante; o seu autosacrificio para protexer as outras amebas das bacterias é similar ao autosacrificio dos fagocitos visto no sistema inmunitario dos vertebrados superiores. Esta función inmunitaria inmune nas amebas sociais indica que un mecanismo de busca de alimento celular conservado evolutivamente se puido ter adaptado a facer funcións de defensa moito antes da diversificación das amebas en formas vivas superiores.[130] Os fagocitos aparecen por todo o reino animal,[3] desde as esponxas mariñas aos insectos e vertebrados inferiores e superiores.[131][132] A capacidade das amebas de distinguir entre o propio e o non propio ten unha importancia esencial e é a raíz do sistema inmunitario de moitas especies de amebas.[8]
Notas
[editar | editar a fonte]- ↑ 1,0 1,1 Little, C., Fowler H.W., Coulson J. (1983). The Shorter Oxford English Dictionary. Oxford University Press (Guild Publishing). pp. 1566–67.
- ↑ 2,00 2,01 2,02 2,03 2,04 2,05 2,06 2,07 2,08 2,09 Delves et al. 2006, pp. 2–10
- ↑ 3,0 3,1 Delves et al. 2006, p. 250
- ↑ Delves et al. 2006, p. 251
- ↑ 5,0 5,1 5,2 5,3 Hoffbrand, Pettit & Moss 2005, p. 331
- ↑ Ilya Mechnikov, consultado o 28 de novembro de 2008. From Nobel Lectures, Physiology or Medicine 1901–1921, Elsevier Publishing Company, Amsterdam, 1967. Arquivado 22 de agosto de 2008 en Wayback Machine.
- ↑ 7,0 7,1 Schmalstieg, FC; AS Goldman (2008). "Ilya Ilich Metchnikoff (1845–1915) and Paul Ehrlich (1854–1915): the centennial of the 1908 Nobel Prize in Physiology or Medicine". Journal of Medical Biography 16 (2): 96–103. PMID 18463079. doi:10.1258/jmb.2008.008006.
- ↑ 8,0 8,1 Janeway, Chapter: Evolution of the innate immune system. consultado o 20 de marzo de 2009
- ↑ 9,0 9,1 Ernst & Stendahl 2006, p. 186
- ↑ 10,0 10,1 Robinson & Babcock 1998, p. 187 and Ernst & Stendahl 2006, pp. 7–10
- ↑ 11,0 11,1 Ernst & Stendahl 2006, p. 10
- ↑ 12,0 12,1 Thompson, CB (1995). "Apoptosis in the pathogenesis and treatment of disease". Science 267 (5203): 1456–62. Bibcode:1995Sci...267.1456T. PMID 7878464. doi:10.1126/science.7878464.
- ↑ 13,0 13,1 13,2 Janeway, Chapter: Induced innate responses to infection.
- ↑ 14,0 14,1 Fang FC (outubro de 2004). "Antimicrobial reactive oxygen and nitrogen species: concepts and controversies". Nat. Rev. Microbiol. 2 (10): 820–32. PMID 15378046. doi:10.1038/nrmicro1004.
- ↑ 15,0 15,1 Delves et al. 2006, pp. 172–84
- ↑ 16,0 16,1 16,2 Kaufmann SH (2019). "Immunology's Coming of Age". Frontiers in Immunology 10: 684. PMC 6456699. PMID 31001278. doi:10.3389/fimmu.2019.00684.
- ↑ Aterman K (1 de abril de 1998). "Medals, memoirs—and Metchnikoff". J. Leukoc. Biol. 63 (4): 515–17. PMID 9544583. doi:10.1002/jlb.63.4.515. Arquivado dende o orixinal o 09 de decembro de 2012. Consultado o 24 de decembro de 2020.
- ↑ "Ilya Mechnikov". The Nobel Foundation. Consultado o 19 de decembro de 2014.
- ↑ Delves et al. 2006, p. 263
- ↑ Robinson & Babcock 1998, p. vii
- ↑ Ernst & Stendahl 2006, p. 4
- ↑ Ernst & Stendahl 2006, p. 78
- ↑ 23,0 23,1 Hampton MB, Vissers MC, Winterbourn CC; Vissers; Winterbourn (febreiro de 1994). "A single assay for measuring the rates of phagocytosis and bacterial killing by neutrophils". J. Leukoc. Biol. 55 (2): 147–52. PMID 8301210. doi:10.1002/jlb.55.2.147. Arquivado dende o orixinal o 28 de decembro de 2012. Consultado o 24 de decembro de 2020.
- ↑ Delves et al. 2006, pp. 6–7
- ↑ Sompayrac 2008, p. 3
- ↑ Sompayrac 2008, p. 2
- ↑ Sompayrac 2008, pp. 13–16
- ↑ Dale DC, Boxer L, Liles WC; Boxer; Liles (agosto de 2008). "The phagocytes: neutrophils and monocytes". Blood 112 (4): 935–45. PMID 18684880. doi:10.1182/blood-2007-12-077917.
- ↑ Dahlgren, C; A Karlsson (17 de decembro de 1999). "Respiratory burst in human neutrophils". Journal of Immunological Methods 232 (1–2): 3–14. PMID 10618505. doi:10.1016/S0022-1759(99)00146-5.
- ↑ Shatwell, KP; AW Segal (1996). "NADPH oxidase". The International Journal of Biochemistry & Cell Biology 28 (11): 1191–95. PMID 9022278. doi:10.1016/S1357-2725(96)00084-2.
- ↑ Klebanoff SJ (1999). "Myeloperoxidase". Proc. Assoc. Am. Physicians 111 (5): 383–89. PMID 10519157. doi:10.1111/paa.1999.111.5.383.
- ↑ Meyer KC (setembro de 2004). "Neutrophils, myeloperoxidase, and bronchiectasis in cystic fibrosis: green is not good". J. Lab. Clin. Med. 144 (3): 124–26. PMID 15478278. doi:10.1016/j.lab.2004.05.014.
- ↑ Hoffbrand, Pettit & Moss 2005, p. 118
- ↑ Delves et al. 2006, pp. 6–10
- ↑ Schroder K, Hertzog PJ, Ravasi T, Hume DA; Hertzog; Ravasi; Hume (febreiro de 2004). "Interferon-gamma: an overview of signals, mechanisms and functions". J. Leukoc. Biol. 75 (2): 163–89. PMID 14525967. doi:10.1189/jlb.0603252. Arquivado dende o orixinal o 03 de xullo de 2010. Consultado o 24 de decembro de 2020.
- ↑ Delves et al. 2006, p. 188
- ↑ 37,0 37,1 Sompayrac 2008, p. 17
- ↑ Lipu HN, Ahmed TA, Ali S, Ahmed D, Waqar MA; Ahmed; Ali; Ahmed; Waqar (setembro de 2008). "Chronic granulomatous disease". J Pak Med Assoc 58 (9): 516–18. PMID 18846805.
- ↑ Kaplan J, De Domenico I, Ward DM; De Domenico; Ward (xaneiro de 2008). "Chediak-Higashi syndrome". Curr. Opin. Hematol. 15 (1): 22–29. PMID 18043242. doi:10.1097/MOH.0b013e3282f2bcce.
- ↑ Sompayrac 2008, p. 7
- ↑ de Almeida SM, Nogueira MB, Raboni SM, Vidal LR; Nogueira; Raboni; Vidal (outubro de 2007). "Laboratorial diagnosis of lymphocytic meningitis". Braz J Infect Dis 11 (5): 489–95. PMID 17962876. doi:10.1590/s1413-86702007000500010.
- ↑ Sompayrac 2008, p. 22
- ↑ Sompayrac 2008, p. 63
- ↑ "Apoptosis". Merriam-Webster Online Dictionary. Consultado o 19 de decembro de 2014.
- ↑ Li MO, Sarkisian MR, Mehal WZ, Rakic P, Flavell RA; Sarkisian; Mehal; Rakic; Flavell (novembro de 2003). "Phosphatidylserine receptor is required for clearance of apoptotic cells". Science 302 (5650): 1560–63. PMID 14645847. doi:10.1126/science.1087621.
- ↑ Nagata S, Sakuragi T, Segawa K (decembro de 2019). "Flippase and scramblase for phosphatidylserine exposure". Current Opinion in Immunology 62: 31–38. PMID 31837595. doi:10.1016/j.coi.2019.11.009.
- ↑ Wang X (2003). "Cell corpse engulfment mediated by C. elegans phosphatidylserine receptor through CED-5 and CED-12". Science 302 (5650): 1563–1566. Bibcode:2003Sci...302.1563W. PMID 14645848. doi:10.1126/science.1087641. (Free registration required for online access)
- ↑ Savill J, Gregory C, Haslett C (2003). "Eat me or die". Science 302 (5650): 1516–17. PMID 14645835. doi:10.1126/science.1092533. hdl:1842/448.
- ↑ Zhou Z, Yu X; Yu (outubro de 2008). "Phagosome maturation during the removal of apoptotic cells: receptors lead the way". Trends Cell Biol. 18 (10): 474–85. PMC 3125982. PMID 18774293. doi:10.1016/j.tcb.2008.08.002.
- ↑ Sompayrac 2008, p. 44
- ↑ Sompayrac 2008, p. 4
- ↑ Sompayrac 2008, pp. 24–35
- ↑ Delves et al. 2006, pp. 171–184
- ↑ Delves et al. 2006, pp. 456
- ↑ Timothy Lee (2004). "Antigen Presenting Cells (APC)". Immunology for 1st Year Medical Students. Dalhousie University. Arquivado dende o orixinal o 12 de xaneiro de 2008. Consultado o 19 de decembro de 2014.
- ↑ Delves et al. 2006, p. 161
- ↑ Sompayrac 2008, p. 8
- ↑ Delves et al. 2006, pp. 237–242
- ↑ Lange C, Dürr M, Doster H, Melms A, Bischof F; Dürr; Doster; Melms; Bischof (2007). "Dendritic cell-regulatory T-cell interactions control self-directed immunity". Immunol. Cell Biol. 85 (8): 575–81. PMID 17592494. doi:10.1038/sj.icb.7100088.
- ↑ Steinman, Ralph M. (2004). "Dendritic Cells and Immune Tolerance". The Rockefeller University. Arquivado dende o orixinal o 11 de marzo de 2009. Consultado o 19 de decembro de 2014.
- ↑ Romagnani, S (2006). "Immunological tolerance and autoimmunity". Internal and Emergency Medicine. 1 (3): 187–96. PMID 17120464. doi:10.1007/BF02934736.
- ↑ Sompayrac 2008, pp. 16–17
- ↑ Sompayrac 2008, pp. 18–19
- ↑ Delves et al. 2006, p. 6
- ↑ Zen K, Parkos CA; Parkos (outubro de 2003). "Leukocyte-epithelial interactions". Curr. Opin. Cell Biol. 15 (5): 557–64. PMID 14519390. doi:10.1016/S0955-0674(03)00103-0.
- ↑ Sompayrac 2008, p. 79
- ↑ Hoffbrand, Pettit & Moss 2005, p. 117
- ↑ Delves et al. 2006, pp. 1–6
- ↑ Sompayrac 2008, p. 45
- ↑ Takahashi K, Naito M, Takeya M; Naito; Takeya (xullo de 1996). "Development and heterogeneity of macrophages and their related cells through their differentiation pathways". Pathol. Int. 46 (7): 473–85. PMID 8870002. doi:10.1111/j.1440-1827.1996.tb03641.x.
- ↑ Krombach F, Münzing S, Allmeling AM, Gerlach JT, Behr J, Dörger M; Münzing; Allmeling; Gerlach; Behr; Dörger (setembro de 1997). "Cell size of alveolar macrophages: an interspecies comparison". Environ. Health Perspect. 105 Suppl 5 (Suppl 5): 1261–63. JSTOR 3433544. PMC 1470168. PMID 9400735. doi:10.2307/3433544.
- ↑ 72,0 72,1 72,2 72,3 72,4 Delves et al. 2006, pp. 31–36
- ↑ Ernst & Stendahl 2006, p. 8
- ↑ Delves et al. 2006, p. 156
- ↑ Delves et al. 2006, p. 187
- ↑ Stvrtinová, Viera; Ján Jakubovský and Ivan Hulín (1995). "Neutrophils, central cells in acute inflammation". Inflammation and Fever from Pathophysiology: Principles of Disease. Computing Centre, Slovak Academy of Sciences: Academic Electronic Press. ISBN 978-80-967366-1-4. Arquivado dende o orixinal o 31 de decembro de 2010. Consultado o 19 de decembro de 2014.
- ↑ Delves et al. 2006, p. 4
- ↑ 78,0 78,1 Sompayrac 2008, p. 18
- ↑ Linderkamp O, Ruef P, Brenner B, Gulbins E, Lang F; Ruef; Brenner; Gulbins; Lang (decembro de 1998). "Passive deformability of mature, immature, and active neutrophils in healthy and septicemic neonates". Pediatr. Res. 44 (6): 946–50. PMID 9853933. doi:10.1203/00006450-199812000-00021.
- ↑ Paoletti, Notario & Ricevuti 1997, p. 62
- ↑ Soehnlein O, Kenne E, Rotzius P, Eriksson EE, Lindbom L; Kenne; Rotzius; Eriksson; Lindbom (xaneiro de 2008). "Neutrophil secretion products regulate anti-bacterial activity in monocytes and macrophages". Clin. Exp. Immunol. 151 (1): 139–45. PMC 2276935. PMID 17991288. doi:10.1111/j.1365-2249.2007.03532.x.
- ↑ Soehnlein O, Kai-Larsen Y, Frithiof R (outubro de 2008). "Neutrophil primary granule proteins HBP and HNP1-3 boost bacterial phagocytosis by human and murine macrophages". J. Clin. Invest. 118 (10): 3491–502. PMC 2532980. PMID 18787642. doi:10.1172/JCI35740.
- ↑ Steinman RM, Cohn ZA; Cohn (1973). "Identification of a novel cell type in peripheral lymphoid organs of mice. I. Morphology, quantitation, tissue distribution". J. Exp. Med. 137 (5): 1142–62. PMC 2139237. PMID 4573839. doi:10.1084/jem.137.5.1142.
- ↑ 84,0 84,1 Steinman, Ralph. "Dendritic Cells". The Rockefeller University. Arquivado dende o orixinal o 27 de xuño de 2009. Consultado o 19 de decembro de 2014.
- ↑ Guermonprez P, Valladeau J, Zitvogel L, Théry C, Amigorena S; Valladeau; Zitvogel; Théry; Amigorena (2002). "Antigen presentation and T cell stimulation by dendritic cells". Annu. Rev. Immunol. 20: 621–67. PMID 11861614. doi:10.1146/annurev.immunol.20.100301.064828.
- ↑ Hoffbrand, Pettit & Moss 2005, p. 134
- ↑ Sallusto F, Lanzavecchia A; Lanzavecchia (2002). "The instructive role of dendritic cells on T-cell responses". Arthritis Res. 4 Suppl 3 (Suppl 3): S127–32. PMC 3240143. PMID 12110131. doi:10.1186/ar567.
- ↑ Sompayrac 2008, pp. 42–46
- ↑ Novak N, Bieber T, Peng WM; Bieber; Peng (2010). "The immunoglobulin E-Toll-like receptor network". International Archives of Allergy and Immunology 151 (1): 1–7. PMID 19672091. doi:10.1159/000232565. Consultado o 19 de decembro de 2014.
- ↑ Kalesnikoff J, Galli SJ; Galli (novembro de 2008). "New developments in mast cell biology". Nature Immunology 9 (11): 1215–23. PMC 2856637. PMID 18936782. doi:10.1038/ni.f.216.
- ↑ 91,0 91,1 Malaviya R, Abraham SN; Abraham (febreiro de 2001). "Mast cell modulation of immune responses to bacteria". Immunol. Rev. 179: 16–24. PMID 11292019. doi:10.1034/j.1600-065X.2001.790102.x.
- ↑ Connell I, Agace W, Klemm P, Schembri M, Mărild S, Svanborg C; Agace; Klemm; Schembri; Mărild; Svanborg (setembro de 1996). "Type 1 fimbrial expression enhances Escherichia coli virulence for the urinary tract". Proc. Natl. Acad. Sci. U.S.A. 93 (18): 9827–32. Bibcode:1996PNAS...93.9827C. PMC 38514. PMID 8790416. doi:10.1073/pnas.93.18.9827.
- ↑ Malaviya R, Twesten NJ, Ross EA, Abraham SN, Pfeifer JD; Twesten; Ross; Abraham; Pfeifer (febreiro de 1996). "Mast cells process bacterial Ags through a phagocytic route for class I MHC presentation to T cells". J. Immunol. 156 (4): 1490–96. PMID 8568252. Consultado o 19 de decembro de 2014.
- ↑ Taylor ML, Metcalfe DD; Metcalfe (2001). "Mast cells in allergy and host defense". Allergy Asthma Proc 22 (3): 115–19. PMID 11424870. doi:10.2500/108854101778148764.
- ↑ Urb M, Sheppard DC (2012). "The role of mast cells in the defence against pathogens". PLOS Pathogens 8 (4): e1002619. PMC 3343118. PMID 22577358. doi:10.1371/journal.ppat.1002619.
- ↑ 96,0 96,1 Paoletti, Notario & Ricevuti 1997, p. 427
- ↑ Birge RB, Ucker DS; Ucker (xullo de 2008). "Innate apoptotic immunity: the calming touch of death". Cell Death Differ. 15 (7): 1096–1102. PMID 18451871. doi:10.1038/cdd.2008.58.
- ↑ Couzinet S, Cejas E, Schittny J, Deplazes P, Weber R, Zimmerli S; Cejas; Schittny; Deplazes; Weber; Zimmerli (decembro de 2000). "Phagocytic uptake of Encephalitozoon cuniculi by nonprofessional phagocytes". Infect. Immun. 68 (12): 6939–45. PMC 97802. PMID 11083817. doi:10.1128/IAI.68.12.6939-6945.2000.
- ↑ Segal G, Lee W, Arora PD, McKee M, Downey G, McCulloch CA; Lee; Arora; McKee; Downey; McCulloch (xaneiro de 2001). "Involvement of actin filaments and integrins in the binding step in collagen phagocytosis by human fibroblasts". Journal of Cell Science 114 (Pt 1): 119–129. PMID 11112696.
- ↑ Rabinovitch M (marzo de 1995). "Professional and non-professional phagocytes: an introduction". Trends Cell Biol. 5 (3): 85–87. PMID 14732160. doi:10.1016/S0962-8924(00)88955-2.
- ↑ Lin A, Loré K (2017). "Granulocytes: New Members of the Antigen-Presenting Cell Family". Frontiers in Immunology 8: 1781. PMC 5732227. PMID 29321780. doi:10.3389/fimmu.2017.01781.
- ↑ 102,0 102,1 102,2 102,3 102,4 Todar, Kenneth. "Mechanisms of Bacterial Pathogenicity: Bacterial Defense Against Phagocytes". 2008. Consultado o 19 de decembro de 2014.
- ↑ Alexander J, Satoskar AR, Russell DG; Satoskar; Russell (setembro de 1999). "Leishmania species: models of intracellular parasitism" (PDF). J. Cell Sci. 112 (18): 2993–3002. PMID 10462516. Arquivado dende o orixinal (PDF) o 02 de xuño de 2021. Consultado o 1 de xuño de 2021.
- ↑ Celli J, Finlay BB; Finlay (maio de 2002). "Bacterial avoidance of phagocytosis". Trends Microbiol. 10 (5): 232–37. PMID 11973157. doi:10.1016/S0966-842X(02)02343-0.
- ↑ Valenick LV, Hsia HC, Schwarzbauer JE; Hsia; Schwarzbauer (setembro de 2005). "Fibronectin fragmentation promotes alpha4beta1 integrin-mediated contraction of a fibrin-fibronectin provisional matrix". Experimental Cell Research 309 (1): 48–55. PMID 15992798. doi:10.1016/j.yexcr.2005.05.024.
- ↑ Burns SM, Hull SI; Hull (agosto de 1999). "Loss of resistance to ingestion and phagocytic killing by O(-) and K(-) mutants of a uropathogenic Escherichia coli O75:K5 strain". Infect. Immun. 67 (8): 3757–62. PMC 96650. PMID 10417134. doi:10.1128/IAI.67.8.3757-3762.1999.
- ↑ Vuong C, Kocianova S, Voyich JM (decembro de 2004). "A crucial role for exopolysaccharide modification in bacterial biofilm formation, immune evasion, and virulence". J. Biol. Chem. 279 (52): 54881–86. PMID 15501828. doi:10.1074/jbc.M411374200.
- ↑ Melin M, Jarva H, Siira L, Meri S, Käyhty H, Väkeväinen M; Jarva; Siira; Meri; Käyhty; Väkeväinen (febreiro de 2009). "Streptococcus pneumoniae capsular serotype 19F is more resistant to C3 deposition and less sensitive to opsonophagocytosis than serotype 6B". Infect. Immun. 77 (2): 676–84. PMC 2632042. PMID 19047408. doi:10.1128/IAI.01186-08.
- ↑ 109,0 109,1 Foster TJ (decembro de 2005). "Immune evasion by staphylococci". Nat. Rev. Microbiol. 3 (12): 948–58. PMID 16322743. doi:10.1038/nrmicro1289.
- ↑ Fällman M, Deleuil F, McGee K (febreiro de 2002). "Resistance to phagocytosis by Yersinia". International Journal of Medical Microbiology : IJMM 291 (6–7): 501–9. PMID 11890550. doi:10.1078/1438-4221-00159.
- ↑ Sansonetti P (decembro de 2001). "Phagocytosis of bacterial pathogens: implications in the host response". Semin. Immunol. 13 (6): 381–90. PMID 11708894. doi:10.1006/smim.2001.0335.
- ↑ Dersch P, Isberg RR; Isberg (marzo de 1999). "A region of the Yersinia pseudotuberculosis invasin protein enhances integrin-mediated uptake into mammalian cells and promotes self-association". EMBO J. 18 (5): 1199–1213. PMC 1171211. PMID 10064587. doi:10.1093/emboj/18.5.1199.
- ↑ Antoine JC, Prina E, Lang T, Courret N; Prina; Lang; Courret (outubro de 1998). "The biogenesis and properties of the parasitophorous vacuoles that harbour Leishmania in murine macrophages". Trends Microbiol. 6 (10): 392–401. PMID 9807783. doi:10.1016/S0966-842X(98)01324-9.
- ↑ Das D, Saha SS, Bishayi B; Saha; Bishayi (xullo de 2008). "Intracellular survival of Staphylococcus aureus: correlating production of catalase and superoxide dismutase with levels of inflammatory cytokines". Inflamm. Res. 57 (7): 340–49. PMID 18607538. doi:10.1007/s00011-007-7206-z.
- ↑ Hara H, Kawamura I, Nomura T, Tominaga T, Tsuchiya K, Mitsuyama M; Kawamura; Nomura; Tominaga; Tsuchiya; Mitsuyama (agosto de 2007). "Cytolysin-dependent escape of the bacterium from the phagosome is required but not sufficient for induction of the Th1 immune response against Listeria monocytogenes infection: distinct role of Listeriolysin O determined by cytolysin gene replacement". Infect. Immun. 75 (8): 3791–3801. PMC 1951982. PMID 17517863. doi:10.1128/IAI.01779-06.
- ↑ Datta V, Myskowski SM, Kwinn LA, Chiem DN, Varki N, Kansal RG, Kotb M, Nizet V; Myskowski; Kwinn; Chiem; Varki; Kansal; Kotb; Nizet (maio de 2005). "Mutational analysis of the group A streptococcal operon encoding streptolysin S and its virulence role in invasive infection". Mol. Microbiol. 56 (3): 681–95. PMID 15819624. doi:10.1111/j.1365-2958.2005.04583.x.
- ↑ Iwatsuki K, Yamasaki O, Morizane S, Oono T; Yamasaki; Morizane; Oono (xuño de 2006). "Staphylococcal cutaneous infections: invasion, evasion and aggression". J. Dermatol. Sci. 42 (3): 203–14. PMID 16679003. doi:10.1016/j.jdermsci.2006.03.011.
- ↑ 118,0 118,1 Denkers EY, Butcher BA; Butcher (xaneiro de 2005). "Sabotage and exploitation in macrophages parasitized by intracellular protozoans". Trends Parasitol. 21 (1): 35–41. PMID 15639739. doi:10.1016/j.pt.2004.10.004.
- ↑ Gregory DJ, Olivier M; Olivier (2005). "Subversion of host cell signalling by the protozoan parasite Leishmania". Parasitology. 130 Suppl: S27–35. PMID 16281989. doi:10.1017/S0031182005008139.
- ↑ Paoletti pp. 426–30
- ↑ Heinzelmann M, Mercer-Jones MA, Passmore JC; Mercer-Jones; Passmore (agosto de 1999). "Neutrophils and renal failure". Am. J. Kidney Dis. 34 (2): 384–99. PMID 10430993. doi:10.1016/S0272-6386(99)70375-6.
- ↑ Lee WL, Downey GP; Downey (febreiro de 2001). "Neutrophil activation and acute lung injury". Curr Opin Crit Care 7 (1): 1–7. PMID 11373504. doi:10.1097/00075198-200102000-00001.
- ↑ 123,0 123,1 Moraes TJ, Zurawska JH, Downey GP; Zurawska; Downey (xaneiro de 2006). "Neutrophil granule contents in the pathogenesis of lung injury". Curr. Opin. Hematol. 13 (1): 21–27. PMID 16319683. doi:10.1097/01.moh.0000190113.31027.d5.
- ↑ Abraham E (abril de 2003). "Neutrophils and acute lung injury". Crit. Care Med. 31 (4 Suppl): S195–99. PMID 12682440. doi:10.1097/01.CCM.0000057843.47705.E8.
- ↑ Ricevuti G (decembro de 1997). "Host tissue damage by phagocytes". Ann. N. Y. Acad. Sci. 832 (1): 426–48. Bibcode:1997NYASA.832..426R. PMID 9704069. doi:10.1111/j.1749-6632.1997.tb46269.x.
- ↑ Charley B, Riffault S, Van Reeth K; Riffault; Van Reeth (outubro de 2006). "Porcine innate and adaptative immune responses to influenza and coronavirus infections". Ann. N. Y. Acad. Sci. 1081 (1): 130–36. Bibcode:2006NYASA1081..130C. PMC 7168046. PMID 17135502. doi:10.1196/annals.1373.014. hdl:1854/LU-369324.
- ↑ Sompayrac 2008, p. 1
- ↑ 128,0 128,1 Cosson P, Soldati T; Soldati (xuño de 2008). "Eat, kill or die: when amoeba meets bacteria". Curr. Opin. Microbiol. 11 (3): 271–76. PMID 18550419. doi:10.1016/j.mib.2008.05.005.
- ↑ Bozzaro S, Bucci C, Steinert M; Bucci; Steinert (2008). Phagocytosis and host-pathogen interactions in Dictyostelium with a look at macrophages. Int Rev Cell Mol Biol. International Review of Cell and Molecular Biology 271. pp. 253–300. ISBN 9780123747280. PMID 19081545. doi:10.1016/S1937-6448(08)01206-9.
- ↑ Chen G, Zhuchenko O, Kuspa A; Zhuchenko; Kuspa (agosto de 2007). "Immune-like phagocyte activity in the social amoeba". Science 317 (5838): 678–81. Bibcode:2007Sci...317..678C. PMC 3291017. PMID 17673666. doi:10.1126/science.1143991.
- ↑ Delves et al. 2006, pp. 251–252
- ↑ Hanington PC, Tam J, Katzenback BA, Hitchen SJ, Barreda DR, Belosevic M; Tam; Katzenback; Hitchen; Barreda; Belosevic (abril de 2009). "Development of macrophages of cyprinid fish". Dev. Comp. Immunol. 33 (4): 411–29. PMID 19063916. doi:10.1016/j.dci.2008.11.004.
Véxase tamén
[editar | editar a fonte]Bibliografía
[editar | editar a fonte]- Delves, P. J.; Martin, S. J.; Burton, D. R.; Roit, I. M. (2006). Roitt's Essential Immunology (11th ed.). Malden, MA: Blackwell Publishing. ISBN 978-1-4051-3603-7.
- Ernst, J. D.; Stendahl, O., eds. (2006). Phagocytosis of Bacteria and Bacterial Pathogenicity. New York: Cambridge University Press. ISBN 978-0-521-84569-4. Website
- Hoffbrand, A. V.; Pettit, J. E.; Moss, P. A. H. (2005). Essential Haematology (4th ed.). Londres: Blackwell Science. ISBN 978-0-632-05153-3.
- Paoletti, R.; Notario, A.; Ricevuti, G., eds. (1997). Phagocytes: Biology, Physiology, Pathology, and Pharmacotherapeutics. New York: The New York Academy of Sciences. ISBN 978-1-57331-102-1.
- Robinson, J. P.; Babcock, G. F., eds. (1998). Phagocyte Function — A guide for research and clinical evaluation. New York: Wiley–Liss. ISBN 978-0-471-12364-4.
- Sompayrac, L. (2008). How the Immune System Works (3rd ed.). Malden, MA: Blackwell Publishing. ISBN 978-1-4051-6221-0.
Ligazóns externas
[editar | editar a fonte]| Wikimedia Commons ten máis contidos multimedia na categoría: Fagocito |
- Phagocytes Medical Subject Headings (MeSH) na Biblioteca Nacional de Medicina dos EUA.
- Vídeo dun glóbulo branco perseguindo e fagocitando unha bacteria