Arctodus
| Arctodus | ||
|---|---|---|
| Rango temporal: 2,5 Ma - 0,01 Ma Plioceno - Holoceno | ||
 Esqueleto de A. simus de Rancho La Brea | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Subfilo: | Vertebrata | |
| Clase: | Mammalia | |
| Subclase: | Theria | |
| Infraclase: | Placentalia | |
| Orden: | Carnivora | |
| Suborden: | Caniformia | |
| Familia: | Ursidae | |
| Subfamilia: | Tremarctinae | |
| Género: |
Arctodus † Leidy, 1854 | |
| Especie tipo | ||
|
Arctodus pristinus Leidy, 1854 | ||
| Distribución | ||
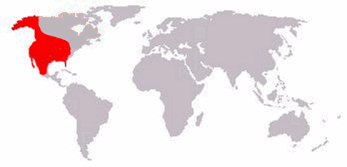 Rango de Arctodus simus | ||
| Especies | ||
| ||
| Sinonimia | ||
Sinónimos a nivel de especie A. simus:
A. pristinus: Arctodus haplodon Hay 1902
Arctotherium pristinum Cope 1895 Ursus haplodon Cope 1896 Tremarctos haplodon Trouessart 1897 | ||
Arctodus es un género extinto de mamífero carnívoro de la familia de los úrsidos, conocidos popularmente como osos de cara corta u osos bulldog, que habitó América del Norte desde finales del Plioceno hasta inicios de la época del Holoceno, entre unos 2.5 millones de años a hace 10,000 años. Era el más común de los osos tremarctinos en Norteamérica. Se reconocen dos especies: el oso de cara corta menor (Arctodus pristinus) y el oso gigante de cara corta (Arctodus simus), siendo considerado este último como uno de los mayores mamíferos terrestres carnívoros que hayan existido. Se ha hipotetizado que su extinción coincide con el periodo de enfriamiento global conocido como el Dryas Reciente que comenzó unos 12,900 años antes del presente.
Taxonomía
[editar]Estas especies parecen haber tenido un hocico desproporcionadamente más corto que el de varios de los osos modernos, lo que llevó a darles su apodo de "cara corta." Esta apariencia de longitud breve es una ilusión causada por los hocicos altos y los huesos nasales cortos que tienen los osos tremarctinos en comparación con los osos ursinos; Arctodus tenía un hocico más alto pero de igual longitud a las de los osos actuales. Esta característica es también compartida por el único oso tremarctino vivo, el oso de anteojos.[1] El cráneo además tiene un rostro amplio y acortado, lo que le da una apariencia vagamente parecida a la de un félido.[2] El nombre científico del género, Arctodus, se deriva del idioma griego, y significa "diente de oso". Los primeros fósiles de A. simus se encontraron en la cueva Potter Creek, en el condado de Shasta, en California.[3][4] El esqueleto más completo de A. simus hallado en Estados Unidos se descubrió en Indiana; los huesos están en el Museo Field de Historia Natural, Chicago.[5][6]
Evolución
[editar]
Arctodus pertenece al grupo de osos conocidos como los tremarctinos, los cuales aparecieron en América del Norte durante el inicio de la época del Mioceno tardío siendo representados por Plionarctos, un género considera como ancestro de los Tremarctinae. Plionarctos habría dado origen a varias formas de tamaño mediano como Arctodus pristinus, Tremarctos floridanus y Arctotherium sp. durante la época del Plioceno superior en América del Norte.[7][8][9] Tanto Arctodus como Tremarctos estuvieron mayormente restringidos a las zonas más boscosas de la parte oriental del continente, ya que se piensa que el cánido Borophagus y el oso Agriotherium habrían limitado la presencia de los tremarctinos en las zonas más abiertas del oeste norteamericano. Tremarctos floridanus establecería un rango que abarca mayormente el Golfo de México (aunque también se extendió por California e Idaho),[9] mientras que Arctodus pristinus se dispersó desde Aguascalientes en México,[10] hasta Nueva Jersey ya en Estados Unidos.[11] Quizás debido a su historia evolutiva, tanto Tremarctos floridanus como Arctodus pristinus tienen la mayor concentración de fósiles en Florida; en particular, en el yacimiento 1 del río Santa Fe en el condado de Gilchrist. No obstante, ya para el inicio del Cuaternario, cuando se extinguieron tanto Borophagus como Agriotherium, Arctodus habría tomado la oportunidad y se extendería en el resto del continente, principalmente con la especie A. simus. Simultáneamente, durante el Gran Intercambio Biótico Americano que seguiría a la unión de América del Norte y del Sur, el género Arctotherium presente en Centroamérica invadiría el sur,[8] llevando a experimentar una notable diversificación, incluyendo a la colosal especie A. angustidens.
A comienzos del Pleistoceno medio (edad kansaniense, hace ~800,000 años), el pequeño A. pristinus sería acompañado por el enorme A. simus.[9] Las dos especies no solo se diferencian por su tamaño, sino también por el hocico más corto, los dientes más robustos y las extremidades más largas de A. simus y las proporciones relativas de los premolares y molares de cada especie. Sin embargo, hay relativamente pocas diferencias morfológicas. Como resultado, diferenciar a Arctodus simus de Arctodus pristinus puede ser difícil, ya que los mayores individuos de Arctodus pristinus son comparables en tamaño a los menores ejemplares de Arctodus simus.[9] Aunque ambas especies coexistieron por al menos medio millón de años (A. pristinus se extinguió hace ~300 000 años),[9] no hay hasta ahora en el registro fósil evidencia directa de simpatría o competencia entre ambos.[9] Debe señalarse que hay algunos registros no confirmados de A. pristinus en Carolina del Sur, California y Florida del Pleistoceno tardío, lo que podría sugerir una posible supervivencia como una especie relicta en ambientes de refugio hasta el evento de extinción del Cuaternario.[12][13][14] En todo caso, mientras que A. pristinus parece haber preferido los ambientes de enclave de bosques densos cálidos del este de Norteamérica,[15] A. simus era más cosmopolita, llegando a ocupar amplias áreas y compartiendo su rango de distribución con el oso negro americano y el oso pardo desde hace unos 100 000 años.[14]

A. simus ocupaba un rango geográfico desde el norte de Alaska hasta el centro de México por el oeste, desde Pensilvania y Florida en el este,[9][16][17][18][19] siendo particualrmente famosos los fósiles de la especie hallados en Rancho La Brea en el sur de California.[20] El Pleistoceno superior representa el pico de la diversidad de los úrsidos en el Cuaternario de América, con los osos pardos, osos negros americanos, Tremarctos floridanus, Arctodus simus y Arctotherium wingei todos recorriendo los territorios situados al sur de la capa de hielo Laurentino,[21][22] mientras los osos polares estaban presentes en las zonas de casquetes polares.[23][24] A pesar del amplio rango temporal y geográfico de Arctodus simus, sus restos fósiles son comparativamente raros (hasta 109 hallazgos reportados para 2010, en yacimientos por lo demás bien estudiados).[9][25] Esto sin embargo no implica que no se presentara diversidad genética en Arctodus, ya que las muestras de material genético de ejemplares de la cueva Chiquihuite de México indican una amplia divergencia con los hallazgos de Arctodus provenientes del Yukón.[25]
Descripción
[editar]Tamaño
[editar]Algunos individuos de A. simus pueden haber sido los mayores representantes terrestres del orden Carnivora que hayan vivido en América del Norte. En un estudio publicado en 2010, se estimó la masa de seis especímenes de A. simus; un tercio de estos pesaba cerca de 900 kilogramos, siendo el mayor de ellos un ejemplar de Utah que alcanzaría los 957 kilogramos, lo que sugiere que los osos de esta especie de grandes tamaños serían más comunes de lo que se había pensado. El rango de peso calculado para todos los ejemplares examinados estaba entre 957 kg a 317 kg, con un peso promedio de ~750 kg.[1] Hay mucha variación en el tamaño de los adultos, lo cual podría ser debido a la escasez de hallazgos, el dimorfismo sexual y los potenciales ecomorfos que podrían estar aumentando ese tamaño promedio para Arctodus.[26] Los mayores individuos procedentes del Rancho La Brea son mucho más pequeños que muchos especímenes de Alaska, Utah y Nebraska. Se ha sugerido que sería resultado de una diferenciación ecomorfológica (si bien los especímenes de La Brea tienen una variación de tamaño del 25%, como podría esperarse en el dimorfismo sexual de un úrsido), o bien de subespecies, con A. s. yukonensis ocupando las partes norte y central de su rango, mientras que A. s. simus se distribuye en el resto.[1] Nuevamente, el bajo número de ejemplares y las muestras sesgadas por sexos hacen dudar de esta designación, junto con el hallazgo de un Arctodus simus de Florida dentro del rango de tamaño de A. s. yukonensis, en un territorio bien adentrado del supuesto rango de A. s. simus.[9]
Aunque se conocen cerca de 100 yacimientos con fósiles de osos de cara corta gigantes en América del Norte, solo en un sitio se ha encontrado un báculo (el hueso del pene) que podría pertenecer a Arctodus simus. La carencia de báculos encontrados de Arctodus probablemente refleja tanto la tafonomía como el compartimiento. La mayoría de los restos de esqueleto son de sitios abiertos en donde usualmente solo se encuentran algunos cuantos huesos. En contraste, los pasajes horizontales en las cuevas aparecen individuos pequeños relativamente completos para los cuales sería esperable que se pudiera encontrar el báculo, de haber estado presente. El tamaño relativamente pequeño y la falta de este hueso en los depósitos de cuevas sugiere que eran las hembras de A. simus las que las usaban como madrigueras, en línea con el comportamiento maternal de los úrsidos.[27][28] Por lo tanto, esto en conjunto con el dimorfismo sexual de los úrsidos (por ejemplo, en los osos de anteojos los machos son 30%-40% más grandes que las hembras) implica que los mayores individuos serían generalmente machos, particularmente los de mayor edad, mientras que los menores corresponden a hembras.[9][29]

Al incorporarse en sus patas traseras, Arctodus simus mediría 2,4 - 3 metros de altura.[30] Al caminar en sus cuatro patas, A. simus mediría entre 1,5 - 1,8 metros de altura hasta los hombros, por lo que sería lo suficientemente alto para ver a un humano adulto a los ojos. Los mayores individuos de A. simus pueden haberse aproximado a los 1,000 kg,[2] o incluso a los 1200 kg de peso, hipotéticamente.[31] En la cueva Riverbluff en Misuri, se han encontrado una serie de marcas de garras de unos 4,6 metros de alto a lo largo de las paredes de la misma, indicando que fueron hechas por un individuo de A. simus que habría medido 3,7 metros de alto.[32][33]
Anatomía
[editar]Con respecto a su locomoción, si bien la forma de la articulación del codo sugiere que Arctodus, Arctotherium bonariense y Arctotherium wingei posiblemente habrían retenido las adaptaciones para trepar a los árboles, el mismo tamaño del codo indica que Arctodus no habría sido arborícola. Ya que el epicóndilo medial presentaba una expansión notable en estas especies, es más probable que como ocurre con el oso panda, los extintos Arctodus y Arctotherium conservaran este rasgo para tener un alto grado de destreza en las extremidades delanteras. La evolución convergente de estos géneros hacia un tamaño corporal grande pudo haber sido favorecido por las adaptaciones para consumir cadáveres,[34] por lo que sus extremidades muy móviles pueden haber sido ventajosas y se habrían preservado en el linaje de los Tremarctinae a pesar de su tendencia en el aumento de tamaño.[35]
Un estudio publicado en 2010 determinó que sus patas no eran proporcionalmente más largas que las de los osos modernos como se esperaba que fueran, y que de hecho los osos en general son animales de patas alargadas aunque esto queda disimulado por el propio contorno del cuerpo y el pelaje. El estudio concluye que la supuesta apariencia "patilarga" de estos osos es mayormente una ilusión óptica causada por el torso y espalda relativamente cortos del animal, proporciones que comparten con las hienas actuales.[1][36]
Paleobiología
[editar]Arctodus pristinus
[editar]Las marcas de mordidas en perezosos y proboscídeos juveniles en restos hallados en Leisey Shell Pit en Florida encajan con el tamaño de los dientes caninos de Arctodus pristinus. Se desconoce si estas marcas son el resultado de depredación activa o de consumo de carroña.[37] En la zona del yacimiento del río Santa Fe 1 que corresponde al Pleistoceno temprano (edad mamífero del Blanquense) en Florida (hace 2.2-1.8 millones de años) en el que habitó Arctodus pristinus, la evidencia indica que era un ambiente mayormente de praderas abiertas. Había presencia de dolinas kársticas y manantiales, como los de la Florida actual. Arctodus pristinus pudo haber coexistido con especies de megafauna como las aves del terror (Titanis), félidos dientes de sable (Xenosmilus), perezosos gigantes (Eremotherium, Glossotherium y Megalonyx), armadillos gigantescos (Holmesina, Glyptotherium), gonfotéridos (Rhynchotherium y ¿Cuvieronius?), hienas (Chasmoporthetes), cánidos (Borophagus y Canis lepophagus), pecaríes (Platygonus), llamas (Hemiauchenia), antilocápridos (Capromeryx), y caballos de tres dedos (Nannippus). La fauna de menor tamaño incluía cóndores, rállidos y patos entre otras aves pequeñas, roedores tales como puercoespines, lagartos, serpientes, aligátores, tortugas y artrópodos.[38][39] La evolución de Arctodus simus, la competencia con Tremarctos floridanus y los osos negros (los cuales solo aparecen en Florida en el Pleistoceno superior),[7] y posiblemente la transición durante el Pleistoceno en Florida de hábitats de bosques con clima cálido y muy húmedo a un bioma cálido pero mucho más seco y abierto, son considerados como los factores relevantes en la desaparición gradual de Arctodus pristinus durante el Pleistoceno medio (hace 300 mil años).[9][14] Arctodus pristinus también ha sido hallado en Kansas, Carolina del Sur, Maryland, Pensilvania y Nueva Jersey en Estados Unidos, y Aguascalientes en México.[7][11][13]
Arctodus simus
[editar]Paleoecología
[editar]
A pesar de una amplia gama de ideas, los académicos han concluido en su mayoría que Arctodus simus habría sido un omnívoro enorme y oportunista, con una dieta flexible y adaptada a la localidad, como ocurre con el oso pardo.[1][40][41][42] Aparte de aprovechar la carroña de los cadáveres de megaherbívoros, y la depredación ocasional de bisontes, ciervos, camélidos y perezosos terrestres los cuales eran abundantes a lo largo del rango geográfico de Arctodus,[1] Arctodus simus habría consumido grandes cantidades de vegetación allí donde estuviera disponible,[36] si es que Arctodus simus no era principalmente herbívoro.[7][43] Al preferir generalmente hábitats abiertos y mixtos, Arctodus simus era particularmente abundante en el oeste de Norteamérica. No obstante, con base en la amplia distribución de la especie, esta habría habitado en condiciones climáticas muy diversas y en toda clase de ambientes, desde los bosques boreales y la estepa de mamut en el norte, las planicies y los bosques montanos en el interior, hasta los bosques y sabanas subtropicales del sur.[9][15][43][44]
Se ha pensado que pudo haber una posible interacción entre Smilodon y Arctodus, sobre la base de la supuesta falta de habilidad del félido dientes de sable para consumir en gran medida a sus presas, concentrándose principalmente en tejidos blandos, habría dejado grandes cantidades de comida disponible para otros animales carnívoros tales como Arctodus. Este animal habría tenido el potencial de ser un cleptoparásito, disputando las presas a los depredadores contemporáneos tales como lobos gigantes, Smilodon, Homotherium, pumas, doles, lobos grises, jaguares, guepardos americanos, leones americanos, leones de las cavernas (en el Yukón) y otros osos. Sin embargo, muchos de estos animales habrían sido gregarios y capaces de defender con éxito sus capturas, y la propia variación de tamaño de Arctodus simus habría limitado la frecuencia de este comportamiento a principalmente los ejemplares de mayor tamaño.[1] A través del rango de esta especie, se encontrarían varios grandes herbívoros como los proboscídeos (el mamut lanudo en el norte, Stegomastodon, Cuvieronius y el mamut colombino en el sur y el mastodonte americano en varias zonas), camélidos (Camelops, Hemiauchenia y Palaeolama), équidos (Equus y Haringtonhippus), cingulados (Glyptodon, Pachyarmatherium, Pampatherium y Holmesina), tapires, perezosos (Eremotherium, Nothrotheriops, Paramylodon y Megalonyx), bisontes, notoungulados (Mixotoxodon) ciervos (Cervalces, Odocoileus), varios caprinos, osos hormigueros, varios antilocápridos, capibaras, pecaríes, el saiga y el castor gigante Castoroides. Los datos preliminares sugieren que determinados hábitats eran más óptimos para las poblaciones de Arctodus simus, como la estepa del mamut en Beringia, los lagos pluviales y las estepas de artemisias en la Gran Cuenca, los bosques de alta montaña en las tierras altas del interior de Estados Unidos, y las sabanas mixtas del sureste de Estados Unidos y la Mesa del Centro en México.[14][45]
Carnívoro oportunista
[editar]La evidencia de un individuo de Arctodus simus de Cedral, en San Luis Potosí, México, con base en valores de isótopos de oxígeno, sugieren que la especie en ese lugar prefería áreas de vegetación cerrada. Al tener solo una muestra de un individuo de esa zona y por la carencia de valores de isótopos de nitrógeno, en este estudio se encontró difícil inferir si Arctodus simus era un omnívoro o un hipercarnívoro. Sin embargo, el valor δ13C mostró que este ejemplar se alimentaba de herbívoros que consumían plantas C3, (aunque pudo haber consumo de vegetación C3 por el mismo Arctodus) los cuales eran diferentes de aquellos consumidos por los depredadores especializados en hábitats abiertos como el león americano y el lobo gigante Aenocyon. El valor de isótopos de oxígeno de Arctodus no se equipara pero si es cercano a los valores de los tapires y Hemiauchenia. Estos animales entonces pudieron haber sido incluidos en su dieta, junto con otros herbívoros que consumían la vegetación C3 (por ejemplo, aquello que ramoneaban hojas, raíces, frutas, tallos y flores) como Camelops, Platygonus, Nothrotheriops y el mastodonte americano. La zona alojaba un bosque de galería cerca de una sabana o zonas de arbustos, con un clima húmedo subtropical. Este mosaico de sabana y bosque soportaba una gran diversidad de mamíferos herbívoros y carnívoros, siendo similar a la hallada en los mesohábitats del Pleistoceno superior en el noroeste de Sonora y el Suroeste de Estados Unidos,[44][46][47] estableciendo un bioma ideal para Arctodus.
Para los especímenes de Alaska y de California (Fairmead Landfill, durante el Irvingtoniense), los estudios publicados en 2008 y en 2012 propusieron que Arctodus simus comería mamuts, caballos, bisontes y ciervos. Sin embargo, en otros sitios de California (como los pozos de brea de McKittrick y Asphalt Seep) Arctodus simus tendría una dieta basada en ciervos y tapires, parecida a la inferida para el individuo de Cedral.[44][48]
Un hallazgo realizado en el islas del Canal de California repleto de firmas de isótopos que se corresponden con las de bisontes y camélidos (seguidas de las de pinípedos) reafirma la sugerencia sobre que aunque no era totalmente carnívoro, Arctodus pudo haber tenido una dieta flexible a lo largo de su rango. El que este fósil de Arctodus de las islas fuera probablemente transportado post-mortem desde las costas californianas complica más la idea de una dieta generalizada para Arctodus.[49]

El último sobreviviente del linaje de los osos tremarctinos o de cara corta es el oso de anteojos, endémico de las zonas andinas. Aunque es mayormente herbívoro, esta especie es un depredador activo ocasionalmente. El oso de anteojos tiene varias técnicas de caza, principalmente sorprendiendo o subyugando a sus presas, montándose sobre su espalda y consumiendo al animal inmovilizado aún estando vivo, mientras lo sujeta con su peso y sus largas garras. De forma alternativa, el oso de anteojos persigue a sus presas hacia terrenos irregulares, faldas de montañas o precipicios, provocando su caída y en ocasiones su muerte. Una vez muerta, la presa es arrastrada a un lugar seguro (como un área de bosque) y consumida, dejando solo restos del esqueleto.[50] Estos comportamientos podrían ser aplicables para los tremactinos gigantes como Arctotherium y Arctodus.
Competencia con los osos pardos
[editar]Los osos pardos, junto con los leones, bisontes y zorros rojos, emigraron por primera vez hacia América del Norte a través de Beringia durante la glaciación illinoiense (hace unos ~170 mil años).[21] Arctodus simus puede haber sido sobrepasado por estos inmigrantes a medida que estos se expandían hacia el sur desde el este de Beringia, y gradualmente se establecieron en América del Norte. Ambas especies se han reportado conjuntamente en Alaska antes de hace 34 mil años antes del presente, y en depósitos posteriores del Pleistoceno en California y en Nevada. Sin embargo, datos de la isla de Vancouver sugieren que ambas especies ocuparon la isla en rápida sucesión el uno del otro a finales del Pleistoceno, Arctodus simus por la fecha de hace ∼11.7 miles de años, y el oso pardo justo antes 12,440 ± 35 y 12,425 ± 30 años antes del presente, y posteriormente Arctodus simus (11,100 ± 30 antes del presente).[42]
Con solo un par de cientos de años separándolos, un estudio de 2018 concluyó que ambas especies estuvieron presentes en la isla de Vancouver en esa época pero sin compartir los mismos territorios. Esto podría ser interpretado como una diferenciación de nicho, para reducir la competencia por el territorio y el uso de cuevas en estas especies simpátricas. Aunque los cambios postglaciales en los recursos disponibles pueden haber ocasionado también la desaparición de los osos pardos en Vancouver, estos a diferencia de Arctodus simus persistieron en las zonas adyacentes.[42]
El consumo de carne en numerosos ejemplares de Arctodus simus de Beringia está confirmado por los elevados valores de isótopos (δ13C y δ15N) en las zonas donde los osos pueden haber competido por comida, pero al parecer usualmente ocuparon un nivel trófico más alto comparados con los osos pardos invasores. El que Arctodus simus pueda haber excluido a los osos pardos del este de Beringia entre hace ∼34 a ∼20 miles de años (que se correlaciona con la expansión de las turberas) sugiere que Arctodus típicamente habría sido dominante sobre los osos pardos en esa zona. Una vez que Arctodus se extinguió en Beringia hace 20 mil años, los osos pardos recolonizarían toda Beringia, pero con dietas más carnívoras que las de sus contrapartes anteriores a la fecha de hace 34 mil años. Esto reforzaría la idea de que estos osos competían por recursos y nichos similares.[21][42]
Ya en una escala continental, el estudio de 2018 explicó que aunque los osos pardos y A. simus fueran simpátricos durante el tiempo en que los primeros colonizaban Norteamérica, Arctodus simus habría sido por lo general quien dominara las interacciones de competencia, particularmente cuando sus poblaciones eran robustas, y desplazaron a los osos pardos de localidades específicas. La razón por la que probablemente los osos pardos persistieron al final del Pleistoceno y no así Arctodus, sería que este último eran menos flexible al adaptarse a las nuevas condiciones ambientales que cambiaban rápidamente y que impactaron la disponibilidad o calidad de la comida y los posibles hábitats.[42]
Hibernación
[editar]De acuerdo con un estudio de 2003, los fósiles de A. simus se han encontrado casi exclusivamente en cuevas en zonas de karst. En los Estados Unidos, 26 de 69 yacimientos con Arctodus simus (~38%) corresponden a cuevas. El que más de un tercio de todos estos yacimientos sean cuevas sugiere una estrecha relación entre estos osos y los ambientes de cavernas. Más aún, cerca del 70% de los especímenes más pequeños (alguna vez asignados a la subespecie A. s. simus) son precisamente de depósitos de cuevas. Ninguno de los especímenes asignados al morfo más grande (A. s. yukonensis) procede de pasajes de cavernas. Tomando en cuenta el hecho de que los úrsidos hembras son menores y tienden a refugiarse en cuevas, parece lógico concluir que la mayoría de restos de Arctodus simus de esos depósitos eran hembras y que pueden haber estado hibernando cuando perecieron.[27] En cuanto a análogos modernos, las hembras de osos negros americanos y osos pardos en climas fríos ingresan en sus madrigueras antes que los machos y permanecen por más tiempo allí. Las hembras de osos polares junto las hembras de osos pardos y osos negros americanos en las zonas más cálidas de su distribución, por lo general entran en estados de dormancia, torpor y cuidado materno durante el invierno, mientras que los machos continúan activos durante todo el año.[29] Se infiere que los ejemplares hembras de Arctodus simus habrían presentado el comportamiento de refugiarse para cuidar a sus crías, aunque no es claro el tipo de metabolismo (hibernación/torpor) que adoptaban.[28] Sin embargo, a la fecha no se conocen registros de adultos con crías asociadas en los yacimientos con cuevas.[29]
Se conocen lechos de fósiles de osos que con frecuencia preservan restos de Arctodus simus asociados a osos negros americanos del Pleistoceno y de la subespecie moderna (U.a. amplidens y U. a. americanus); estos depósitos se han encontrado en Misuri y Oklahoma. Se asume que estos depósitos mixtos son el resultado de la acumulación a lo largo del tiempo de osos individuales (incluyendo a Arctodus) que murieron durante el invierno.[51]
Discusiones sobre la dieta
[editar]La hipótesis del "superdepredador"
[editar]
Una propuesta sobre la ecología potencial de A. simus lo visualiza como un depredador tosco que subyugaba a los mamíferos grandes del Pleistoceno con su gran fuerza física.[52] Sin embargo, se ha sugerido que sus extremidades, a pesar de ser muy grandes, serían también demasiado gráciles para ese tipo de ataques,[53][54] significativamente más que los miembros de Arctotherium angustidens.[31]
Dado que sus largas extremidades le habrían permitido a Arctodus correr a velocidades de 50 - 70 km/h, una hipótesis alternativa es que puede haber cazado persiguiendo a herbívoros del Pleistoceno como los caballos y el antílope saiga, e incluso mamuts, una idea que en su momento le ganó el apodo de "oso corredor".[1][53][55] Sin embargo, durante la persecución de presas veloces, la propia masa física del oso y su postura plantígrada habrían sido una desventaja; los osos pardos modernos pueden correr a velocidades similares pero se cansan rápidamente y no pueden mantener la persecución por largo tiempo. Los esqueletos de Arctodus no se articulan de modo que permitieran hacer giros rápidos – una habilidad requerida por cualquier depredador que sobreviva al cazar presas ágiles.[30][53]
Por otra parte, algunos análisis de las extremidades delanteras de Arctodus sugieren que este oso estaba en las fases iniciales de la evolución de una locomoción cursorial, al tener más tendencia hacia la carrera, se piensa que esta especie era capaz de una locomoción más eficiente en línea recta y con mayor velocidad en comparación con los osos actuales, y posiblemente estando más adaptado a perseguir presas grandes que los actuales osos pardos y polares.[54] No obstante, la carencia de adaptaciones claras para ser depredador como la ausencia de caninos comprimidos lateralmente, y el tener dientes carnasiales más aptos para aplastar y moler que para trocear carne, hace dudar de la interpretación de Arctodus simus como un hipercarnívoro.[35][36] Si bien el único úrsido actual que es hipercarnívoro, el oso polar, también carece de carnasiales cortantes, en esa especie su principal medio de subsistencia es la grasa de sus presas y no la mucho más gruesa carne, lo que puede haber inhibido la necesidad de evolucionar una dentadura especializada en procesar carne (aunque también hay que señalar que la evolución de los osos polares a partir de los osos pardos es un fenómeno relativamente reciente).[36][43]
Cleptoparásito especializado vs omnívoro
[editar]

Arctodus se movería eficientemente en una marcha de paso moderadamente rápido, más especializada que la de los osos modernos. El gran tamaño corporal, las largas patas delanteras, la espalda corta y en pendiente de Arctodus se combinarían para tener eficiencia locomotora, ya que estas características incrementaban la cantidad de energía de tensión elástica disponible en los tendones e incrementaban la longitud de las zancadas, haciendo de Arctodus un animal mejor constituido para la resistencia que para la velocidad.[30][53] De acuerdo con estos argumentos, A. simus estaba mal equipado para ser un depredador activo, llevando a algunos investigadores a concluir que era un cleptoparásito,[30] habiendo evolucionado en un carroñero especializado adaptado a cubrir territorios enormes a fin de incrementar las probabilidades de localizar los cadáveres de grandes animales que estarían dispersos y distribuidos de forma irregular, y que podría dominar este recurso alimenticio atractivo pero impredecible.[36] Bajo este modelo, existiría una presión selectiva adicional para el incremento de su tamaño, de modo que Arctodus pudiera aprovechar y defender las piezas de carroña de otros grandes carnívoros, algunos de los cuales eran gregarios, o perseguirlos para alejarlos de sus presas y robar su comida.[53] Además de esto, el rostro corto sería el resultado del incremento de las fuerzas de los músculos involucrados en cerrar la mandíbula (el músculo temporal y el masetero), que pudieron ser una adaptación para romper huesos con sus anchos carnasiales. Dicho uso de sus dientes P4 (cuarto premolar superior) y m1 (primer molar inferior) se basa en el extenso desgaste de estos dientes en los individuos viejos de Arctodus simus y en Agriotherium (otro oso de gran tamaño).[36]
Esta idea fue cuestionada en un estudio de 2013 del microdesgaste de los dientes de varias especies de osos extintos y modernos (concretamente analizando especímenes de Arctodus de los pozos de La Brea), y por un estudio de 2015 enfocado en los carnívoros encontrados en Rancho La Brea.[56][57] Las hienas que están más especializadas en alimentarse de carroña muestran patrones característicos de daño en los molares al triturar huesos. Con base en la carencia de desgaste "rompehuesos" en los especímenes de Rancho La Brea, los investigadores concluyeron que Arctodus simus no era un carroñero especializado. Al compararse con los osos actuales, esta población de A. simus mostró un patrón de desgate dental muy similar al de su pariente vivo más cercano, el oso de anteojos, el cual tiene una dieta muy variada, siendo un omnívoro y en ocasiones con una dieta casi enteramente herbívora.[43] Esto depende de la disponibilidad regional y estacional de recursos.[56] En cambio, se identificaron varias fracturas en las coronas dentales e infecciones alveolares en el oso suramericano gigante Arctotherium angustidens. Estas se interpretaron como evidencia de consumo de materiales duros (por ejemplo, huesos), que indican tentativamente que este oso comía en forma regular cadáveres de ungulados obtenidos mediante el cleptoparasitismo. Estas patologías dentales no son observadas en cambio en A. simus, salvo por las fuertes facetas de desgaste en los individuos viejos.[1] Además, la relativa carencia de restos de Arctodus en las zonas de trampas de depredadores en sitios como Rancho La Brea sugiere que Arctodus no competía por los cadáveres, si bien este yacimiento ha producido más especímenes de A. simus que cualquier otro sitio, esto podría relacionarse con la calidad de la preservación de la brea y en todo caso solo llegan a ser el 1% de todos los carnívoros hallados en esos pozos,[57] añadiendo más dudas sobre la hipótesis de ser un carroñero.[45] Las patologías dentales que se han encontrado, tales como el cálculo dental supragingival en un individuo joven,[27] y cavidades asociadas con el consumo de carbohidratos de individuos de La Brea, son evidencia adicional de una dieta omnívora para Arctodus simus.[45]

Un posible modelo actual para su ecomorfología serían la hiena rayada y la hiena parda. Arctodus simus se asemeja a estos hiénidos y a la depredadora hiena manchada, por la forma del cráneo y las longitudes relativas del tronco, espalda y extremidades. Las hienas pardas y rayadas tienen una dieta compuesta mayormente de carroña de animales grandes y cazan animales pequeños, complementando con materia vegetal como frutas, las cuales pueden componer más de la mitad de la dieta de algunos individuos de la hiena parda en algunos momentos de año.[36] Otro modelo de comparación sería el lobo de crin de América del Sur. Este cánido omnívoro habita en zonas de planicies abiertas, tiene extremidades extremadamente largas y delgadas en comparación con su tamaño corporal (como también se ha interpretado a veces a Arctodus simus), no es particularmente rápido, ni caza presas veloces, y corre a zancadas. Las largas extremidades pueden ser una adaptación para incrementar el campo visual por sobre la vegetación alta de los entornos abiertos. Sin embargo, es igualmente posible que las largas extremidades de Arctodus simus fueran usadas para romper y empujar la vegetación, incluyendo arbustos y pequeños árboles, con el fin de alimentarse de las hojas, frutas y cortezas.[43]
Herbívoro
[editar]El que Arctodus no se diferenciara significativamente en dentadura o constitución de los osos modernos a llevado a algunos autores a apoyar la hipótesis de que A. simus y el oso de las cavernas eran omnívoros como muchos de sus parientes actuales, y el primero habría comido plantas dependiendo de la disponibilidad.[58] Un estudio de 2006 encontró que había adaptaciones dentales y craneanas para comer plantas en Arctodus simus, sugiriendo que la dieta de la especie incluía una gran cantidad de materia vegetal. Sus adaptaciones craneanas para el incremento de la fuerza de mordida incluyendo el rostro corto, el hocico ancho (el cual habría evitado que se ramoneara de forma selectiva, y la ausencia de adaptaciones para cavar en sus extremidades delanteras (lo que habría limitado la búsqueda de raíces) sugiere que los vegetales de su dieta eran principalmente follaje grueso, el cual se recolectaba de forma poco selectiva.[36] Además, el tamaño corporal de Arctodus simus excede las limitaciones esperadas para un carnívoro terrestre del Cuaternario (tomando en cuenta la base energética más restringida para una dieta carnívora). Un estudio de 1985 sugiere que esta discrepancia en el tamaño, junto con una dentadura similar a la del oso de anteojos, indica una dieta principalmente herbívora, pero con el potencial para ser un carnívoro oportunista.[43] No obstante, esto ha sido discutido, específicamente basándose en que el cráneo y las proporciones de Arctodus serían un impedimento para forrajear (especialmente en áreas abiertas), y la abundancia de grandes presas contemporáneas.[52] En todo caso, morfológicamente Arctodus simus muestra características comunes con los osos herbívoros. Estas incluyen dientes posteriores con grandes superficies de masticación, una mandíbula alta, y grandes zonas de anclaje muscular. Dado que las especies de tendencias herbívoras del orden Carnivora carecen de un tracto digestivo que pueda procesar de forma eficiente el material vegetal a través de la acción microbiana, ellos deben desmenuzar los alimentos vegetales usando una larga masticación para molerla, por lo que poseen rasgos que permiten una mayor ventaja mecánica de la mandíbula.[56]
En Beringia
[editar]Los análisis de huesos de ejemplares de Alaska mostraron altas concentraciones de nitrógeno-15, un isótopo estable de nitrógeno que es acumulado por los carnívoros, sin evidencia de ingestión de vegetación. Con base en esta evidencia, se sugirió que A. simus habría sido un hipercarnívoro (con porcentajes inusualmente altos de consumo de caribúes) y que un adulto habría requerido 16 kg de carne al día para sobrevivir.[30][59][60] Adicionalmente, Arctodus parece haber excluido tanto a los leones cavernarios como los osos pardos del este de Beringia en algunas épocas durante el Pleistoceno tardío.[21] Los estudios señalan que el análisis de nitrógeno-15 no puede distinguir entre los hipercarnívoros y los omnívoros que consumen una cantidad significativa de comida de origen animal,[36] que la especie habría tenido una dieta a lo largo de su rango,[56] y que los rasgos de su cráneo y dientes encajan con los de los osos omnívoros modernos. Sería razonable asumir que la carne y médulas serían probablemente los principales recursos alimenticios para algunas de las poblaciones boreales de A. simus, para las cuales la supervivencia durante las temporadas de invierno dependerían del consumo regular de cadáveres de ungulados, como ocurre con los osos de Kodiak.[1] Sin embargo, la extinción local de Arctodus en Beringia hace unos 20 mil años (tras haber llegado hace unos 50 mil años),[21][42] mucho antes que en otras zonas de su rango, hace preguntarse que tan equipado estaba Arctodus en un hipotético nicho de carnívoro, y por qué, si los osos pardos y leones de las cavernas y otros animales lograron recolonizar Beringia una vez que el corredor libre de hielo se reabrió hacia el sur al final del Pleistoceno, Arctodus no invadió de nuevo la zona.[61]
Interacción con humanos
[editar]Se ha documentado una posible interacción de estos osos con gente de la cultura Clovis en Lubbock Lake Landmark, Texas. Un ejemplar posiblemente ya muerto de Arctodus simus habría sido procesado para uso de subsistencia (las marcas de corte indican que los huesos fueron desarticulados, y se retiró la piel y carne) y de tecnología (el material restante sería usado para producción de herramientas), de la misma forma en que se usó un cuerpo de mamut colombino de ese sitio (hace 11,100 años).[24] Adicionalmente, se han encontrado otros restos de Arctodus simus en asociación con artefactos paleoamericanos en Sheriden Cave, en el condado de Wyandot en Ohio.[62]
Alguna vez se pensó que los humanos que se desplazaban dentro de Norteamérica habrían encontrado en los grandes carnívoros del Pleistoceno como Arctodus simus un obstáculo para poder establecerse. Los ejemplares de Beringia de Arctodus eran los mayores y más poderosos mamíferos carnívoros norteamericanos, con la potencial especialización de obtener y dominar recursos alimenticios distantes. En esta hipótesis, los humanos aunque estaban familiarizados con los osos pardos, no habrían podido contender efectivamente contra Arctodus simus y otros grandes carnívoros de la época, una situación que hubiera podido frenar la expansión de la población humana. Sin embargo, con los hallazgos de ocupación humana anteriores al Último Máximo Glacial a lo largo del continente americano (incluyendo la cueva Chiquihuite, con una fecha de ocupación humana de hace 26 mil años), indican que hubo un periodo extenso de coexistencia con Arctodus y pone en duda esa idea.[42] Aunque los humanos estaban débilmente dispersos en Norteamérica durante el final del Pleistoceno, existe evidencia de tecnologías líticas diversas y posiblemente aisladas entre sí (como los Clovis, Tradición de canto del oeste y Cola de pescado) pero no así de que estas mismas poblaciones humanas fueran obstaculizadas por Arctodus u otros mamíferos del Pleistoceno al sur de la capa de hielo Laurentino, especialmente considerando que las dataciones sugieren un ritmo de expansión incluso más rápido a lo largo del continente.
Probablemente los humanos estarían involucrados en ocasiones en la muerte o aprovechamiento de cuerpos de varios de los diferentes megaherbívoros del Pleistoceno, particularmente mamuts y mastodontes. Estas situaciones pueden haber puesto a los pobladores en competencia con Arctodus simus por las presas, seguramente defendiéndose así como abandonado los restos. En todo caso, la relación entre los humanos y Arctodus simus habría sido cuando mucho, incómoda.[42]

Extinción
[editar]La datación más reciente para Arctodus simus es 9,630 ± 60 años antes del presente, sin embargo dado que la vértebra en cuestión proveniente de Bonner Springs, Kansas, fue datada por otro laboratorio en 10,921 ± 50 antes del presente con colágeno bien preservado, la fecha más reciente es vista con cautela. Un rango de edad para la vértebra puede ser estimado al usar las dataciones para el yacimiento de Bonner Springs (9,510–11,021 años antes del presente, en 2σ). Dos otros especímenes de la Cueva Friesenhahn en Texas y Huntington Dam en Utah) fueron datados igualmente en cerca de 10,814 años antes del presente (± 55 antes del presente).[29] La zona central y sur de las Montañas Rocosas puede haber actuado como un refugio para Arctodus simus, así como para otra fauna contemporánea de zonas altas como Mammuthus colombi, Mammut americanum, Equus conversidens y Bison latifrons hace ≤10,000 años antes del presente.[63]
Se piensa que Arctodus simus se habría extinto hace unos 10,000 años, lo cual es relativamente tardío al compararse con otras especies de megafauna que desaparecieron en el evento de extinción del Cuaternario. Se han sugerido varios factores para la desaparición de la especie, incluyendo la disminución en el número del número de los grandes herbívoros, la reducción de la calidad nutritiva de las plantas durante el cambio climático, y la competencia con otros grandes omnívoros (humanos y osos pardos) por los recursos alimenticios.[24] Sin embargo, varios estudios ha cuestionado que los osos pardos sean responsables de la extinción de Arctodus simus.[14][42][52] Adicionalmente, no hay evidencia fuerte de que los humanos cazaran a los grandes carnívoros del Pleistoceno en Norteamérica, y no hay indicación clara de una intervención humana directa en su desaparición.[42] Tampoco se conoce en la evidencia recuperada en Rancho La Brea algo que sugiera que habían periodos de hambruna que pudieran ser responsables del fin de Arctodus simus, o de otros carnívoros de gran tamaño.[56]
De todos estos factores, los cambios en la vegetación al final del Pleistoceno pueden haber sido particularmente desfavorables para Arctodus simus, debido a la reducción de alimentos con la calidad mínima para la subsistencia. Por ejemplo, en la isla de Vancouver (∼11.7 años antes del presente), la vegetación cambió rápidamente de bosques abiertos con abundantes pinos lodgepole a bosques cada vez más cerrados con árboles tolerantes a la sombra como píceas, cicutas de montaña y alisos rojos. Estos cambios, que se volvieron extensos hace ∼10.5 miles de años, apuntan hacia las condiciones más frías y húmedas durante el estadio del Dryas Reciente. Los bosques cerrados continuaron expandiéndose en el inicio del Holoceno, volviéndose dominante la tsuga del Pacífico. Incluso aunque Arctodus simus no estaba restringido a las áreas abiertas y podía vivir en diferentes ambientes, la sincronización del cambio regional de un hábitat de bosques de pinos abiertos a una vegetación boscosa densa implica que este cambio de vegetación contribuyó a la extirpación local de Arctodus simus, junto con muchos otros miembros de la megafauna.[42]
Especies
[editar]Véase también
[editar]Referencias
[editar]- ↑ a b c d e f g h i j Figueirido (2010). «Demythologizing Arctodus simus, the 'short-faced' long-legged and predaceous bear that never was». Journal of Vertebrate Paleontology 30 (1): 262-275. S2CID 85649497. doi:10.1080/02724630903416027.
- ↑ a b Christiansen, Per (1999). «What size were Arctodus simus and Ursus spelaeus (Carnivora: Ursidae)?». Annales Zoologici Fennici 36 (2): 93-102. ISSN 0003-455X.
- ↑ Cope E. D. (1879). «The cave bear of California». American Naturalist 13: 791.
- ↑ Feranec, Robert S. (2009). «Implications of Radiocarbon Dates from Potter Creek Cave, Shasta County, California, USA». Radiocarbon (en inglés) 51 (3): 931-936. ISSN 0033-8222. doi:10.1017/S0033822200034007.
- ↑ Richards, Ronald L.; Neiburger, Ellis J.; Turnbull, William D. (1995). Giant short-faced bear (Arctodus simus yukonensis) remains from Fulton County, northern Indiana. Chicago, Ill.: Field Museum of Natural History.
- ↑ Stark, Mike (2022). Chasing the Ghost Bear: On the Trail of America’s Lost Super Beast (en inglés). U of Nebraska Press. pp. Fig. 19. ISBN 978-1-4962-2902-1.
- ↑ a b c d Emsliet, Steven D. (2016). «The fossil record of Arctodus pristinus (Ursidae, Tremarctinae) in Florida». www.semanticscholar.org (en inglés). Consultado el 26 de febrero de 2022.
- ↑ a b Soibelzon, Leopoldo H.; Romero, M.R. Aguilar (14 de octubre de 2008). «A Blancan (Pliocene) short-faced bear from El Salvador and its implications for Tremarctines in South America». Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen 250 (1): 1-8. ISSN 0077-7749. doi:10.1127/0077-7749/2008/0250-0001.
- ↑ a b c d e f g h i j k l Schubert, Blaine; Hulbert, Richard; MacFadden, Bruce; Searle, Michael; Searle, Seina (1 de enero de 2010). «Giant Short-faced Bears (Arctodus simus) in Pleistocene Florida USA, a Substantial Range Extension». Journal of Paleontology - J PALEONTOL 84: 79-87. doi:10.1666/09-113.1.
- ↑ Dalquest, W. W.; Mooser, O. (19 de diciembre de 1980). «Arctodus pristinus Leidy in the Pleistocene of Aguascalientes, Mexico». Journal of Mammalogy 61 (4): 724-725. ISSN 1545-1542. doi:10.2307/1380320.
- ↑ a b I. Ferrusquia-Villafranca. 1978. Bol Univ Nac Aut Mex Inst Geol 101:193-321
- ↑ Feranec, Robert S.; Hadly, Elizabeth A.; Blois, Jessica L.; Barnosky, Anthony D.; Paytan, Adina (2007). «Radiocarbon Dates from the Pleistocene Fossil Deposits of Samwel Cave, Shasta County, California, USA». Radiocarbon (en inglés) 49 (1): 117-121. ISSN 0033-8222. doi:10.1017/S0033822200041941.
- ↑ a b Sanders, Albert E. (2002). Additions to the Pleistocene Mammal Faunas of South Carolina, North Carolina, and Georgia (en inglés). American Philosophical Society. ISBN 978-0-87169-925-1.
- ↑ a b c d e Esker, Donald; Wilkins, William; Agenbroad, Larry (13 de agosto de 2010). A multivariate analysis of the ecology of North American Pleistocene bears, with a focus on Arctodus simus.
- ↑ a b Russell, Dale A.; Rich, Fredrick J.; Schneider, Vincent; Lynch-Stieglitz, Jean (May 2009). «A warm thermal enclave in the Late Pleistocene of the South-eastern United States». Biological Reviews (en inglés) 84 (2): 173-202. doi:10.1111/j.1469-185X.2008.00069.x.
- ↑ C. S. Churcher, A. V. Morgan, and L. D. Carter. 1993. Arctodus simus from the Alaskan Arctic Slope. Canadian Journal of Earth Sciences 30(5):1007-1013, collected by A. V. Morgan
- ↑ Cassiliano M. L. (1999). «Biostratigraphy of Blancan and Irvingtonian mammals in the Fish Creek-Vallecito Creek section, southern California, and a review of the Blancan-Irvingtonian boundary». Journal of Vertebrate Paleontology 19 (1): 169-186. doi:10.1080/02724634.1999.10011131.
- ↑ Carranza-Castañeda, Oscar; Miller, Wade E. (16 de septiembre de 1987). «Rediscovered type specimens and other important published Pleistocene mammalian fossils from Central Mexico». Journal of Vertebrate Paleontology 7 (3): 335-341. ISSN 0272-4634. doi:10.1080/02724634.1987.10011664.
- ↑ Richards, Ronald L.; Churcher, C. S.; Turnbull, William D. (18 de noviembre de 2019). Distribution and size variation in North American Short-faced bears, Arctodus simus (en inglés). University of Toronto Press. ISBN 978-1-4875-7415-4. doi:10.3138/9781487574154-012.
- ↑ Brown, Gary (1996). Great Bear Almanac. p. 340. ISBN 978-1558214743. (requiere registro).
- ↑ a b c d e Salis, Alexander T.; Bray, Sarah C. E.; Lee, Michael S. Y.; Heiniger, Holly; Barnett, Ross; Burns, James A.; Doronichev, Vladimir; Fedje, Daryl; Golovanova, Liubov; Harington, C. Richard; Hockett, Bryan (3 de septiembre de 2020). Lions and brown bears colonized North America in multiple synchronous waves of dispersal across the Bering Land Bridge (en inglés). pp. 2020.09.03.279117. doi:10.1101/2020.09.03.279117v1.full.
- ↑ Schubert, Blaine W.; Chatters, James C.; Arroyo-Cabrales, Joaquin; Samuels, Joshua X.; Soibelzon, Leopoldo H.; Prevosti, Francisco J.; Widga, Christopher; Nava, Alberto; Rissolo, Dominique; Erreguerena, Pilar Luna (May 2019). «Yucatán carnivorans shed light on the Great American Biotic Interchange». Biology Letters 15 (5): 20190148. ISSN 1744-9561. PMC 6548739. PMID 31039726. doi:10.1098/rsbl.2019.0148.
- ↑ Arroyo-Cabrales, Joaquin; Johnson, Eileen; Graham, Ruswell; perez crespo, Victor (24 de julio de 2016). «North American ursid (Mammalian: Ursidae) defaunation from Pleistocene to recent.». Cranium 33: 51-56.
- ↑ a b c Pérez-Crespo, J. Arroyo-Cabrales E. Johnson R.W. Graham V.A. (1 de enero de 2016). North American ursid (Mammalia: Ursidae) defaunation from Pleistocene to recent. OCLC 1227719621.
- ↑ a b Pedersen, Mikkel Winther; De Sanctis, Bianca; Saremi, Nedda F.; Sikora, Martin; Puckett, Emily E.; Gu, Zhenquan; Moon, Katherine L.; Kapp, Joshua D.; Vinner, Lasse; Vardanyan, Zaruhi; Ardelean, Ciprian F. (21 de junio de 2021). «Environmental genomics of Late Pleistocene black bears and giant short-faced bears». Current Biology (en inglés) 31 (12): 2728-2736.e8. ISSN 0960-9822. doi:10.1016/j.cub.2021.04.027.
- ↑ Nelson, Michael E.; Madsen, James H. (1983). «A Giant Short-Faced Bear (Arctodus simus) from the Pleistocene of Northern Utah». Transactions of the Kansas Academy of Science (1903-) 86 (1): 1-9. ISSN 0022-8443. doi:10.2307/3628418.
- ↑ a b c Chubert, Blaine; Kaufmann, James (1 de agosto de 2003). «A partial short-faced bear skeleton from an Ozark Cave with comments on the paleobiology of the species». Journal of Cave and Karst Studies 65.
- ↑ a b Fowler, Nicholas L.; Spady, Thomas J.; Wang, Guiming; Leopold, Bruce D.; Belant, Jerrold L. (October 2021). «Denning, metabolic suppression, and the realisation of ecological opportunities in Ursidae». Mammal Review (en inglés) 51 (4): 465-481. ISSN 0305-1838. doi:10.1111/mam.12246.
- ↑ a b c d Schubert, Blaine W. (15 de abril de 2010). «Late Quaternary chronology and extinction of North American giant short-faced bears (Arctodus simus)». Quaternary International. Faunal Dynamics and Extinction in the Quaternary: Studies in Honor of Ernest L. Lundelius, Jr. (en inglés) 217 (1): 188-194. ISSN 1040-6182. doi:10.1016/j.quaint.2009.11.010.
- ↑ a b c d e Nancy Sisinyak. «The Biggest Bear ... Ever». Alaska Fish and Wildlife News. Consultado el 12 de enero de 2008.
- ↑ a b Soibelzon, Leopoldo H.; Schubert, Blaine W. (2011). «The largest known bear, Arctotherium angustidens, from the Early Pleistocene Pampean region of Argentina: with a discussion of size and diet trends in bears». Journal of Paleontology 85 (1): 69-75. ISSN 0022-3360.
- ↑ «Bear Claw Marks». Riverbluff Cave - The Official Website. Consultado el 17 de abril de 2014.
- ↑ «Cave Animals». Riverbluff Cave - The Official Website. Consultado el 17 de abril de 2014.
- ↑ Mitchell, Kieren J.; Bray, Sarah C.; Bover, Pere; Soibelzon, Leopoldo; Schubert, Blaine W.; Prevosti, Francisco; Prieto, Alfredo; Martin, Fabiana; Austin, Jeremy J.; Cooper, Alan (30 de abril de 2016). «Ancient mitochondrial DNA reveals convergent evolution of giant short-faced bears (Tremarctinae) in North and South America». Biology Letters 12 (4): 20160062. PMC 4881349. PMID 27095265. doi:10.1098/rsbl.2016.0062.
- ↑ a b Meloro, Carlo; de Oliveira, Alessandro Marques (1 de marzo de 2019). «Elbow Joint Geometry in Bears (Ursidae, Carnivora): a Tool to Infer Paleobiology and Functional Adaptations of Quaternary Fossils». Journal of Mammalian Evolution (en inglés) 26 (1): 133-146. ISSN 1573-7055. doi:10.1007/s10914-017-9413-x.
- ↑ a b c d e f g h i Sorkin, B. (2006). «Ecomorphology of the giant short-faced bears Agriotherium and Arctodus». Historical Biology 18 (1): 1-20. S2CID 85301983. doi:10.1080/08912960500476366.
- ↑ «Arctodus pristinus». Florida Museum (en inglés estadounidense). 30 de marzo de 2017. Consultado el 21 de febrero de 2022.
- ↑ Gould, G.C.; Quitmyer, Irvy (1 de enero de 2005). «Titanis walleri: Bones of contention». Bulletin of the Florida Museum of Natural History 45: 201-229.
- ↑ MacFadden, Bruce; Labs-Hochstein, Joann; Hulbert, Richard; Baskin, Jon (1 de febrero de 2007). «Revised age of the late Neogene terror bird (Titanis) in North America during the Great American Interchange». Geology 35. doi:10.1130/G23186A.1.
- ↑ Nowak, Ronald M. (1999). Walker's mammals of the world (Sixth edición). Baltimore: Johns Hopkins University Press. ISBN 0-8018-5789-9. OCLC 39045218.
- ↑ Figueirido, Borja; Soibelzon, Leopoldo H. (19 de agosto de 2009). «Inferring palaeoecology in extinct tremarctine bears (Carnivora, Ursidae) using geometric morphometrics». Lethaia 43 (2): 209-222. ISSN 0024-1164. doi:10.1111/j.1502-3931.2009.00184.x.
- ↑ a b c d e f g h i j k Steffen, Martina L.; Fulton, Tara L. (1 de febrero de 2018). «On the association of giant short-faced bear (Arctodus simus) and brown bear (Ursus arctos) in late Pleistocene North America». Geobios (en inglés) 51 (1): 61-74. ISSN 0016-6995. doi:10.1016/j.geobios.2017.12.001.
- ↑ a b c d e f Emslie, Steven D.; Czaplewski, Nicholas J. (15 de noviembre de 1985). «A new record of giant short-faced bear, Arctodus simus, from western North America with a re-evaluation of its paleobiology». Contributions in science 371: 1-12. ISSN 0459-8113. doi:10.5962/p.226835.
- ↑ a b c Pérez-Crespo, Víctor Adrián; Arroyo-Cabrales, Joaquín; Morales-Puente, Pedro; Cienfuegos-Alvarado, Edith; Otero, Francisco J. (March 2018). «Diet and habitat of mesomammals and megamammals from Cedral, San Luis Potosí, México». Geological Magazine (en inglés) 155 (3): 674-684. ISSN 0016-7568. doi:10.1017/S0016756816000935.
- ↑ a b c Figueirido, Borja; Perez, Alejandro; Schubert, Blaine; Serrano, Francisco; Farrell, Aisling; Pastor, Francisco; Neves, Aline; Romero, Alejandro (19 de diciembre de 2017). «Dental caries in the fossil record: A window to the evolution of dietary plasticity in an extinct bear». Scientific Reports 7. doi:10.1038/s41598-017-18116-0.
- ↑ Harris, Arthur (30 de agosto de 2014). Pleistocene Vertebrates of Southwestern USA and Northwestern Mexico.
- ↑ Harris, Arthur H. «Reconstruction of Mid Wisconsin Environments in Southern New Mexico». NATIONAL GEOGRAPHIC RESEARCH.
- ↑ Trayler, Robin B.; Dundas, Robert G.; Fox-Dobbs, Kena; Van De Water, Peter K. (13 de noviembre de 2012). «Inland California during the Pleistocene—Megafaunal stable isotope records reveal new paleoecological and paleoenvironmental insights». Palaeogeography, Palaeoclimatology, Palaeoecology 437: 132-140. ISSN 0031-0182. doi:10.1016/j.palaeo.2015.07.034.
- ↑ Mychajliw, Alexis M.; Rick, Torben C.; Dagtas, Nihan D.; Erlandson, Jon M.; Culleton, Brendan J.; Kennett, Douglas J.; Buckley, Michael; Hofman, Courtney A. (16 de septiembre de 2020). «Biogeographic problem-solving reveals the Late Pleistocene translocation of a short-faced bear to the California Channel Islands». Scientific Reports (en inglés) 10 (1): 15172. ISSN 2045-2322. doi:10.1038/s41598-020-71572-z.
- ↑ Soibelzon, Leopoldo H.; Grinspan, Gustavo A.; Bocherens, Hervé; Acosta, Walter G.; Jones, Washington; Blanco, Ernesto R.; Prevosti, Francisco (November 2014). «South American giant short-faced bear (Arctotherium angustidens) diet: evidence from pathology, morphology, stable isotopes, and biomechanics». Journal of Paleontology (en inglés) 88 (6): 1240-1250. ISSN 0022-3360. doi:10.1666/13-143.
- ↑ Czaplewski, Nicholas; Rogers, Kyler; Russell, Clayton (1 de junio de 2018). «Late Pleistocene vertebrates from three-forks cave, Adair county, Oklahoma Ozark highland». Journal of Cave and Karst Studies 80 (2): 1-16. doi:10.4311/2017PA0118.
- ↑ a b c Mattson, David J. (1998). «Diet and Morphology of Extant and Recently Extinct Northern Bears». Ursus 10: 479-496. JSTOR 3873160.
- ↑ a b c d e E., Matheus, Paul (2003). Locomotor adaptations and ecomorphology of short-faced bears (Arctodus simus) in eastern Beringia. Yukon Palaeontologist, Gov't. of Yukon. OCLC 243520303.
- ↑ a b Randally, Lynch, Eric (6 de agosto de 2012). Cursorial Adaptations in the Forelimb of the Giant Short-Faced Bear, Arctodus simus, Revealed by Traditional and 3D Landmark Morphometrics. East Tennessee State University. OCLC 818344518.
- ↑ «The Giant Short-Faced Bear». North American Bear Center. 2 de marzo de 2018. Consultado el 1 de marzo de 2022.
- ↑ a b c d e Donohue, Shelly L.; DeSantis, Larisa R. G.; Schubert, Blaine W.; Ungar, Peter S. (2013). «Was the giant short-faced bear a hyper-scavenger? A new approach to the dietary study of ursids using dental microwear textures». PLOS ONE 8 (10): e77531. Bibcode:2013PLoSO...877531D. PMC 3813673. PMID 24204860. doi:10.1371/journal.pone.0077531.
- ↑ a b DeSantis, Larisa; Schubert, Blaine; Schmitt-Linville, Elizabeth; Ungar, Peter; Donohue, Shelly; Haupt, Ryan (1 de enero de 2015). «Dental Microwear Textures of Carnivorans from the La Brea Tar Pits, California, and Potential Extinction Implications». La Brea and Beyond: The Paleontology of Asphalt-Preserved Biotas: 37-52.
- ↑ ScienceDaily, 13 April 2009.«Prehistoric bears ate everything and anything, just like modern cousins». ScienceDaily. Consultado el 13 de abril de 2009.
- ↑ Bocherens, H.; Emslie, S. D.; Billiou, D.; Mariotti A. (1995). «Stable isotopes (13C, 15N) and paleodiet of the giant short-faced bear (Arctodus simus)». C R Acad Sci 320: 779-784.
- ↑ National Geographic Channel, 16 September 2007, Prehistoric Predators: Short-faced bear,’’ interview with Dr. Paul Matheus
- ↑ Pedersen, Mikkel W.; Ruter, Anthony; Schweger, Charles; Friebe, Harvey; Staff, Richard A.; Kjeldsen, Kristian K.; Mendoza, Marie L. Z.; Beaudoin, Alwynne B.; Zutter, Cynthia; Larsen, Nicolaj K.; Potter, Ben A. (2016). «Postglacial viability and colonization in North America's ice-free corridor». Nature 537 (7618): 45-49. Bibcode:2016Natur.537...45P. PMID 27509852. S2CID 4450936. doi:10.1038/nature19085.
- ↑ Brian G. Redmond, PhD., Curator of Archaeology (March 2006). «Before the Western Reserve: An Archaeological History of Northeast Ohio». The Cleveland Museum of Natural History. p. 2. Consultado el 28 de enero de 2020.
- ↑ Gillette, David D.; Madsen, David B. (6 de marzo de 1992). «The short-faced bear Arctodus simus from the late Quaternary in the Wasatch Mountains of central Utah». Journal of Vertebrate Paleontology 12 (1): 107-112. doi:10.1080/02724634.1992.10011436.
Enlaces externos
[editar]Wikimedia Commons alberga una galería multimedia sobre Arctodus.
Wikispecies tiene un artículo sobre Arctodus.
- Arctodus simus descripción e ilustración (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última).