Mitokondrio
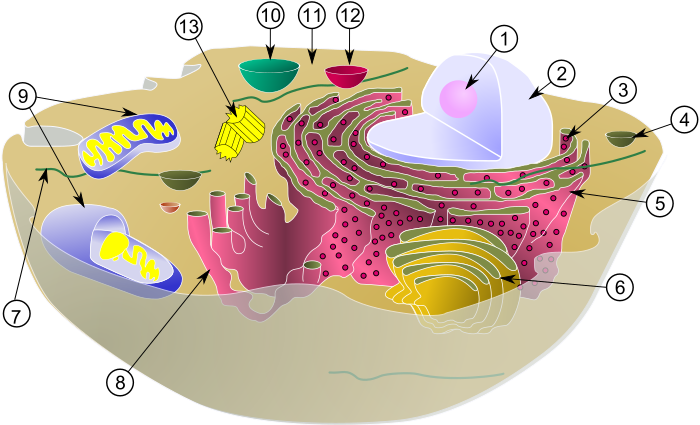
(1) nukleoloa
(2) nukleoa
(3) erribosoma (puntu txikiak)
(4) besikula
(5) erretikulu endoplasmatiko pikortsua
(6) Golgiren aparatua
(7) zitoeskeletoa
(8) erretikulu endoplasmatiko leuna
(9) mitokondrioak
(10) bakuoloa
(11) zitoplasma
(12) lisosoma
(13) zentrioloak zentrosoman

Mitokondrioak zelula eukariotoetan dauden organuluak dira, biribilduak ala baba itxurakoak, zelularen energia-beharrak asetzen dituztenak. Izan ere, mitokondrioetan ATP sintetizatzen da, energia-eramailea den molekularik garrantzitsuena[1]. Prokariotoek ez dute mitokondriorik.
1-7 μ luze dira, eta 0,3-1 μ zabal. Mikroskopio optikoarekin ez dira ondo ikusten, eta beren egitura zehatza aztertzeko mikroskopio elektronikoa erabiltzen da. Haien kopurua oso aldakorra da zelularen arabera, globulu gorriek ez dute mitokondriorik baina gibeleko zelulek 2000 ere izan ditzakete[2]. Animalia-zelula gehienek 500 inguru dituzte. Mitokondrio multzoari kondrioma deritzo.
Etimologia
[aldatu | aldatu iturburu kodea]Mitokondrio izena grezierazko μίτος (mitos, haria) eta χονδρίον (kondrion, alea) hitzen elkarketatik dator eta "harizko alea" esan nahi du[3].
Aurkikuntza eta ikerketaren historia
[aldatu | aldatu iturburu kodea]Mitokondrioa 1840. hamarkadan aurkitu zela uste da[4] baina Richard Altmann patologo alemaniarrak 1890 urtean deskribatu zuen aurrenekoz[5]. Bioblasto izena eman zion eta 1898an Carl Benda izan zen Mitokondrio izena asmatu zuena[6]. 1904. urtean Leonor Michaelis alemaniarrak landare zeluletan aurkitu zuen[7] eta 1908an Claudius Regaudekin batera proteina eta lipidoz egina zegoela proposatu zuen.
1912. urtean arnasketa zelularrarekin lotu zen[4] eta 1925ean David Keilin errusiarrak zitokromoa aurkitu eta arnas katea deskribatu zuen[4]. 1940. hamarkadan arnas katea ikertu zen eta 1952an mikrografia elektronikoak agertu zirenean mitokondrioaren egituren analisi askoz sakonagoak egiteko aukera izan zen[4]. Philip Siekevitz estatubatuarrak "zelularen energia etxea" kontzeptua plazaratu zuen 1957. urtean[8].
1967an erribosomak aurkitu ziren mitokondrioaren barnean[9] eta 1976an legamiaren DNA mitokondrialaren mapa osatu zen[4].
Egitura
[aldatu | aldatu iturburu kodea]
1) Barneko mintza
2) Kanpoko mintza
3) Mitokondrio-gandorrak
4) Matrizea
Mitokondrioaren molfologia aldakorra izan daiteke, itxura deformatu, elkarrekin fusionatu edo zatitu egiteko gaitasuna dutelako baina orokorrean elipse luzexkak izaten dira. Egitura ordea ongi mantentzen dute, elkarren artean desberdinak diren bi mintz, bien arteko tartea eta barneko matrizea hain zuzen ere.
Kanpoko mintza
[aldatu | aldatu iturburu kodea]Zelula mintzaren antzeko egitura du, lipido geruza bikoitza eta fosfolipido-protina ratio antzekoa. Ioi, metabolito, nukleotido eta polipeptido askorekiko iragazkorra da. Horretarako proteina bereziak ditu, porina izenekoak. Voltaiaren araberako jarduera dute eta 5.000 daltonetako pisu molekularra duten molekula handiak pasatzen uzten dituzte[10]. Entzima batzuk ere baditu, gantz azidoak luzatu, adrenalina oxidatu edota triptofanoa degradatzen dutenak adibidez.
Mitokondrioen kanpoko mintza erretikulu endoplasmatikoaren mintzarekin lotzen da batzuetan eta MAM konplexua (Mitokondrioari Asozaturiko E.E.-ren Mintza) eratzen du. Organulu bien arteko kaltzio ioien (Ca2+) eta fosfolipidoen transferentzia gertatzen da orduan[11].
Kanpoko mintza hausten bada, mintz arteko eremuko proteinak zitosolean zabalduko dira eta zelularen heriotza eragingo du[12].
Mintz arteko eremua
[aldatu | aldatu iturburu kodea]Mitokondrioaren kanpoko eta barneko mintzen artean dagoen espazioa da. Kanpoko mintza hain iragazkorra izanik eremu horien ioi, azukre eta gainerako metabolito txikien kontzentrazioa zitosolekoaren oso antzekoa da[13]. Tamaina handiko proteinak ordea desberdinak dira, hona iristen direnek seinaleztapen espezifikoa behar baitute kanpoko mintza zeharkatzeko. Proteina hauetako bat C zitokromoa da[12].
Barneko mintza
[aldatu | aldatu iturburu kodea]Konposizio kimikoari dagokionez kanpo mintzarekin alderatuta proteina kopuru handiagoa dauka barnekoak, %80 inguru, hau da, 15 fosfolipido bakoitzeko proteina bat batez beste. Berta daude mitokondrio guztiko proteinen 1/5 eta guztira 151 polipeptido desberdin aurkitu dira[13]. Mintzaren berezitasuna azpimarratzeko mintzetan ezohikoa de kardiolipina izeneko fosfolipidoa ere aurkitzen da[14].
Barneko mintzak edo du porinarik eta iragazkortasun oso hautakorra du. Hainbat proteina mota agertzen dira[13]:
- Fosforilazio oxidatzaileko erredox erreakzioak burutzen dituztenak.
- ATP sintasak.
- Arnas katean parte hartzen dutenak.
- Metabolito espezifikoak garraiatzen dituztenak (pirubatoa, H+, peptidoak...)
Barneko mintzak gandor mitokondrialak ditu. Mitokondrioaren matrizean sartzen diren tolesturak dira eta organulua bera konpartimentalizatzen dute. Gibeleko zeluletan esate baterako mitokondrioen barneko mintza kanpokoa baino bost aldiz luzeagoa da. Bertan arnas kateko proteinak eta ATP sintasak kokatzen dira.
Matrizea
[aldatu | aldatu iturburu kodea]Mitokondrioen matrizea barneko mintzaren barruan geratzen den espazioa da, egitura likidoa duena. DNA, RNA eta erribosomak dauzka, bakterioek dutenen oso antzekoak[13]. Azido nukleiko horiei esker mitokondrioa gai da proteina batzuk bere kabuz sintetizatzeko, gaitasun hori duen zelula-organulu bakarra izanik.
Funtzioak
[aldatu | aldatu iturburu kodea]Mitokondrioak zelularen energia sortzaileak dira, zelulak behar duen ATPa ekoizten dutelako. Beste funtzio batzuk ere betetzen ditu.
Energia ekoizpena
[aldatu | aldatu iturburu kodea]Barneko mintzetako gandorretan dauden proteinek ATP kopuru andanak ekoizteko funtzioa dute. Horretarako arnasketa zelularra burutzen du mitokondrioak.
Zitosolean glukolisia gertatu ondoren pirubatoa mitokondrioaren matrizera sartzen da[2] eta honen deskarboxilazioa burutzen du, lehenik eta behin azetil-CoA bihurtuz eta ondoren Krebsen zikloaren bidez. Prozesu hauetan gertatzen diren oxidazio erreakzioetan NADH+H+ eta FADH2 lortzen dira. Arnasketaren azken fasea barneko mintzeko gandorretan gertatzen da, NADH+H+ eta FADH2 elektroien garraio kati elektroiak ematen dizkiote eta amaieran ATPak ekoizten dira. fosforilazio oxidatiboa deritzo azken prozesu honi[2].
Ekoiztutako ATP hauek mitokondriotik ateratzen dira, aurrena barneko mintzetik proteina espezifikoek garraiatuta eta ondoren kanpoko mintzeko porinen bitartez. Era berean, ADPak kontrako bidea egiten du.
Gantz azidoen beta oxidazioa mitokondrioetan ere burutzen da.
Kaltzio ioien biltegia
[aldatu | aldatu iturburu kodea]Zelulen seinaleztapen prozesuetan kaltzio ioiek ezinbesteko garrantzia dute. Zelularen kaltzioarekiko homeostasia mantentzeko erretikulu endoplasmatikoak biltegi lanak egiten ditu[15] eta mitokondrioak ere laguntzen dio zeregin horretan[16]. Barneko mintzeko garraiatzaile espezifikoen bidez matrizean pilatzen da kaltzioa. Zelulak kaltzioa behar duenean sodio-kaltzio trukatzaileak edo antzeko beste proteina batzuk erabiltzen dira kaltzioa mitokondriatik ateratzeko[17].
Beste funtzio batzuk
[aldatu | aldatu iturburu kodea]- Zelularen apoptosian (heriotza programatua) parte hartzen du[12].
- Mintz potentzialaren erregulazioan parte hartzen du[2].
- Esteroideak sintetizatzen ditu[18].
- Hormona espezifiko batzuei erantzuten die. Mitokondrioko estrogeno hartzaileak (mtER) ditu horretarako. Burmuinetako[19] eta bihotzeko[20] zeluletan gertatzen da bereziki.
Zelula zehatzetan funtzio gehigarriak izan ditzake. Gibeleko zeluletako mitokondrioek amonioaren destoxifikazioa burutzen dute adibidez.
Mitokondrioen jatorria
[aldatu | aldatu iturburu kodea]Mitokondrioek berezko DNA edukitzeak (eta berezko proteinak sortzeak) zientzialariei pentsarazi zien organulu horiek garai batean autonomoak zirela. Teoria horren arabera, mitokondrioak eboluzioaren bitartez zelula eukarioto barnean bizitzera moldatutako bakterioak izango lirateke. Sinbiosi bat eratuko litzateke mitokondrioa eta zelularen artean, zeinean bakterio sartu berriak (mitokondrioak) energia (ATP) emango lioke zelulari, eta honek ingurugiro egonkorra eta elikagaiak mitokondrioari[21].
Teoria endosinbiotiko honen aldeko argudioak badaude:
- mitokondrioek prokarioto handien tamaina dute
- mitokondrioen erribosomak eta bakterioarenak ia berdinak dira[22]
- mitokondrioen DNA biribila da, bakterioen antzekoa[23]
- gandor mitokondrialek eta bakterioen mesosomek antzeko egitura dute
- mitokondrioek autonomia genetikoa dute.
Erreferentziak
[aldatu | aldatu iturburu kodea]- ↑ «Biology: Exploring Life» www.phschool.com (Noiz kontsultatua: 2020-02-16).
- ↑ a b c d Donald Voet. (2006). Fundamentals of biochemistry. Wiley (Noiz kontsultatua: 2020-02-16).
- ↑ (Ingelesez) «mitochondria | Origin and meaning of mitochondria by Online Etymology Dictionary» www.etymonline.com (Noiz kontsultatua: 2020-02-16).
- ↑ a b c d e Ernster L, Schatz G. (1981). Mitochondria: a historical review. The Journal of Cell Biology. 91 (3 Pt 2):, 227-255 or..
- ↑ Altmann, R.. (1890). Die Elementarorganismen und ihre Beziehungen zu den Zellen. Veit, Leipzig.
- ↑ Physiologische Gesellschaft zu Berlin.; Berlin, Physiologische Gesellschaft zu. (1898). Archiv für Physiologie.. Veit & Comp., (Noiz kontsultatua: 2020-02-16).
- ↑ (Alemanez) Meves, Friedrich. (1908-05-01). «Die Chondriosomen als Träger erblicher Anlagen. Cytologische Studien am Hühnerembryo» Archiv für mikroskopische Anatomie 72 (1): 816–867. doi:. ISSN 0176-7364. (Noiz kontsultatua: 2020-02-16).
- ↑ (Ingelesez) «Powerhouse of the Cell» Scientific American (Noiz kontsultatua: 2020-02-16).
- ↑ Martin, William F.; Garg, Sriram; Zimorski, Verena. (2015-09-26). «Endosymbiotic theories for eukaryote origin» Philosophical Transactions of the Royal Society B: Biological Sciences 370 (1678): 20140330. doi:. PMID 26323761. PMC PMC4571569. (Noiz kontsultatua: 2020-02-16).
- ↑ (Ingelesez) Herrmann, Johannes M; Neupert, Walter. (2000-04-01). «Protein transport into mitochondria» Current Opinion in Microbiology 3 (2): 210–214. doi:. ISSN 1369-5274. (Noiz kontsultatua: 2020-02-16).
- ↑ (Ingelesez) Hayashi, Teruo; Rizzuto, Rosario; Hajnoczky, Gyorgy; Su, Tsung-Ping. (2009-02-01). «MAM: more than just a housekeeper» Trends in Cell Biology 19 (2): 81–88. doi:. ISSN 0962-8924. PMID 19144519. (Noiz kontsultatua: 2020-02-16).
- ↑ a b c (Ingelesez) Chipuk, J. E.; Bouchier-Hayes, L.; Green, D. R.. (2006-08). «Mitochondrial outer membrane permeabilization during apoptosis: the innocent bystander scenario» Cell Death & Differentiation 13 (8): 1396–1402. doi:. ISSN 1476-5403. (Noiz kontsultatua: 2020-02-16).
- ↑ a b c d Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P. (1994). Molecular Biology of the Cell. New York: Garland Publishing Inc. ISBN 978-0-8153-3218-3..
- ↑ (Ingelesez) McMillin, Jeanie B; Dowhan, William. (2002-12-30). «Cardiolipin and apoptosis» Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids 1585 (2): 97–107. doi:. ISSN 1388-1981. (Noiz kontsultatua: 2020-02-16).
- ↑ (Ingelesez) Marks, Gaetano Santulli and Andrew R.. (2015-07-31). «Essential Roles of Intracellular Calcium Release Channels in Muscle, Brain, Metabolism, and Aging» Current Molecular Pharmacology (Noiz kontsultatua: 2020-02-16).
- ↑ (Ingelesez) Pizzo, Paola; Pozzan, Tullio. (2007-10-01). «Mitochondria–endoplasmic reticulum choreography: structure and signaling dynamics» Trends in Cell Biology 17 (10): 511–517. doi:. ISSN 0962-8924. PMID 17851078. (Noiz kontsultatua: 2020-02-16).
- ↑ (Ingelesez) Santulli, Gaetano; Pagano, Gennaro; Sardu, Celestino; Xie, Wenjun; Reiken, Steven; D’Ascia, Salvatore Luca; Cannone, Michele; Marziliano, Nicola et al.. (2015-05-01). «Calcium release channel RyR2 regulates insulin release and glucose homeostasis» The Journal of Clinical Investigation 125 (5): 1968–1978. doi:. ISSN 0021-9738. (Noiz kontsultatua: 2020-02-16).
- ↑ (Ingelesez) Rossier, Michel F.. (2006-08-01). «T channels and steroid biosynthesis: in search of a link with mitochondria» Cell Calcium 40 (2): 155–164. doi:. ISSN 0143-4160. (Noiz kontsultatua: 2020-02-16).
- ↑ (Ingelesez) Álvarez-Delgado, Carolina; Mendoza-Rodríguez, C. Adriana; Picazo, Ofir; Cerbón, Marco. (2010-08-01). «Different expression of α and β mitochondrial estrogen receptors in the aging rat brain: Interaction with respiratory complex V» Experimental Gerontology 45 (7): 580–585. doi:. ISSN 0531-5565. (Noiz kontsultatua: 2020-02-16).
- ↑ (Ingelesez) Pavón, Natalia; Martínez-Abundis, Eduardo; Hernández, Luz; Gallardo-Pérez, Juan Carlos; Alvarez-Delgado, Carolina; Cerbón, Marco; Pérez-Torres, Israel; Aranda, Alberto et al.. (2012-10-01). «Sexual hormones: Effects on cardiac and mitochondrial activity after ischemia–reperfusion in adult rats. Gender difference» The Journal of Steroid Biochemistry and Molecular Biology 132 (1): 135–146. doi:. ISSN 0960-0760. (Noiz kontsultatua: 2020-02-16).
- ↑ Margulis, Lynn; Sagan, Dorion. (1986). Origins of sex : three billion years of genetic recombination. New Haven : Yale University Press (Noiz kontsultatua: 2020-02-16).
- ↑ (Ingelesez) O'Brien, Thomas W.. (2003). «Properties of Human Mitochondrial Ribosomes» IUBMB Life 55 (9): 505–513. doi:. ISSN 1521-6551. (Noiz kontsultatua: 2020-02-16).
- ↑ Allen, J. F.; Raven, J. A.; Andersson, G. E.; Karlberg, Olof; Canbäck, Björn; Kurland, Charles G.. (2003-01-29). «On the origin of mitochondria: a genomics perspective» Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences 358 (1429): 165–179. doi:. PMID 12594925. PMC PMC1693097. (Noiz kontsultatua: 2020-02-16).
Ikus, gainera
[aldatu | aldatu iturburu kodea]Kanpo estekak
[aldatu | aldatu iturburu kodea]Datuak: Q39572
Multimedia: Mitochondria / Q39572