Hautespen natural

Hautespen naturala populazio biologiko baten fenotipoen ugalketa diferentzialean datza. Eboluzioaren mekanismo giltzarria da, banako ongi egokituen biziraupena ezartzen duena: populazio batean fenotipo oso desberdin daude; horietako batzuk ez dira ongi egokitzen haien ingurunera, eta desagertuko dira. Ingurune horretara ongi egokitzen direnak izango dira ugaldu eta beren ezaugarri genetikoak transmitituko dituztenak. Charles Darwinek “hautespen natural” terminoa zabaldu zuen, eta hautespen artifizialarekin alderatu zuen, zeina, bere ustez, intentziozkoa baita; hautespen naturala, aldiz, ez.
Organismo-populazio guztietan dago dibertsitatea. Hori, neurri batean, zorizko mutazioak organismo indibidual baten genoman sortzen direlako gertatzen da, eta haren ondorengoek mutazio horiek jaraunts ditzakete. Banakoen bizitzan zehar, haien genomek beren inguruneekin elkarreraginean jarduten dute, ezaugarriak aldatzeko. Genoma baten ingurunean sartzen dira biologia molekularra zelulan, beste zelula batzuk, beste indibiduo batzuk, populazioak, espezieak eta ingurune abiotikoa. Karaktereen zenbait aldaera dituzten indibiduoek beste batzuek baino bizirauteko eta ugaltzeko joera handiagoa dutenez, populazioak eboluzionatu egiten du. Ugalketaren arrakastari eragiten dioten beste faktore batzuk dira hautespen sexuala (gaur egun hautespen naturalean sartua) eta ugalkortasunaren hautapena.
Hautespen naturalak fenotipoan eragiten du, hau da, ingurumenean benetan eragiten duten organismoen ezaugarrietan, baina fenotipo horri ugalketa-abantaila ematen dion genetikaren (herentziazkoa) edozein fenotipo ohikoagoa izan daiteke populazio batean. Denborarekin, prozesu horrek txoko ekologiko partikularretarako espezializatzen diren populazioak sor ditzake (mikroeboluzioa), eta, batzuetan, espezializazio bihur daiteke (espezie berriak agertzea, makroeboluzioa). Bestela esanda, hautespen naturala funtsezko prozesua da populazio baten bilakaeran.
Hautespen naturala biologia modernoaren giltzarria da. Darwinek eta Alfred Russel Wallacek 1858an argitaratutako kontzeptua Darwinen 1859ko liburu eraginkorrean landu zen: Espezieen jatorriaz[1]. Hautespen naturala hautespen artifizialaren antzekotzat deskribatu zuen. Prozesu horren bidez, giza hazleek nahi dituzten ezaugarriak dituzten animaliak eta landareak sistematikoki ugaltzen dira. Hautespen naturalaren kontzeptua, hasiera batean, herentziaren teoria baliagarririk gabe garatu zen; Darwinek liburua idazteko unean, zientziak oraindik genetikari buruzko teoria modernoak garatu behar zituen. Darwinen eboluzionismo tradizionala eta genetika klasikoaren ondorengo aurkikuntzak elkartzeak XX. mendearen erdialdeko sintesi modernoa osatu zuen. Genetika molekularra gehitzeak garapenaren biologia ebolutibora eraman du, eta horrek azaltzen du molekula mailako eboluzioa. Genotipoen ordez, ausazko jito genetikoa erabil daiteke pixkanaka ematen diren aldaketak azaltzeko; hautespen naturalak, berriz, egokitze-eboluzioaren azalpen nagusia izaten jarraitzen du.
Ideiaren garapena
[aldatu | aldatu iturburu kodea]Darwinen aurreko teoriak
[aldatu | aldatu iturburu kodea]
Aro klasikoko zenbait filosofok, Enpedokles[2] eta haren ondorengo intelektual Lukrezio poeta erromatarrak barne[3], ideia hau adierazi zuten: naturak izaki ugari sortzen dituela, zoriz, eta bakarrik irauten dutela beren burua hornitu eta arrakastaz ugaltzea lortzen duten izakiek. Aristotelesek Fisikako II. Liburuan kritikatu zuen Enpedoklesen ideia, hots, beroa eta hotza bezalako kausen funtzionamendu intzidentalaren ondorioz sortzen zirela organismoak.[4] Bere ordez, teleologia naturala planteatu zuen, eta uste zuen forma hori helburu batekin lortu zela, eta froga gisa aipatzen zuen espezieetako herentziaren erregulartasuna.[5][6] Hala ere, bere biologian onartu zuen animalia-mota berriak, munstroak (τερας, teras), oso kasu arraroetan gerta daitezkeela (Animalien Sorkuntza, IV. liburua).[7] Darwinen 1872ko Espezien Jatorria edizioan aipatzen den moduan, Aristotelesek pentsatu zuen hainbat forma (adibidez, hortzak) istripuz agertu zitezkeela, baina forma erabilgarriek bakarrik irauten zutela bizirik:
| « | Horrela, bada, zerk eragozten du atal ezberdinek naturan akzidentalki bakarrik erlazio hau izatea? Hortzak, adibidez, beharrez hazten diren bezala, aurrealde zorrotzak, zatitzeko egokituak, eta ehogailu lauak, janaria murtxikatzeko erabilgarriak, ez baitziren horretarako eginak, istripu baten emaitza baizik. Eta, era berean, xede baterako egokitzapena dagoela dirudien beste zatiei dagokienez. Beraz, gauza guztiak batera (hau da, osotasun baten zati guztiak) zerbaitek eginak balira bezala gertatu ziren heinean, gauza horiek gorde egin ziren, barne-berezkotasun batez egoki eratuak izan zirelarik, eta horrela eratuak izan ez ziren gauza guztiak, galdu egin ziren, eta oraindik ere hil egiten dira. | » |
—Aristoteles, Fisika, II Liburua, 8. atala[8] | ||
Baina Aristotelesek uko egiten dio aukera horri hurrengo paragrafoan, esanez animalien garapenaz hitz egiten ari dela enbrioi gisa "beti edo normalean gertatzen dira" esaldiarekin, ez espezieen jatorriaz:
| « | ... Hala ere, ezinezkoa da ikuspuntu hori egiazkoa izatea. Izan ere, hortzak eta gainerako gauza natural guztiak beti edo normalean modu jakin batean sortzen dira, baina ez da egia zoriaren edo berezkotasunaren ondoriorik dagoenik. Ez diogu ausari edo kointzidentzia hutsari neguko euriaren maiztasuna egozten, baina bai udan maiz egiten duen euria; ezta txakur-egunetako beroa ere, soilik neguan. Gauzak, beraz, kasualitatearen emaitza edo helburu baten emaitza direla, eta horiek kasualitatearen edo bat-batekotasunaren emaitza izan ezin daitezkeela uste bada, orduan ondoriozta daiteke helburu baten bidez izan behar dutela, eta gauza horiek guztiak naturari zor zaizkiola, are gehiago, aztertzen ari garen teoriaren defendatzaileak ere ados leudekeela. Beraz, xede baten araberako ekintza presente dago izatez izatera iristen diren eta diren gauzetan. | » |
—Aristoteles, Fisika, II Liburua, 8. atala[8] | ||
Existentziaren aldeko borroka geroago deskribatu zuen Al-Jahiz idazle islamiarrak IX. mendean.[9][10][11]
Pierre Louis Maupertuisek[12] eta beste batzuek, Darwinen aitona zen Erasmus Darwinek barne, XVIII. mendean berriro sartu zituzten argudio klasikoak.
XIX. mendearen hasierara arte, mendebaldeko gizarteetan iritzi nagusia zen espezie bateko banakoen arteko desberdintasunak sortutako moten ideal platonikoen (edo typusen) interesik gabeko desbideratzeak zirela. Hala ere, geologian aktualismoaren teoriak ideia hau sustatu zuen: indar sinple eta ahulek denbora luzez etengabe jardun zezaketela Lurraren paisaian aldaketa erradikalak eragiteko. Teoria horren arrakastak denbora geologikoaren eskala zabalaz jabetzea eragin zuen, eta hurrengo belaunaldietan aldaketa txiki eta ia hautemanezinek espezieen arteko desberdintasunen eskalan ondorioak eragin zitzaketelako ideia onargarri bihurtu zuen.
XIX. mendearen hasierako Jean-Baptiste Lamarck zoologoak aldaketa ebolutiborako mekanismo gisa hartutako ezaugarrien herentzia iradoki zuen; organismo batek bere bizitzan bereganatutako egokitze-ezaugarriak organismo horren ondorengoek jaraunts zitzaketen, eta, hala, espezieen transmutazioa eragin.[13] Teoria hori, lamarckismoa, Trofim Lysenko biologo sobietarrak XX. mendean defendatu zuen teoria genetikoari aurre egiteko.[14]
1835 eta 1837 bitartean, Edward Blyth zoologoak bariazioa, hautespen artifiziala eta antzeko prozesuak naturan nola gerta daitezkeen landu zuen. Darwinek Blythen ideiak onartu zituen espezieen jatorriaren aldaketari buruzko lehen kapituluan.[15]
Darwinen teoria
[aldatu | aldatu iturburu kodea]1859an, Charles Darwinek hautespen naturalaren bidezko eboluzioaren teoria ezarri zuen egokitzapenerako eta espezializaziorako azalpen gisa. Honela definitu zuen hautespen naturala: "[ezaugarri baten] aldaera txiki bakoitza kontserbatzeko printzipioa, erabilgarria bada".[16] Kontzeptua sinplea zen, baina boteretsua: beren inguruneetara ondoen egokitzen diren gizabanakoak bizirauteko eta ugaltzeko joera handiagoa dute. Haien artean aldaketaren bat dagoen bitartean eta aldaketa hori hereditarioa den bitartean, aldaketa onuragarrienak dituzten banakoen hautaketa saihestezina egongo da. Aldaketak herentzia bidez zabaltzen badira, ugalketaren arrakasta diferentzialak espezie baten populazio partikularren bilakaerara darama, eta behar bezain desberdinak izateko eboluzionatzen duten populazioak espezie desberdin bihurtzen dira.[17][18]

Darwinen ideiak HMS Beagleren bigarren bidaian (1831 – 1836) egindako oharretan oinarritu ziren, eta ekonomialari politiko baten lanean, Thomas Robert Malthus, zeinak, An Essay on the Principles of Population (1798) lanean, adierazi zuen populazioa (ez bada bakarrik kontrolatzen) modu esponentzialean hazten dela, elikagaien eskaintza aritmetikoki hazten den bitartean[19]. Darwinek Malthus irakurri zuenean, 1838an, naturalista gisa bere lana indartsua zela ikusi zuen, naturan ematen den "existentziaren aldeko borroka" azaltzeko. Bere ustez, populazioak baliabideak gainditzen zituen heinean, "aldeko aldakuntzak mantendu egingo lirateke, eta kaltegarriak, suntsitu. Horren emaitza espezie berriak sortzea izango litzateke".[20] Darwinek hau idatzi zuen:
| « | Baldin, bizi-baldintza aldakorretan, izaki organikoek ezberdintasun indibidualak agertzen badituzte beren egituraren ia alde guztietan -eta hau, ene ustez, eztaibadagaitza da-, eta baldin, progresio geometrikoan ugaltzen direlako, adin, urtaro edo urteren batean biziaren aldeko borroka latza gertatzen bada, eta hau ere eztabaidagaitza da, orduan, kontuan harturik izaki organiko guztiek elkarrekin eta beren bizi-baldintzekin dituzten erlazioen amaigabeko konplexutasuna, egituran, osaeran eta azturetan abantailatsua zaien amaigabeko ezberdintasuna daragiena, egitaterik normaletik kanpokoena litzateke izaki bakoitzaren onerako aldakuntzarik inoiz ere ez gertatu izana, gizakiarentzat baliagarri izan diren hainbat aldakuntza gertatu den bezala. Baina izaki organikoren bati baliagarri zaion aldakuntzarik inoiz gertatu bada, aipaturiko ezaugarriak dituzten banakoek biziaren aldeko borrokan salbatzeko aukerarik onena izanen dute ziur, eta herentziaren hastapen ahaltsua bide, antzeko ezaugarridun ondorengoak sortzeko joera izanen dute. Kontserbazio edo egokienen biziraupenaren hastapenari Hautespen Naturala deitu diot. | » |
Behin bere teoria izan zuenean, Darwin oso zorrotza izan zen frogak bildu eta hobetzeko orduan, bere ideia publiko egin aurretik. Bere "liburu handia" idazten ari zen bere ikerketa aurkezteko, Alfred Russel Wallace naturalistak hasiera modu independentean bururatu eta Darwini Charles Lyelli bidali zion saiakera batean deskribatu zuenean. Lyellek eta Joseph Dalton Hookerrek beren saiakera aurkeztea erabaki zuten Darwinek beste naturalista batzuei bidalitako idatzi argitaragabeekin batera, eta espezieek barietateak sortzeko zuten joerari buruz eta barietateak eta espezieak hautatzeko bitarteko naturalen bidez betikotzeari buruz Londresko Sociedad Linneanaren aurrean irakurri zen, hasierako aurkikuntza 1858ko uztailean iragarriz. Darwinek bere proben eta ondorioen zerrenda zehatza argitaratu zuen 1859ko Espezien Jatorriaz liburuaren hirugarren edizioan.[23]

Darwinek hautespen naturalean pentsatu zuenean nekazariek nola hautatzen dituzten hazkuntzarako laboreak edo azienda aztertu zuen, berak "hautespen artifiziala" deitu zion horri; lehen eskuizkribuetan, hautapena egingo zuen "natura" aipatzen zuen. Une hartan, beste eboluzio-mekanismo batzuk, hala nola jito genetikoaren bidezko bilakaera, oraindik ez zeuden esplizituki adierazita, eta Darwinek uste zuen hautespena historiaren zati bat besterik ez zela: "Ziur nago hautespen naturala izan dela aldaketarako bide nagusia, baina ez esklusiboa".[24] Charles Lyelli 1860ko irailean idatzitako gutun batean, Darwinek deitoratu egin zuen "Hautespen naturala" terminoa erabiltzea, "Preserbazio naturala" hitza nahiago zuela.[25]
Darwinentzat eta haren garaikideentzat, hautespen naturala, funtsean, hautespen naturalaren bidezko eboluzioaren sinonimoa zen. Espezieen jatorriari buruzko liburua argitaratu ondoren, hezkuntza minimoa zuten pertsonek onartu zuten eboluzioa nolabait gertatzen zela.[26] Hala ere, hautespen naturalak eztabaidagarria izaten jarraitzen zuen mekanismo gisa, neurri batean jendeari ahulegia iruditzen zitzaiolako organismo bizidunen ezaugarri-sorta azaltzeko, eta, hein batean, eboluzioaren aldekoak ere beren izaera "ez-gidatutik" zein ez progresibotik urruntzen zirelako, hori zenean ideia onartzeko eragozpen bakarra.[27][28] Hala ere, pentsalari batzuek gogo biziz besarkatu zuten hautespen naturala; Darwin irakurri ondoren, Herbert Spencerrek «egokienaren biziraupena» esaldia sortu zuen, teoriaren laburpen ezagun bihurtu zena.[29][30] 1869an argitaratutako Espezieen jatorriari buruzko bosgarren edizioan, Spencerren esaldia hautespen naturalaren alternatiba gisa agertzen zen, emandako kredituarekin: "Baina Herbert Spencer jaunak Egokienaren Biziraupenaren inguruan maiz erabiltzen duen adierazpena zehatzagoa da, eta batzuetan komenigarria ere bada".[31] Nahiz eta oraindik biologo ez direnek esaldia maiz erabiltzen duten, biologo modernoek saihestu egiten dute, tautologikoa baita baldin eta "egokiena" irakurtzen bada "funtzionalki hobea" dela adierazteko, eta indibiduoei aplikatzen zaie, populazioen batez besteko kopurua kontuan hartu ordez.[32]
Sintesi modernoa
[aldatu | aldatu iturburu kodea]Hautespen naturala, funtsean, herentziaren ideian oinarritzen da, baina genetikaren oinarrizko kontzeptuen aurretik garatua. Genetika modernoaren aita zen Gregor Mendel monje moraviarra Darwinen garaikidea zen arren, haren lana ilunpetan zegoen, eta 1900ean bakarrik aurkitu zuten.[33] XX. mendearen hasieran, Mendelen herentzia-legeekin, sintesi modernoa deiturikoarekin, eboluzioaren integrazioarekin, zientzialariek oro har hautespen naturala onartu zuten. Sintesia hainbat arlotako aurrerapenetatik sortu zen. Ronald Fisherrek beharrezko matematika-hizkuntza garatu eta Hautespen Naturalaren Teoria Genetikoa (1930) idatzi zuen.[34] J. B. S. Haldanek hautespen naturalaren "kostua" kontzeptua sartu zuen.[35][36] Sewall Wright-ek hautapenaren eta egokitzapenaren izaera azaldu zuen.[37] Theodosius Dobzhanskyk, Genetika eta Espezieen Jatorria (1937) liburuan, ideia hau ezarri zuen: mutazioak, hautapenaren aurkaritzat ikusi ondoren, hautespen naturalerako lehengaia hornitzen zuela, dibertsitate genetikoa sortuz.[38][39]
Hautespena eta biziraupena
[aldatu | aldatu iturburu kodea]|
|
Artikulu honek erreferentziak behar ditu. Hemen erreferentzia egiaztagarriak gehituz lagun dezakezu. |

Hautespen naturala biziraupen eta birsorkuntzan dauden ezberdintasunak direla-eta sortzen da. Hilkortasun diferentzia deritzo indibiduoen bizirautea euren birsortze adinaren arabera. Mutazioak eta jito genetikoa ausazkoak diren arren, hautespen naturala ez da eta normalean hobekuntzak aurkezten dituzten mutazioak aukeratzen dira. Adibidez, dado bat jaurtikitzen badugu emaitza ausazkoa izango da, baina bi dadoen arteko zenbakirik altuena hartzen badugu hau ez da ausazkoa. Hau dela eta eboluzioa oso lotuta dago ekologiaren ikerketarekin.
Hautespen naturala bi esparrutan bana daiteke: hautespen ekologikoa gertatuko da bizirauten duten izakiak birsortu eta euren geneen ugaritasuna handiagotzen denean gene guztien artean; hautespen sexuala gertatzen da izaki bat beste sexuarekiko interesgarriagoa denean hainbat faktore direla eta, eta horrek ere igoko ditu hainbat generen frekuentziak.
Hautespen naturalak modu ezberdinetan eragiten du mutazioetan. Normalki modurik normalena hautespena egonkortzea da, mutazio txarren frekuentzia txikituz. Beste hautespen mota batek hautespen direkzionala barne hartzen du, mutazio onuragarrien frekuentzia handiagotuz, eta hautespen artifiziala ere hor da, gizakiok hainbat espezieren arraza eta aldaerarekin egiten duguna.
Hautespen naturalaren prozesuarekin organismoak euren ingurunera hobeto moldatzen dira denboran zehar. Hala ere ingurunea aldakorra izan daitekeenez onura horiek beti egongo dira zalantzan eta momentu batean hobekuntza bat dena etorkizunean oztopo bat izan daiteke.
Esan liteke eboluzioaren prozesua mutazioak gertatutakoan hasten dela, horiek baitira genotipo berriak sortzen dituztenak. Genotipo horien gainean jarduten du hautespen naturalak, egokienak (ingurunera hobekien moldatzen direnak) hautatuz eta desegokienak baztertuz. Beraz, mutaziorik gabe ez dago eboluziorik, baina azken finean eboluzioaren norabidea finkatzen duena hautespen naturala da: horrek ez du genotipo berririk sortzen, baina egokienen biziraupena eragiten du.
Terminologia
[aldatu | aldatu iturburu kodea]Hautespen natural terminoa maizago definitzen da herentziazko ezaugarrien gainean lan egiten, horiek zuzenean parte hartzen baitute eboluzioan. Hala ere, hautespen naturala "itsua" da, fenotipoaren aldaketek ugalketa-abantaila eman baitezakete, edozein dela ere ezaugarria, heredagarria izan ala ez. Darwinen erabilera primarioari jarraituz, hautespen itsuaren ondorio ebolutiboa eta haren mekanismoak izendatzeko erabiltzen da terminoa.[26][34][40][41] Batzuetan, baliagarria izaten da hautespen-mekanismoak eta horien ondorioak esplizituki bereiztea; bereizketa hori garrantzitsua denean, zientzialariek "hautespen natural fenotipikoa" definitzen dute, berariaz, "birsortzen diren indibiduoak aukeratzen laguntzen duten mekanismoak", hautapenaren oinarria herentziazkoa den kontuan hartu gabe.[42][43][44] Esaten da organismo baten ugalketa-arrakasta handiena eragiten duten ezaugarriak hautatzen direla, eta arrakasta murrizten dutenek, berriz, aurkako hautespena dutela.[45]
Mekanismoa
[aldatu | aldatu iturburu kodea]Aldaketa hereditarioa, ugalketa diferentziala
[aldatu | aldatu iturburu kodea]
Organismoetako edozein populaziotako indibiduoen artean gertatzen da aldaketa naturala. Desberdintasun batzuek hobekuntza eskaintzen dizkiote organismo batek bizirik irauteko eta ugaltzeko dituen aukerei, halako moldez non ugalketa-tasak gora egiten baituen, eta horrek kume gehiago uzten baitituen. Indibiduo horiei ugaltze-abantaila ematen dieten ezaugarriak ere herentziazkoak badira, hau da, gurasoengandik seme-alabengana pasatzen badira, ugalketa diferentziala izango da, hau da, hurrengo belaunaldian untxi azkarren edo alga eraginkorren proportzio handixeagoa izango da. Ugalketa-abantaila oso txikia bada ere, belaunaldi askotan herentziazko ezaugarri onuragarri guztiak dira nagusi biztanlerian. Horrela, organismo baten inguru naturalak ugalketa-abantaila ematen duten ezaugarriak "hautatzen" ditu, eboluzio-aldaketa eraginez, Darwinek deskribatu zuenez.[46] Honek asmoz egindakoaren itxura du, baina hautespen naturalean ez dago nahitako hautaketarik. Hautespen artifiziala hautespen naturala ez den lekuetan egiten da, nahiz eta biologoek askotan hizkuntza teleologikoa erabiltzen duten hautespen hori deskribatzeko.[47]
Biston betularia kolore argiko zein iluneko motatan agertzen da Britainia Handian, baina industria-iraultzan, sitsak zeuden zuhaitz asko belztu egin zituen kedarrak, eta sits ilunei abantaila eman zieten harrapariengandik ezkutatzeko garaian. Horregatik, kolore iluneko sitsek bizirauteko aukera hobea izan zuten kolore iluneko ondorengoak sortzeko, eta lehen sits iluna ikusi zenetik berrogeita hamar urtera, Manchester industrialeko ia sits guztiak ilunak ziren. Oreka 1956ko Aire Garbiaren Legearen eraginez leheneratu zen, eta sits ilunak berriz ere bakan bihurtu ziren, hautespen naturalak sits hauen bilakaeran duen eragina erakutsiz.[48] Berriki egindako azterlan batek, hegaztien ikusmen-gaitasuna aztertuta, erakusten du likenen koloretik gertuen daudela banako argiak ilunak baino, eta, lehenengo aldiz, sitsen kamuflajea baloratzeko aukera eman zuen.[49]
Egokiera
[aldatu | aldatu iturburu kodea]
Egokiera edo fitness kontzeptua funtsezkoa da hautespen naturalerako. Oro har, organismo "egokituenek" bizirauteko aukera hobea dute, "egokienaren biziraupena" esaldi ezagunean bezala, baina terminoaren esanahi zehatza askoz sotilagoa da. Eboluzioaren teoria modernoak egokiera definitzen du, ez organismo bat zenbat denbora bizi den, ugaltzean duen arrakastaren arabera baizik. Organismo bat bere espezieko beste batzuen denboraren erdia bizi bada, baina helduarora arte bizirik irauten duten ondorengoen bikoitza badu, haren geneak ohikoagoak bihurtzen dira hurrengo belaunaldiko populazio helduetan. Hautespen naturalak banakoei eragiten dien arren, zoriaren ondorioek adierazten dute egokiera "batez beste" soilik definitu daitekeela populazio bateko banakoentzat. Genotipo partikular baten gaitasuna genotipo horrekiko banako guztien batez besteko efektuari dagokio.[50]
"Egokienaren biziraupena" eta "egokienaren hobekuntza" kontzeptuak bereizi behar dira. "Egokienaren biziraupenak" ez du "egokiena hobetzen", populazio baten aldaerarik ahulenak ezabatzea baizik ez du esan nahi. Haldanek adibide matematiko bat eskaintzen du, "egokienaren biziraupena", "hautespen naturalaren kostua" artikuluan.[51] Haldanek "ordezkapen" deitu zion prozesu horri, edo, eskuarki, biologian, "finkapen". Hori ondo deskribatzen da, fenotipoan dauden desberdintasunak direla eta, indibiduoen biziraupen diferentziala eta ugalketa direla eta. Bestalde, "egokitasunaren hobekuntza" ez dago biziraute diferentzialaren eta indibiduoen ugalketaren mende, fenotipoan dauden desberdintasunak direla eta, baizik eta aldaera zehatz horren biziraupen absolutuaren menpe. Populazio bateko kideren batean mutazio onuragarria gertatzeko probabilitatea aldaera horren erreplikazio guztien araberakoa da. Kleinmanek deskribatu zuen egokitasuna hobetzeko matematika.[52] Kishonyren Mega-Plate esperimentuak "egokitasuna hobetzeko" adibide enpiriko bat eman zuen.[53] Esperimentu honetan, "egokitasunaren hobekuntza" aldaera partikularraren erreplikazio-kopuruaren araberakoa da, farmakoaren hurrengo kontzentrazio-eskualde altuenean hazteko gai den aldaera berri bat ager dadin. Finkatzea edo ordezkatzea ez da beharrezkoa "egokitasuna hobetzeko". Bestalde, "egokitasuba hobetzea" ere gerta daiteke "egokienaren biziraupena" gertatzen ari den bitartean. Richard Lenskiren E. coli-ren epe luzeko eboluzio-esperimentu klasikoa ingurune lehiakor bateko egokitzapenaren adibidea da ("egokienaren biziraupenean", "egokitasuna hobetzen" den bitartean).[54] Leinuko kide batean egokitasun hobea emateko mutazio onuragarria gertatzeko probabilitatea motelagoa da lehiakideak badaude. Karga-ahalmen mugatuko ingurune horretan mutazio onuragarria izateko aukera ematen duen aldaerak, lehenik eta behin, "balio txikieneko" aldaerekin lehiatu behar du, behar adina erreplikazio pilatzeko, mutazio onuragarri hori gertatzeko arrazoizko probabilitatea egon dadin.[55]
Lehia
[aldatu | aldatu iturburu kodea]Biologian, lehia organismoen arteko elkarrekintza bat da, non bataren gaitasuna beste baten presentziak murrizten duen. Izan ere, elikagai, ur edo lurralde baten hornidura mugatua da.[56] Lehia espezieen artean edo barruan egon daiteke, eta zuzena edo zeharkakoa izan daiteke.[57] Teorian, lehiatzeko hain egokiak ez diren espezieak egokitu edo desagertu egin beharko lirateke, lehiak rol indartsua baitu hautespen naturalean, baina "alderrai ibiltzeko gelaren" teoriaren arabera, garrantzitsuagoa izan daiteke klado handienen arteko hedapena baino.[57][58]
Lehia r/K hautapen-teoriak modelatzen du. Teoria hori Robert MacArthurrek eta E. O. Wilsonek uhartetako-biogeografiari buruz egindako lanean oinarritzen da.[59] Teoria horretan, hautespen-presioek bi norabide estereotipatuetako baten bilakaera bultzatzen dute: r edo k hautespena.[60] Termino horiek, r eta K, biztanleria-dinamikako eredu logistiko batean irudika daitezke:[61]
non r baita biztanleriaren hazkunde-tasa (N), eta K baita tokiko ingurumenaren karga-ahalmena. Oro har, r hautatutako espezieek nitxo hutsak ustiatzen dituzte eta kume asko sortzen dituzte, bakoitza helduaroan bizirik irauteko probabilitate txiki samarra duena. Aitzitik, K hautatutako espezieek lehiakide sendoak dituzte euren txoko ekologikoan, eta gehiago inbertitzen dute askoz ere kume gutxiagotan, bakoitza helduaroan bizirik irauteko probabilitate handi samarrarekin.[61]
Sailkapena
[aldatu | aldatu iturburu kodea]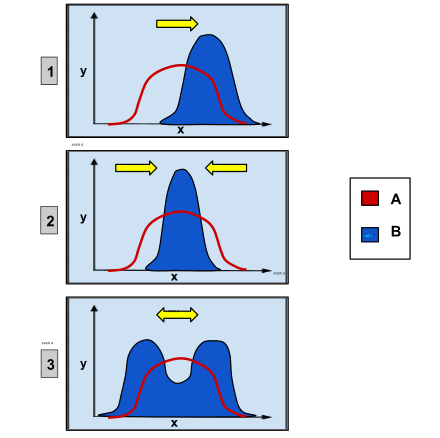
2: hautespen egonkortzailea: erdikoa muturren gainean aldeztua.
3: hautespen disruptiboa: muturrak erdikoaren gainean aldeztuak.
Hautespen naturalak herentziazko edozein karaktere fenotipikoren gainean jardun dezake, eta presio selektiboa ingurumenaren edozein alderdiren ondorioz gerta daiteke, hautespen sexuala eta espezie bereko edo beste batzuetako kideekin duen lehia barne.[62][63] Hala ere, horrek ez du esan nahi hautespen naturala beti norabidekoa denik eta eboluzioa eragiten duenik; hautespen naturala, maiz, statu quo mantentzean lortzen da, hain egokiak ez diren aldaerak kenduz.[46]
Hautespena hainbat modutan sailka daiteke, hala nola ezaugarri batean duen eragina, dibertsitate genetikoa, jarduten duen bizi-zikloaren etapa, hautapen-unitatea edo lehiatzeko baliabidea.
Karaktereen gainean duen ondorioaren arabera
[aldatu | aldatu iturburu kodea]Hautespenak ondorio desberdinak ditu karaktereetan. Hautespen egonkortzailea ezaugarri batek bere egoerarik egonkorrenean mantentzen du, eta, kasurik sinpleenean, optimoaren desbideratze guztiak desabantailatsuak dira selektiboki. Norabide-hautespenak ezaugarri baten muturreko balioei laguntzen die. Oso gutxitan gertatzen den hautespen disruptiboak trantsizio-aldietan ere eragiten du uneko modua azpioptimoa denean, baina norabidea aldatzen du leku batean baino gehiagotan. Bereziki, ezaugarria kuantitatiboa eta unibariatua bada, karaktere altuenak eta baxuenak hobesten dira. Hautespen disruptiboa espezializazioaren aitzindari izan daiteke.[46]
Aniztasun genetikoan duen eraginaren arabera
[aldatu | aldatu iturburu kodea]Hautespena, aukeran, aniztasun genetikoan duen eraginaren arabera bana daiteke. Hautespen garbitzaileak edo negatiboak populazioaren bariazio genetikoa desagerrarazteko jarduten du (eta aurka de novo mutazioa du, aldaketa berriak sartzen dituena.)[64][65] Aitzitik, hautaketaren orekak populazio bateko bariazio genetikoari eusteko jarduten du, baita de novorik ez dagoenean ere, menpeko hautespen negatiboaren bidez. Horretarako mekanismo bat da abantaila heterozigotoa, non bi alelo desberdin dituzten banakoek abantaila selektibo bat baituten alelo bakarra duten banakoen aldean. Horrela azaldu da giza ABO odol-taldearen locus polimorfismoa.[66]

Bizi-zikloaren arabera
[aldatu | aldatu iturburu kodea]Beste aukera bat hautespena bizi-zikloaren etaparen arabera sailkatzea da. Biologo batzuek bi mota bakarrik onartzen dituzte: batetik, bideragarritasun-hautapena (edo biziraupena), organismo batek bizirik irauteko duen probabilitatea handitzeko jarduten duena, eta, bestetik, ugalkortasun-tasa, biziraupena dela-eta ugalketa-tasa handitzeko jarduten duena. Beste batzuek hautaketaren beste osagai batzuetan banatzen dute bizi-zikloa. Beraz, bideragarritasunaren eta biziraupenaren aukeraketa bereizita eta hurrenez hurren defini daiteke: ugalketa-adinaren aurretik eta ondoren bizirik irauteko probabilitatea hobetzeko ekintza; ugalkortasunaren hautapena, berriz, azpiosagai gehigarrietan bana daiteke, hala nola sexu-hautapena, gametoen biziraupenean eta bateragarritasunaren hautaketan eraginez, zigotoen eraketan jardunez.[67]
Hautapen unitatearen arabera
[aldatu | aldatu iturburu kodea]Hautespena mailaren edo hautespen-unitatearen arabera ere sailka daiteke. Banakako hautespenak banakoaren gain eragiten du, hau da, egokitzapenak norbanakoaren onurarako sorten dira, eta gizabanakoen arteko hautespenen emaitza dira. Geneen hautaketak zuzenean eragiten du genearen mailan. Ahaidetasuna hautatzean eta gatazka intragenomikoan, gene mailako hautespenak mendeko prozesuak hobeki azaltzen laguntzen du. Talde hautespenak, halakorik bada, organismo-taldeetan jarduten du, taldeak geneen eta banakoen antzera erreplikatu eta mutatzen dutela proposatuz. Talde hautespenak naturan zenbateraino gertatzen diren eztabaidan dago.[68]
Lehian dagoen errekurtsoaren arabera
[aldatu | aldatu iturburu kodea]
Azkenik, lehia sortzen duen baliabidearen arabera sailka daiteke hautespena. Sexu-hautespena bikotegaia eskuratzeko lehiaren ondorioa da. Sexu-hautespena, eskuarki, ugalkortasun hautespenaren bidez egiten da, batzuetan bideragarritasunaren kontura. Hautespen ekologikoa hautespen naturala da, hautespen sexuala ez den beste edozein bide erabiliz, hala nola ahaidetasunaren hautapena, lehia eta infantizidioa. Darwini jarraiki, hautespen naturala hautespen ekologiko gisa definitzen da batzuetan; kasu horretan, hautespen sexuala mekanismo bereizitzat hartzen da.[70]
Darwinek lehen aldiz artikulatutako sexu-hautespena (indioilarraren isatsaren adibidea erabiliz) bikotea aurkitzeko gaitasunari dagokio bereziki. Lehia hori, berez, sexu bereko banakoen artekoa izan daiteke, arren arteko lehia edo sexu artekoa, non genero batek bikotekidea aukeratzen baitu, sarritan arrak erakusten eta emeak aukeratzen.[71][72] Hala ere, espezie batzuetan, batez ere arrek aukeratzen dute matea, Syngnathidae familiako arrain batzuetan bezala.[73][74]
Ezaugarri fenotipikoak sexu batean ager daitezke eta beste sexuan desiratu, eta fishertar lasterketa izeneko atzeraelikadura-ziklo positiboa eragin dezakete; adibidez, hegazti ar batzuen (indioilarrean, adibidez) lumaje bitxiena.[69] Ronald Fisherrek berak 1930ean proposatutako teoria alternatibo bat seme sexyaren hipotesia da: amek seme-alaba promiskuoak nahi dituzte biloba kopuru handia emateko eta, hartara, seme-alaben guraso promiskuoak aukeratzen dituzte. Sexu bereko kideen arteko erasoa oso ezaugarri bereizgarriekin lotzen da batzuetan, hala nola oreinen adarrekin, beste ar batzuekin borrokatzeko erabiltzen baitira. Oro har, sexuen arteko hautespena dimorfismoarekin lotzen da maiz, espezie bateko arren eta emeen arteko gorputz-tamainan desberdintasunak barne.[72]
Arma-lasterketa
[aldatu | aldatu iturburu kodea]Hautespen naturala ikusten da mikroorganismoetan antibiotikoekiko erresistentziaren garapenean. 1928an penizilina aurkitu zenetik, antibiotikoak gaixotasun bakterianoei aurre egiteko erabili dira. Antibiotikoen erabilera oker orokortuak erabilera klinikoko antibiotikoekiko mikrobio-erresistentzia hautatu du, eta metizilinarekiko erresistentea den Staphylococcus aureus (SARM) "superbakterio" gisa deskribatu da, osasunari eragiten dion mehatxuagatik eta dauden farmakoekiko menderagaiztasun erlatiboagatik[75]. Erantzun-estrategietan antibiotiko desberdinak eta indartsuagoak erabili ohi dira; hala ere, duela gutxi SARM andui berriak agertu dira, sendagai horiekiko erresistenteak direnak[76]. Arma-lasterketa ebolutibo bat da, non bakterioek antibiotikoekiko hain sentikorrak ez diren anduiak garatzen dituzten, eta bitartean, ikertzaile medikoak antibiotiko berriak garatzen saiatzen dira, andui horiekin amaitzeko. Antzeko egoera gertatzen da landareen eta intsektuen pestizidekiko erresistentziarekin. Armamentu-karrerak ez dira gizakiak eragindakoak nahitaez; ondo dokumentatutako adibide bat da gene bat zabaltzea Hypolimnas bolina tximeletan, Wolbachia bakterioaren parasitoek arrak hiltzea eragozten duena Samoako uhartean, non genea bost urteko epean bakarrik hedatu zen[77][78].
Hautespen naturalaren bidezko eboluzioa
[aldatu | aldatu iturburu kodea]Hautespen naturalaren bidezko eboluzioa, karaktere berriak eta espeziazioa eragiteko aurretiazko baldintza bat da heredatu daitekeen aldakuntza genetiko bat egotea, gaitasun-desberdintasunak sorrarazten dituena. Aldakuntza genetikoa mutazioen, birkonbinazio genetikoen eta kariotipoaren alterazioen emaitza da (kromosomen zenbakia, forma, tamaina eta barne-antolaketa). Aldaketa horietako edozeinek eragin oso onuragarria edo oso desabantailatsua izan dezake, baina efektu handiak arraroak dira. Iraganean, material genetikoaren aldaketa gehienak neutrotzat edo ia neutrotzat jotzen ziren, DNA ez-kodifikatzailean gertatzen zirelako edo ordezkapen sinonimoa eragiten zutelako. Hala ere, DNA ez-kodetzailearen mutazio askok ondorio kaltegarriak dituzte[79][80]. Mutazio-tasak eta mutazioek gaitasunean dituzten batez besteko ondorioak organismoaren araberakoak badira ere, gizakien mutazio gehienak zertxobait txarrak dira[81].
Mutazio batzuk "Toolkit" edo gene erregulatzaileetan gertatzen dira. Aldaketa horiek eragin handia izaten dute banakoaren fenotipoan, beste gene askoren funtzioa arautzen dutelako. Gene erregulatzaileetako mutazio gehienek, baina ez guztiek, bideragarriak ez diren enbrioiak sortzen dituzte. Gizakietan mutazio erregulatzaile ez-hilgarri batzuk gertatzen dira HOX geneetan, saihets zerbikal gehigarri bat[82] edo polidaktilia eragin dezaketenak[83], eskuetako edo oinetako behatz kopurua handitzea. Mutazio horiek gaitasun handiagoa ematen dutenean, hautespen naturalak fenotipo horien alde egiten du, eta ezaugarri berria populazioan zabaltzen da. Ezarritako ezaugarriak ez dira aldaezinak; ingurumen-testuinguru batean gaitasun handia duten ezaugarriak askoz ere egokiagoak izan daitezke ingurumen-baldintzak aldatzen badira. Ezaugarri hori gordetzen duen hautespen naturalik ezean, aldakorragoa bihurtzen da eta denborarekin hondatu egiten da, eta, beharbada, ezaugarriaren bestigio bat sortzen da, motxila ebolutibo ere deitua. Egoera askotan, itxuraz aztarna den egiturak funtzionaltasun mugatua gorde dezake, edo beste ezaugarri abantailatsu batzuetarako kooptatua izan daiteke aurre-adaptazio gisa ezagutzen den fenomeno batean. Aztarna-egituraren adibide ospetsu bat, Spalax sator itsuaren begia, fotoperiodikotasunaren pertzepzioan duen funtzioa gordetzen duela uste da[84].
Espeziazioa
[aldatu | aldatu iturburu kodea]Espeziazioak nolabaiteko ugalketa-isolamendua eskatzen du, hau da, gene-fluxua murriztea. Hala ere, espezie kontzeptuaren berezkoa da hibridoen aurkako hautespena egotea, ugalketa-isolamenduaren bilakaeraren aurka eginez, eta Darwinek arazo hori onartu zuen. Arazoa ez da espeziazio alopatrikoan planteatzen, geografikoki bereizitako populazioekin, mutazio multzo desberdinekin dibergitu baitaitezke. E. B. Poulton 1903an konturatu zen ugalketa-isolamenduak dibergentziagatik eboluzionatu zezakeela, leinu bakoitzak gene beraren alelo desberdin eta bateraezina hartzen bazuen. Heterozigotoaren aurkako hautaketak, orduan, zuzenean ugalketa-isolamendua sortuko luke, Bateson-Dobzhansky-Muller modeloa sortuz, geroago H. Allen Orr[85] eta Sergey Gavrilets[86] autoreek egina. Hala ere, errefortzuarekin, hautespen naturalak isolamendu preziatua areagotzea eragin dezake, espeziazio prozesuan zuzenean eraginez.
Oinarri genetikoa
[aldatu | aldatu iturburu kodea]Genotipoa eta fenotipoa
[aldatu | aldatu iturburu kodea]Hautespen naturalak organismo baten fenotipoan edo ezaugarri fisikoetan eragiten du. Fenotipoa organismo baten forma genetikoak (genotipoa) eta organismoa bizi den inguruneak zehazten dute. Populazio bateko organismoek ezaugarri jakin baterako gene baten bertsio desberdinak dituztenean, bertsio horietako bakoitzari alelo deitzen zaio. Aldaketa genetiko horrek eragiten ditu fenotipoaren desberdintasunak. Horren adibide dira gizakietan ABO odol-motako antigenoak, non hiru alelok gobernatzen baitute fenotipoa.[87]
Ezaugarri batzuk gene bakar batek zuzentzen ditu, baina ezaugarri gehienak gene askoren elkarreraginaren ondorio dira. Ezaugarri bat sortzen duten generen batean gertatzen den aldaketa baten efektua txikia izan daiteke fenotipoan; elkarrekin, gene horiek balio fenotipiko posibleen segida bat eragin dezakete.[88]
Hautespen norabidetua
[aldatu | aldatu iturburu kodea]Ezaugarri baten osagairen bat hereditarioa denean, hautespenak aldatu egiten ditu aleloen maiztasunak, edo ezaugarriaren aldaerak sortzen dituen genearen aldaerak. Hautaketa hiru motatan bana daiteke, maiztasun alelikoetan duen eraginaren arabera: norabidezkoa, egonkortzailea eta hautapen disruptiboa.[89] Noranzko hautespena alelo batek beste batzuek baino egokiera hobea duenean gertatzen da; hori dela eta, maizago aukeratzen da, eta gero eta populazioaren zati handiagoak du. Prozesu horrek jarraitu egin dezake aleloa finko egon arte, eta populazio osoa fenotipo egokienarekin bizi da.[90] Askoz ohikoagoa da hautespen egonkortzailea, fenotipoan eragin kaltegarria duten aleloen maiztasuna murrizten baitu, hau da, ezaugarri fisiko txikiagoko organismoak kentzen ditu. Prozesu horrek aleloa populaziotik kanporatu arte jarrai dezake. Hautespenaren egonkortasunak ezaugarri genetiko funtzionalak gordetzen ditu, hala nola proteina gene kodetzaileak edo sekuentzia erregulatzaileak, eta denboraren poderioz, presio selektiboaren bidez, aldagai deletereoen aurka egiten du.[91] Hautespen disruptiboa (edo dibertsifikatzailea) muturreko ezaugarrien balioei mesede egiten dien hautespena da. Hautespen disruptiboak espeziazio sinpatrikoa eragin dezake txokoen zatiketaren bidez.
Hori gertatu daiteke espezie diploideetan (kromosoma-pareak dituztenetan), indibiduo heterozigotoak (aleloaren kopia bakarra dutenak) homozigotoak (bi kopia dituztenak) baino egokiagoak direnean. Horri abantaila heterozigota esaten zaio, eta horren adibiderik ezagunena malariarekiko erresistentzia da anemia faltziformerako heterozigotikoak diren gizakien kasuan. Aldakuntza alelikoari eustea hautespen disruptibo edo dibertsifikatu baten bidez ere gerta daiteke, eta horrek edozein norabidetan batezbestekotik aldentzen diren genotipoen alde egiten du (hau da, gehiegizko dominantziaren aurka doazenen alde), eta ezaugarriaren balioen banaketa bimodala eragin dezake. Azkenik, oreka-hautespena mendeko hautespenaren bidez egin daiteke, non fenotipo jakin baten egokitasuna beste fenotipo batzuek populazioan duten banaketaren araberakoa baita. Jokoen teoriaren printzipioak egoera horietako egokitasunen banaketak ulertzeko erabili dira, bereziki ahaidetasunaren hautapena eta elkarrekiko altruismoaren bilakaera aztertzeko.[92][93]
Hautapena, bariazio genetikoa eta jitoa
[aldatu | aldatu iturburu kodea]Bariazio genetiko guztiaren zati bat funtzionalki neutroa da, eta ez du inolako efektu fenotipikorik edo ezberdintasun esanguratsurik eragiten gaitasunean. Motoo Kimurak jito genetiko bidezko eboluzio molekularrari buruzko teoria neutroak proposatzen du aldaketa horrek behatutako bariazio genetikoaren zati handi bat azaltzen duela[94]. Gertakari neutralek aldaketa genetikoa errotik murritz dezakete populazio botila-lepoen bidez[95], eta horrek, besteak beste, hasieran txikiak ziren populazio berrietan eragin sortzailea eragin dezake[96]. Bariazio genetikoak gaitasun-desberdintasunik eragiten ez duenean, hautespenak ezin dio zuzenean eragin bariazio horren maiztasunari. Ondorioz, leku horietan bariazio genetikoa handiagoa da bariazioak gaitasunean eragina duen lekuetan baino. Hala ere, mutazio berririk gabeko aldi baten ondoren, leku horietako bariazio genetikoa ezabatu egiten da jito genetikoaren ondorioz. Hautespen naturalak bariazio genetikoa murrizten du, egokitu gabeko banakoak eta, beraz, egokitu ezina eragin zuten mutazioak ezabatuz. Aldi berean, mutazio berriak sortzen dira, eta horrek mutazio-hautespen oreka sortzen du. Bi prozesuen emaitza zehatza mutazio berrien abiaduraren araberakoa da, baita hautespen naturalaren indarraren araberakoa ere, azken hau mutazioaren kaltegarritasun mailaren ondorio[97].
Lotura genetikoa gertatzen da bi aleloren kokapena kromosoma batean oso hurbil daudenean. Gametoak sortzen diren bitartean, birkonbinazioak aleloak berrantolatzen ditu. Bi aleloren artean birkonbinazio hori gertatzeko probabilitateak alderantzizko lotura du haien arteko distantziarekin. Hautazko ekorketa populazio batean hautespen positiboaren ondorioz alelo bat ohikoagoa bihurtzen denean gertatzen dira. Alelo baten prebalentzia handitzen den heinean, estuki lotutako aleloak ere ohikoagoak bihur daitezke "autostopismo genetikoaren" bidez, neutroak izan zein apur bat txarrak izanda ere. Ekorketa selektibo handi batek genomaren eskualde bat sorrarazten du, non haplotipoa (aleloa eta bizilagunak), funtsean, populazioan dauden bakarrak diren. Ekorketa selektiboak loturen desoreka neurtuz antzeman daitezke, hau da, haplotipo jakin bat populazioan gainordezkatuta dagoen aztertuz. Ekorketa selektibo batek inguruko aleloak ere hautatzen dituenez, lotura-desoreka handiko bloke bat egoteak blokearen erdigunetik gertu "duela gutxiko" ekorketa selektiboa adieraz lezake[98].
Atzealdeko hautespena ekorketa selektiboaren kontrakoa da. Gune espezifiko batek hautespen araztaile indartsu eta iraunkor bat esperimentatzen badu, lotutako aldakuntza horrekin batera ezabatzeko joera du, aldakortasun orokor txikiko eskualde bat sortuz genoman. Hondoko hautatzea mutazio kaltegarri berrien emaitza denez, edozein haplotipotan ausaz gerta daitezkeenak, ez du sortzen lotura-desorekako bloke argirik, nahiz eta birkonbinazio baxuarekin lotura-desoreka apur bat negatiboa eragin dezakeen, oro har[99].
Eragina
[aldatu | aldatu iturburu kodea]Darwinen ideiek, Adam Smith eta Karl Marxen ideiekin batera, eragin sakona izan zuten XIX. mendeko pentsamenduan, eta haren baieztapen erradikala ere jaso zen: "modu landuan eraikiak, hain desberdinak eta elkarren mendekoak hain modu konplexuan", bizitza-forma sinpleenetatik eboluzionatu zuten printzipio sinple batzuen bidez.[15] Horrek Darwinen aldeko sutsuenetako batzuk inspiratu zituen, eta aurkaritza gogorrena eragin zuen. Hautespen naturalak, Stephen Jay Goulden arabera, "mendebaldeko pentsamenduaren erosotasun sakon eta tradizionalenetako batzuk kentzeko" ahalmena zuen, gizakiek munduan leku berezia dutenaren ustea, besteak beste.[100]
Daniel Dennett filosofoaren hitzetan, hautespen naturalaren bidezko eboluzioaren "Darwinen ideia arriskutsua" "azido unibertsal" bat da, eta ezin da mugatu ezein ontzi edo edukiontzitara, laster urtzen baita, gero eta ingurune zabalagorako bidean.[101] Hala, azken hamarkadetan, hautespen naturalaren kontzeptua biologia ebolutibotik beste diziplina batzuetara hedatu da, hala nola konputazio ebolutiboa, darwinismo kuantikoa, ekonomia ebolutiboa, epistemologia ebolutiboa, psikologia ebolutiboa eta hautespen natural kosmologikoa. Aplikazio mugagabe horri darwinismo unibertsal deritzo[102].
Biziaren jatorria
[aldatu | aldatu iturburu kodea]Bizia materia ez-organikotik abiatzeko modua biologian oraindik konpondu gabeko arazoa da. Hipotesi nabarmena da bizitza, lehenik, RNA polimero autoerreplikatzaile motz gisa agertu zela.[103] Ikuspegi horretatik, bizitza sor daiteke RNAren kateek lehen aldiz hautespen naturala gertatzeko oinarrizko baldintzak izan zituztenean, Charles Darwinek proposatu zuen bezala. Hauek dira baldintzak: herentzia, moten arteko alden arteko lehiengatik. RNAren erreplikatzaile goiztiar baten gaitasuna egokitze-ahalmen intrintsekoen (hau da, sekuentzia nukleotidikoak zehaztutakoak) eta baliabideen eskuragarritasunaren funtzio bat izango zen ziurrenik.[104][105] Hiru egokitze-gaitasun nagusiak, jakina, honako hauek izan zitezkeen: 1) leialtasun moderatuarekin erantzuteko gaitasuna (herentzia eta motaren aldaketa eragiten ditu), 2) gainbehera saihesteko gaitasuna eta 3) baliabideak eskuratu eta prozesatzeko gaitasuna.[104][105] Gaitasun horiek, hasiera batean, RNA erreplikatzaileen konfigurazio tolestuen bidez zehaztuko ziren (erribozima-jarduera duten konfigurazioak barne), eta horiek, era berean, banakako nukleotidoen sekuentzietan kodetuko ziren.[106]
Biologia zelularra eta molekularra
[aldatu | aldatu iturburu kodea]1881ean, Wilhelm Roux enbriologoak Der Kampf der Theile im Organismus (Atalen borroka organismoetan) lana argitaratu zuen. Bertan, organismo baten garapena enbrioiaren atalen arteko lehia darwiniarraren emaitza dela iradoki zuen, maila guztietan gertatzen dena, molekuletatik organoetaraino[107]. Azken urteotan, teoria horren bertsio modernoa proposatu du Jacques Kupiec-ek. Darwinismo zelular horren arabera, molekula-mailako zorizko bariazioak aniztasuna sortzen du zelula-motetan, eta zelula-interakzioek, berriz, ordena bereizgarri bat ezartzen diote garatzen ari den enbrioiari.[108]
Teoria soziala eta psikologikoa
[aldatu | aldatu iturburu kodea]Hautespen naturalaren bidezko eboluzioaren teoriaren ondorio sozialak ere eztabaida etengabearen iturri bihurtu ziren. Friedrich Engels filosofo politiko alemanak, komunismoaren ideologiaren sortzaileetako batek, idatzi zuen 1872an: "Darwinek ez zekien zer satira garratza idatzi zuen gizateriari buruz, eta, bereziki, bere herrikideei buruz, frogatu zuenean lehia askea, existentziaren aldeko borroka, ekonomialariek lorpen historikorik altuena bezala ospatzen dutena, animalien erreinuaren egoera normala dela".[109] Herbert Spencerrek eta Francis Galton eugenistak hautespen naturala nahitaez progresiboa dela interpretatzearen alde zeuden. Hautespen horrek adimena eta zibilizazioa lortzeko ustezko aurrerapenak ekartzen ditu, eta kolonialismoa, eugenesia eta darwinismo sozialerako justifikazio bihurtu zen. Esate baterako, 1940an, Konrad Lorenzek, geroago errefusatu zituen idatzietan, teoria hori erabili zuen Estatu naziaren politiketarako justifikazio gisa.
Hauxe idatzi zuen: "... nekaezintasunagatik, heroismoagatik eta gizarte-onuragatik egindako hautapena giza erakunderen batek egin behar du, baldin eta gizateria, hautatze-faktorerik ezean, ez bada menderatzeak eragindako endekapenak hondatu behar. Arrazaren ideiak, gure estatuaren oinarri gisa, asko lortu du zentzu horretan".[110] Beste batzuek, berriz, gizarteek eta kulturak espezieen eboluzioari aplikatzen zaizkionen antzeko mekanismoen bidez eboluzionatzen duten ideiak garatu dituzte.[111]
Duela gutxi, antropologoen eta psikologoen arteko lanaren ondorioz, soziobiologia garatu da, eta, geroago, psikologia ebolutiboa. Giza psikologiaren ezaugarriak azaltzen saiatzen da eremu hori, antzinako ingurunera egokitzeari dagokionez. Psikologia ebolutiboaren adibiderik nabarmenena, bereziki aurreratua Noam Chomskyren lehen lanetan eta geroago Steven Pinkerren lanean, hipotesi hau da: giza garuna egokitu egin dela hizkuntza naturalaren gramatika-arauak eskuratzeko.[112] Giza portaeraren eta gizarte-egituren beste alderdi batzuk, kultura-arau espezifikoetatik hasi (intzestua saihestea, adibidez) eta genero-rolak bezalako eredu zabalagoetaraino, hipotesi gisa planteatu dira, gizaki modernoek eboluzionatu zuten ingurune goiztiarrerako egokitzapenen antzeko jatorria dutenak. Geneetako hautespen naturalaren ekintzaren analogiaz, honako kontzeptu hauek sortu dira: memeak, "transmisio kulturaleko unitateak" edo hautapenaren eta birkonbinazioaren mende dauden geneen kulturaren baliokideak. Richard Dawkinsek deskribatu zuen lehen aldiz, 1976an, eta, ondoren, Daniel Dennett eta beste filosofo batzuek zabaldu zuten kultura-jarduera konplexuetarako azalpen gisa, giza kontzientzia barne.[113][114]
Informazioaren eta sistemen teoria
[aldatu | aldatu iturburu kodea]1922an, Alfred J. Lotkak proposatu zuen hautespen naturala printzipio fisiko gisa uler zitekeela, sistema batek energia erabiltzeari dagokionez. Kontzeptu hori, gero, Howard T. Odum-ek garatu zuen termodinamikako potentzia maximoaren printzipiotzat, eta, beraz, abantaila selektiboa duten sistema ebolutiboek energia erabilgarriaren transformazio-tasa maximizatzen dute.[115]
Hautespen naturalaren printzipioek askotariko teknika konputazionalak inspiratu dituzte, hala nola bizi artifizial "leuna", zeinak hautatze-prozesuak simulatzen dituen, eta oso eraginkorrak izan baitaitezke erakundeak gaitasun espezifikoko funtzio batek definitutako ingurunera egokitzeko.[116] Adibidez, algoritmo genetiko gisa ezagutzen den optimizazio-algoritmo heuristiko mota batek, 1970eko hamarkadan John Henry Holland-ek aitzindariak eta David E. Goldbergek zabalduak, soluzio optimoak identifikatzen zituen, hasierako probabilitate-banaketa batek definitutako ebazpen-populazio baten erreprodukzio simulatuaren eta mutazioaren bidez.[117][118] Algoritmo horiek bereziki erabilgarriak dira energia-paisaia oso latza duten edo gutxieneko lokal asko dituzten problemei aplikatzen zaizkienean.[119]
Fikzioan
[aldatu | aldatu iturburu kodea]Hautespen naturalaren bidezko eboluzio darwindarra ohikoa da literaturan, gizateriak perfekziorantz nola eboluzionatu dezakeen modu baikorrean hartuta, edo, modu ezkorrean, giza naturaren elkarrekintzaren eta bizirauteko borrokaren ondorio negargarriei dagokienez. Erantzun nagusien artean Samuel Butlerren 1872ko Erewhon ezkorra dago ("inon ere ez", atzeraka idatzia). 1893an, H. G. Wellsek "The Man of the Year Million" imajinatu zuen (milioigarren urteko gizakia), hautespen naturalaren bidez buru eta begi izugarriak zituen eta gorputza uzkurtuta zuen izaki bihurtuta.
Erreferentziak
[aldatu | aldatu iturburu kodea]- ↑ Darwin, Charles, 1809-1882.. (1994). Espezieen jatorria : hautespen naturalaren bidez. (1. argit. argitaraldia) Klasikoak ISBN 8488303734. PMC 434074858. (Noiz kontsultatua: 2019-06-04).
- ↑ Empedocles, 1898
- ↑ Lucretius, 1916
- ↑ Aristoteles,
- ↑ Lear, 1988
- ↑ Henry, Devin. (September 2006). «Aristotle on the Mechanism of Inheritance» Journal of the History of Biology 39 (3): 425–455. doi:..
- ↑ Ariew, 2002
- ↑ a b (Darwin 1872, xiii orr. )
- ↑ Zirkle, Conway. (25 April 1941). «Natural Selection before the 'Origin of Species'» Proceedings of the American Philosophical Society 84 (1): 71–123..
- ↑ Agutter eta Wheatley, 2008
- ↑ Muhammad AS Abdel, Haleem. (2013). «Pre-Darwinian Muslim Scholars' Views on Evolution» Encyclopedia of Sciences and Religions: 74–77..
- ↑ (Frantsesez) Maupertuis, Pierre Louis. (1746). «Les Loix du mouvement et du repos déduites d'un principe metaphysique» Histoire de l'Académie Royale des Sciences et des Belles Lettres: 267–294..
- ↑ Lamarck, 1809
- ↑ Joravsky, David. (January 1959). «Soviet Marxism and Biology before Lysenko» Journal of the History of Ideas 20 (1): 85–104. doi:..
- ↑ a b Darwin, 1859
- ↑ Darwin, 1859, or. 61
- ↑ Darwin, 1859, or. 5
- ↑ Hall, Brian K.. (2008). Strickberger's evolution : the integration of genes, organisms and populations. (4th ed. argitaraldia) Jones and Bartlett ISBN 978-0-7637-0066-9. PMC 85814089. (Noiz kontsultatua: 2022-09-04).
- ↑ Malthus, 1798
- ↑ Darwin, 1958
- ↑ (Darwin 1859, 126–127 orr. )
- ↑ Darwin 1994.
- ↑ Darwin, 1861
- ↑ Darwin, 1859, or. 6
- ↑ Darwin, Charles. (28 September 1860). «Darwin, C. R. to Lyell, Charles» Darwin Correspondence Project (Cambridge University Library) Letter 2931.
- ↑ a b Darwin, 1859
- ↑ Eisley, 1958
- ↑ Kuhn, 1996
- ↑ Darwin, Charles. (5 July 1866). «Darwin, C. R. to Wallace, A. R.» Darwin Correspondence Project (Cambridge University Library) Letter 5145.
- ↑ Stucke, Maurice E.. (Summer 2008). «Better Competition Advocacy» St. John's Law Review 823: 951–1036..—Herbert Spencer, Principles of Biology (1864), vol. 1, pp. 444–445
- ↑ Darwin, 1872
- ↑ Mills, Susan K.; Beatty, John H.. (1979). «The Propensity Interpretation of Fitness» Philosophy of Science 46 (2): 263–286. doi:..
- ↑ Ambrose, Mike. Mendel's Peas. Germplasm Resources Unit, John Innes Centre.
- ↑ a b Fisher, 1930
- ↑ Haldane, 1932
- ↑ Haldane, J. B. S.. (December 1957). «The Cost of Natural Selection» Journal of Genetics 55 (3): 511–524. doi:..
- ↑ Wright, Sewall. (1932). «The roles of mutation, inbreeding, crossbreeding and selection in evolution» Proceedings of the VI International Congress of Genetrics 1: 356–366..
- ↑ Dobzhansky, 1937
- ↑ Dobzhansky, 1951
- ↑ Williams, 1966
- ↑ Endler, 1986
- ↑ Haldane, 1954
- ↑ Lande, Russell; Arnold, Stevan J.. (November 1983). «The Measurement of Selection on Correlated Characters» Evolution 37 (6): 1210–1226. doi:. PMID 28556011..
- ↑ Futuyma, 2005
- ↑ Sober, 1993
- ↑ a b c Evolution and Natural Selection. University of Michigan 10 October 2010.
- ↑ «Teleological Notions in Biology» Stanford Encyclopedia of Philosophy 18 May 2003.
- ↑ van't Hof, Arjen E.; Campagne, Pascal; Rigden, Daniel J. (June 2016). «The industrial melanism mutation in British peppered moths is a transposable element» Nature 534 (7605): 102–105. doi:. PMID 27251284. Bibcode: 2016Natur.534..102H..
- ↑ Walton, Olivia; Stevens, Martin. (2018). «Avian vision models and field experiments determine the survival value of peppered moth camouflage» Communications Biology 1: 118. doi:. OCLC .6123793 PMID 30271998..
- ↑ Orr, H. Allen. (August 2009). «Fitness and its role in evolutionary genetics» Nat Rev Genet 10 (8): 531–539. doi:. OCLC .2753274 PMID 19546856..
- ↑ Haldane, J. B. S.. (November 1992). «The Cost of Natural Selection» Current Science 63 (9/10): 612–625..
- ↑ Kleinman, A.. (2014). «The basic science and mathematics of random mutation and natural selection» Statistics in Medicine 33 (29): 5074–5080. doi:. PMID 25244620..
- ↑ Baym, M.; Lieberman, T. D.; Kelsic, E. D.; Chait, R.; Gross, R.; Yelin, I.; Kishony, R.. (2016). «Spatiotemporal microbial evolution on antibiotic landscapes» Science 353 (6304): 1147–51. doi:. OCLC .5534434 PMID 27609891. Bibcode: 2016Sci...353.1147B..
- ↑ Blount, Zachary D.; Borland, Christina Z.; Lenski, Richard E.. (2008). «Historical contingency and the evolution of a key innovation in an experimental population of Escherichia coli» Proceedings of the National Academy of Sciences 105 (23): 7899–906. doi:. OCLC .2430337 PMID 18524956. Bibcode: 2008PNAS..105.7899B..
- ↑ Good, B. H.; Rouzine, I. M.; Balick, D. J.; Hallatschek, O.; Desai, M. M.. (27 February 2012). «Distribution of fixed beneficial mutations and the rate of adaptation in asexual populations» Proceedings of the National Academy of Sciences 109 (13): 4950–4955. doi:. OCLC .3323973 PMID 22371564..
- ↑ Begon, Townsend eta Harper, 1996
- ↑ a b Sahney, Sarda; Benton, Michael J.; Ferry, Paul A.. (23 August 2010). «Links between global taxonomic diversity, ecological diversity and the expansion of vertebrates on land» Biology Letters 6 (4): 544–547. doi:. OCLC .2936204 PMID 20106856..
- ↑ Jardine, Phillip E.; Janis, Christine M.; Sahney, Sarda; Benton, Michael J.. (1 December 2012). «Grit not grass: Concordant patterns of early origin of hypsodonty in Great Plains ungulates and Glires» Palaeogeography, Palaeoclimatology, Palaeoecology 365–366: 1–10. doi:. Bibcode: 2012PPP...365....1J..
- ↑ MacArthur eta Wilson, 2001
- ↑ Pianka, Eric R.. (November–December 1970). «On r- and K-Selection» The American Naturalist 104940: 592–597. doi:..
- ↑ a b (Frantsesez) Verhulst, Pierre François. (1838). «Notice sur la loi que la population suit dans son accroissement» Correspondance Mathématique et Physique 10: 113–121. OCLC .490225808.
- ↑ Zimmer eta Emlen, 2013
- ↑ Miller, 2000
- ↑ Lemey, Salemi eta Vandamme, 2009
- ↑ Loewe, Laurence. (2008). «Negative Selection» Nature Education (Nature Publishing Group) OCLC .310450541.
- ↑ Villanea, Fernando A.; Safi, Kristin N.; Busch, Jeremiah W.. (May 2015). «A General Model of Negative Frequency Dependent Selection Explains Global Patterns of Human ABO Polymorphism» PLOS ONE 10 (5): e0125003. doi:. OCLC .4422588 PMID 25946124. Bibcode: 2015PLoSO..1025003V..
- ↑ a b Christiansen, 1984, or. 65–79
- ↑ Wade, Michael J.. (2010). «Multilevel and kin selection in a connected world» Nature 463 (7283): E8–E9. doi:. OCLC .3151728 PMID 20164866. Bibcode: 2010Natur.463....8W..
- ↑ a b Greenfield, M.D.; Alem, S.; Limousin, D.; Bailey, N.W.. (2014). «The dilemma of Fisherian sexual selection: Mate choice for indirect benefits despite rarity and overall weakness of trait-preference genetic correlation» Evolution 68 (12): 3524–3536. doi:. PMID 25308282..
- ↑ Mayr, 2006
- ↑ Andersson, 1994
- ↑ a b Hosken, David J.; House, Clarissa M.. (January 2011). «Sexual Selection» Current Biology 21 (2): R62–R65. doi:. PMID 21256434..
- ↑ Eens, Marcel; Pinxten, Rianne. (5 October 2000). «Sex-role reversal in vertebrates: behavioural and endocrinological accounts» Behavioural Processes 51 (1–3): 135–147. doi:. PMID 11074317..
- ↑ Barlow, George W.. (March 2005). «How Do We Decide that a Species is Sex-Role Reversed?» The Quarterly Review of Biology 80 (1): 28–35. doi:. PMID 15884733..
- ↑ (Ingelesez) «MRSA superbug found in supermarket pork raises alarm over farming risks» the Guardian 2015-06-18 (Noiz kontsultatua: 2022-09-04).
- ↑ (Ingelesez) Schito, G. C.. (2006-01-01). «The importance of the development of antibiotic resistance in Staphylococcus aureus» Clinical Microbiology and Infection 12: 3–8. doi:. ISSN 1198-743X. (Noiz kontsultatua: 2022-09-04).
- ↑ (Ingelesez) Charlat, Sylvain; Hornett, Emily A.; Fullard, James H.; Davies, Neil; Roderick, George K.; Wedell, Nina; Hurst, Gregory D. D.. (2007-07-13). «Extraordinary Flux in Sex Ratio» Science 317 (5835): 214–214. doi:. ISSN 0036-8075. (Noiz kontsultatua: 2022-09-04).
- ↑ Moran, Gregory J.; Krishnadasan, Anusha; Gorwitz, Rachel J.; Fosheim, Gregory E.; McDougal, Linda K.; Carey, Roberta B.; Talan, David A.. (2006-08-17). «Methicillin-Resistant S. aureus Infections among Patients in the Emergency Department» New England Journal of Medicine 355 (7): 666–674. doi:. ISSN 0028-4793. PMID 16914702. (Noiz kontsultatua: 2022-09-04).
- ↑ Kryukov, Gregory V; Schmidt, Steffen; Sunyaev, Shamil. (2005-06-30). «Small fitness effect of mutations in highly conserved non-coding regions» Human Molecular Genetics 14 (15): 2221–2229. doi:. ISSN 1460-2083. (Noiz kontsultatua: 2022-09-04).
- ↑ Bejerano, Gill; Pheasant, Michael; Makunin, Igor; Stephen, Stuart; Kent, W. James; Mattick, John S.; Haussler, David. (2004-05-01). «Ultraconserved Elements in the Human Genome» Science 304: 1321–1325. doi:. ISSN 0036-8075. (Noiz kontsultatua: 2022-09-04).
- ↑ academic.oup.com doi:. PMID 16547091. PMC PMC1526495. (Noiz kontsultatua: 2022-09-04).
- ↑ Galis, F.. (1999-04-15). «Why do almost all mammals have seven cervical vertebrae? Developmental constraints, Hox genes, and cancer» The Journal of Experimental Zoology 285 (1): 19–26. ISSN 0022-104X. PMID 10327647. (Noiz kontsultatua: 2022-09-04).
- ↑ (Ingelesez) Zákány, József; Fromental-Ramain, Catherine; Warot, Xavier; Duboule, Denis. (1997-12-09). «Regulation of number and size of digits by posterior Hox genes: A dose-dependent mechanism with potential evolutionary implications» Proceedings of the National Academy of Sciences 94 (25): 13695–13700. doi:. ISSN 0027-8424. PMID 9391088. PMC PMC28368. (Noiz kontsultatua: 2022-09-04).
- ↑ Sanyal, S.; Jansen, H. G.; de Grip, W. J.; Nevo, E.; de Jong, W. W.. (1990-07). «The eye of the blind mole rat, Spalax ehrenbergi. Rudiment with hidden function?» Investigative Ophthalmology & Visual Science 31 (7): 1398–1404. ISSN 0146-0404. PMID 2142147. (Noiz kontsultatua: 2022-09-04).
- ↑ Orr, H Allen. (1996-12-01). «D<scp>obzhansky</scp>, B<scp>ateson</scp>, and the Genetics of Speciation» Genetics 144 (4): 1331–1335. doi:. ISSN 1943-2631. PMID 8978022. PMC PMC1207686. (Noiz kontsultatua: 2022-09-04).
- ↑ Gavrilets, Sergey. (2004). Fitness landscapes and the origin of species. Princeton University Press ISBN 0-691-11758-6. PMC 56023319. (Noiz kontsultatua: 2022-09-04).
- ↑ McKusick, Victor A.; Gross, Matthew B.. (18 November 2014). «ABO Glycosyltransferase; ABO» Online Mendelian Inheritance in Man (National Library of Medicine).
- ↑ Falconer eta Mackay, 1996
- ↑ Rice, 2004, See especially chapters 5 and 6 for a quantitative treatment
- ↑ Rieseberg, L.H.; Widmer, A.; Arntz, A.M.; Burke, J.M.. (2002). «Directional selection is the primary cause of phenotypic diversification» PNAS 99 (19): 12242–12245. doi:. OCLC .129429 PMID 12221290. Bibcode: 2002PNAS...9912242R..
- ↑ «A neo-Darwinian commentary on macroevolution» Evolution 36 (3): 474–498. 1982 doi:. PMID 28568049..
- ↑ Hamilton, William D.. (July 1964). «The genetical evolution of social behaviour. II» Journal of Theoretical Biology 7 (1): 17–52. doi:. PMID 5875340. Bibcode: 1964JThBi...7...17H..
- ↑ Trivers, Robert L.. (March 1971). «The Evolution of Reciprocal Altruism» The Quarterly Review of Biology 46 (1): 35–57. doi:..
- ↑ Kimura, Motoo. (1983). The neutral theory of molecular evolution. Cambridge University Press ISBN 0-521-23109-4. PMC 9081989. (Noiz kontsultatua: 2022-09-07).
- ↑ Genetics. Macmillan Reference USA 2003 ISBN 0-02-865890-6. PMC 55983868. (Noiz kontsultatua: 2022-09-07).
- ↑ Campbell, Neil A.. (1996). Biology. (4th ed. argitaraldia) Benjamin/Cummings Pub. Co ISBN 0-8053-1940-9. PMC 33333455. (Noiz kontsultatua: 2022-09-07).
- ↑ (Ingelesez) Lynch, Michael. (2010-08-01). «Evolution of the mutation rate» Trends in Genetics 26 (8): 345–352. doi:. ISSN 0168-9525. PMID 20594608. PMC PMC2910838. (Noiz kontsultatua: 2022-09-07).
- ↑ (Ingelesez) Smith, John Maynard; Haigh, John. (1974-02). «The hitch-hiking effect of a favourable gene» Genetics Research 23 (1): 23–35. doi:. ISSN 1469-5073. (Noiz kontsultatua: 2022-09-07).
- ↑ (Ingelesez) Keightley, Peter D.; Otto, Sarah P.. (2006-09). «Interference among deleterious mutations favours sex and recombination in finite populations» Nature 443 (7107): 89–92. doi:. ISSN 1476-4687. (Noiz kontsultatua: 2022-09-07).
- ↑ Gould, Stephen Jay. (12 June 1997). «Darwinian Fundamentalism» The New York Review of Books 44 (10).
- ↑ Dennett, 1995
- ↑ Sydow, Momme von. (2012). From darwinian metaphysics towards understanding the evolution of evolutionary mechanisms : a historical and philosophical analysis of gene-darwinism and universal darwinism. Universitätsverlag Göttingen c/o SUB Göttingen ISBN 978-3-86395-006-4. PMC 794698133. (Noiz kontsultatua: 2022-09-07).
- ↑ Eigen, Manfred; Gardiner, William; Schuster, Peter; Winkler-Oswatitsch, Ruthild. (April 1981). «The Origin of Genetic Information» Scientific American 244 (4): 88–92, 96, et passim. doi:. PMID 6164094. Bibcode: 1981SciAm.244d..88E..
- ↑ a b Bernstein, Harris; Byerly, Henry C.; Hopf, Frederick A.; Michod, Richard A.; Vemulapalli, G. Krishna. (June 1983). «The Darwinian Dynamic» The Quarterly Review of Biology 582: 185–207. doi:..
- ↑ a b Michod, 1999
- ↑ Orgel, Leslie E.. (1987). «Evolution of the Genetic Apparatus: A Review» Cold Spring Harbor Symposia on Quantitative Biology 52: 9–16. doi:. PMID 2456886..
- ↑ Roux 1881.
- ↑ Kupiec, Jean-Jacques. (3 May 2010). «Cellular Darwinism (stochastic gene expression in cell differentiation and embryo development)» SciTopics.
- ↑ Engels, 1964
- ↑ Eisenberg, Leon. (September 2005). «Which image for Lorenz?» American Journal of Psychiatry 162 (9): 1760. doi:. PMID 16135651.. Eisenberg quoting translation of Durch Domestikation verursachte Störungen arteigenen Verhaltens (1940, p. 2) by Konrad Lorenz.
- ↑ Wilson, 2002
- ↑ Pinker, 1995
- ↑ Dawkins, 1976
- ↑ Dennett, 1991
- ↑ Lotka, Alfred J.. (June 1922). «Contribution to the energetics of evolution» PNAS 8 (6): 147–151. doi:. OCLC .1085052 PMID 16576642. Bibcode: 1922PNAS....8..147L..
- ↑ Kauffman, 1993
- ↑ Goldberg, 1989
- ↑ Mitchell, 1996
- ↑ «Genetic Algorithms» Pharmacological Sciences 7 November 2016.
Bibliografia
[aldatu | aldatu iturburu kodea]- Agutter, Paul S.; Wheatley, Denys N.. (2008). Thinking about Life: The History and Philosophy of Biology and Other Sciences. Springer Science+Business Media ISBN 978-1-4020-8865-0. OCLC .304561132.
- Andersson, Malte. (1994). Sexual Selection. Princeton University Press ISBN 978-0-691-00057-2. OCLC .28891551.
- Ariew, André. (2002). Functions: New Essays in the Philosophy of Psychology and Biology. Oxford University Press ISBN 978-0-19-824103-4. OCLC .48965141.
- Aristoteles. Physics. The Internet Classics Archive OCLC .54350394.
- Begon, Michael; Townsend, Colin R.; Harper, John L.. (1996). Ecology: Individuals, Populations and Communities. Blackwell Science ISBN 978-0-632-03801-5. OCLC .32893848.
- Christiansen, Freddy B.. (1984). Evolutionary Ecology: The 23rd Symposium of the British Ecological Society, Leeds, 1982. Blackwell Scientific Publications ISBN 978-0-632-01189-6. OCLC .12586581. Modified from Christiansen by adding survival selection in the reproductive phase.
- Darwin, Charles. (1859). On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life. John Murray OCLC .741260650. The book is available from The Complete Work of Charles Darwin Online. Retrieved 2015-07-23.
- Darwin, Charles. (1861). On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life. John Murray OCLC .550913.
- Darwin, Charles. (1872). The Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life. John Murray OCLC .1185571.
- Darwin, Charles. (1958). The Autobiography of Charles Darwin, 1809–1882: With original omissions restored; Edited and with Appendix and Notes by his grand-daughter, Nora Barlow. Collins OCLC .869541868.
- Darwin, Charles. (1994). Espezieen jatorria : hautespen naturalaren bidez. (1. argit. argitaraldia) Klasikoak ISBN 84-88303-73-4. PMC 434074858. (Noiz kontsultatua: 2022-09-04).
- Dawkins, Richard. (1976). The Selfish Gene. Oxford University Press ISBN 978-0-19-857519-1. OCLC .2681149.
- Dennett, Daniel C.. (1991). Consciousness Explained. Little, Brown and Company ISBN 978-0-316-18065-8. OCLC .23648691.
- Dennett, Daniel C.. (1995). Darwin's Dangerous Idea: Evolution and the Meanings of Life. Simon & Schuster ISBN 978-0-684-80290-9. OCLC .31867409.
- Dobzhansky, Theodosius. (1937). Genetics and the Origin of Species. Columbia University Press OCLC .766405.
- Dobzhansky, Theodosius. (1951). Genetics and the Origin of Species. Columbia University Press OCLC .295774.
- Eisley, Loren. (1958). Darwin's Century: Evolution and the Men Who Discovered It. Doubleday OCLC .168989.
- Empedocles. (1898). The izena Philosophers of Greece. Kegan Paul, Trench, Trübner & Co. Ltd. OCLC .1376248. Txantiloi:Internet Archive.
- Endler, John A.. (1986). Natural Selection in the Wild. Princeton University Press ISBN 978-0-691-08386-5. OCLC .12262762.
- Engels, Friedrich. (1964). Dialectics of Nature. Progress argitaletxeas OCLC .807047245. The book is available from the Marxist Internet Archive.
- Falconer, Douglas S.; Mackay, Trudy F.C.. (1996). Introduction to Quantitative Genetics. Longman ISBN 978-0-582-24302-6. OCLC .824656731.
- Fisher, Ronald Aylmer. (1930). The Genetical Theory of Natural Selection. The Clarendon Press OCLC .493745635.
- Futuyma, Douglas J.. (2005). Evolution. Sinauer Associates ISBN 978-0-87893-187-3. OCLC .57311264.
- Goldberg, David E.. (1989). Genetic Algorithms in Search, Optimization and Machine Learning. Addison-Wesley Publishing Company ISBN 978-0-201-15767-3. OCLC .17674450.
- Haldane, J B.S.. (1932). The Causes of Evolution. Longmans, Green & Co. OCLC .5006266. "This book is based on a series of lectures delivered in January 1931 at the Prifysgol Cymru, Aberystwyth, and enizenburuad 'A re-examination of Darwinism'."
- Haldane, J.B.S.. Atti del IX Congresso Internazionale di Genetica, Bellagio (Como) 24–31 agosto 1953. University of Florence OCLC .9069245.
- Kauffman, Stuart. (1993). The Origins of Order: Self-Organisation and Selection in Evolution. Oxford University Press ISBN 978-0-19-507951-7. OCLC .23253930.
- Lamarck, Jean-Baptiste. (1809). Philosophie Zoologique. Dentu et L'Auteur OCLC .2210044. Txantiloi:Internet Archive.
- Lear, Jonathan. (1988). Aristotle: The Desire to Understand. Cambridge University Press ISBN 978-0-521-34762-4. OCLC .16352317.
- Kuhn, Thomas S.. (1996). The Structure of Scientific Revolutions. University of Chicago Press ISBN 978-0-226-45808-3. OCLC .34548541.
- The Phylogenetic Handbook: A Practical Approach to Phylogenetic Analysis and Hypothesis Testing. Cambridge University Press ISBN 978-0-521-73071-6. OCLC .295002266.
- Lucretius. (1916). De rerum natura. Tufts University OCLC .33233743.
- MacArthur, Robert H.; Wilson, Edward O.. (2001). The Theory of Island Biogeography. Princeton University Press ISBN 978-0-691-08836-5. OCLC .45202069.
- Malthus, Thomas Robert. (1798). An Essay on the Principle of Population, As It Affects the Future Improvement of Society: with Remarks on the Speculations of Mr. Godwin, M. Condorcet, and Other Writers. J. Johnson OCLC .65344349. The book is available here from Frank Elwell, Rogers State University.
- Mayr, Ernst. (1942). Systematics and the Origin of Species from the Viewpoint of a Zoologist. Columbia University Press OCLC .766053.
- Mayr, Ernst. (2006). Sexual Selection and the Descent of Man: The Darwinian Pivot. AldineTransaction ISBN 978-0-202-30845-6. OCLC .62857839.
- Michod, Richard A.. (1999). Darwinian Dynamics: Evolutionary Transitions in Fitness and Individuality. Princeton University Press ISBN 978-0-691-02699-2. OCLC .38948118.
- Miller, Geoffrey. (2000). The Mating Mind: How Sexual Choice Shaped the Evolution of Human Nature. Doubleday ISBN 978-0-385-49516-5. OCLC .43648482.
- Mitchell, Melanie. (1996). An Introduction to Genetic Algorithms. MIT Press ISBN 978-0-262-13316-6. OCLC .42854439.
- Pinker, Steven. (1995). The Language Instinct: How the Mind Creates Language. Harper Perennial ISBN 978-0-06-097651-4. OCLC .670524593.
- Rice, Sean H.. (2004). Evolutionary Theory: Mathematical and Conceptual Foundations. Sinauer Associates ISBN 978-0-87893-702-8. OCLC .54988554.
- Roux, Wilhelm. (1881). Der Kampf der Theile im Organismus. Wilhelm Engelmann OCLC .8200805. Txantiloi:Internet Archive Retrieved 2015-08-11.
- Sober, Elliott. (1993). The Nature of Selection: Evolutionary Theory in Philosophical Focus. University of Chicago Press ISBN 978-0-226-76748-2. OCLC .896826726.
- Wallace, Alfred Russel. (1871). Contributions to the Theory of Natural Selection. A Series of Essays. Macmillan & Co. OCLC .809350209.
- Williams, George C.. (1966). Adaptation and Natural Selection: A Critique of Some Current Evolutionary Thought. Princeton University Press ISBN 978-0-691-02615-2. OCLC .35230452.
- Wilson, David Sloan. (2002). Darwin's Cathedral: Evolution, Religion, and the Nature of Society. University of Chicago Press ISBN 978-0-691-02615-2. OCLC .48777441.
- Zimmer, Carl; Emlen, Douglas J.. (2013). Evolution: Making Sense of Life. Roberts and Company argitaletxeas ISBN 978-1-936221-17-2. OCLC .767565909.
Ikus, gainera
[aldatu | aldatu iturburu kodea]Kanpo estekak
[aldatu | aldatu iturburu kodea]| Autoritate kontrola |
|
|---|
Datuak: Q43478
Multimedia: Natural selection / Q43478