History of genetics
| Part of a series on |
| Genetics |
|---|
 |
The history of genetics dates from the classical era with contributions by Pythagoras, Hippocrates, Aristotle, Epicurus, and others. Modern genetics began with the work of the Augustinian friar Gregor Johann Mendel. His works on pea plants, published in 1866, provided the initial evidence that, on its rediscovery in 1900's, helped to establish the theory of Mendelian inheritance.
In ancient Greece, Hippocrates suggested that all organs of the body of a parent gave off invisible “seeds,” miniaturised components, that were transmitted during sexual intercourse and combined in the mother's womb to form a baby. In the Early Modern times, William Harvey's book On Animal Generation contradicted Aristotle's theories of genetics and embryology.
The 1900 rediscovery of Mendel's work by Hugo de Vries, Carl Correns and Erich von Tschermak led to rapid advances in genetics. By 1915 the basic principles of Mendelian genetics had been studied in a wide variety of organisms — most notably the fruit fly Drosophila melanogaster. Led by Thomas Hunt Morgan and his fellow "drosophilists", geneticists developed the Mendelian model, which was widely accepted by 1925. Alongside experimental work, mathematicians developed the statistical framework of population genetics, bringing genetic explanations into the study of evolution.
With the basic patterns of genetic inheritance established, many biologists turned to investigations of the physical nature of the gene. In the 1940s and early 1950s, experiments pointed to DNA as the portion of chromosomes (and perhaps other nucleoproteins) that held genes. A focus on new model organisms such as viruses and bacteria, along with the discovery of the double helical structure of DNA in 1953, marked the transition to the era of molecular genetics.
In the following years, chemists developed techniques for sequencing both nucleic acids and proteins, while many others worked out the relationship between these two forms of biological molecules and discovered the genetic code. The regulation of gene expression became a central issue in the 1960s; by the 1970s gene expression could be controlled and manipulated through genetic engineering. In the last decades of the 20th century, many biologists focused on large-scale genetics projects, such as sequencing entire genomes.
Pre-Mendel ideas on heredity
[edit]Ancient theories
[edit]
The most influential early theories of heredity were that of Hippocrates and Aristotle. Hippocrates' theory (possibly based on the teachings of Anaxagoras) was similar to Darwin's later ideas on pangenesis, involving heredity material that collects from throughout the body. Aristotle suggested instead that the (nonphysical) form-giving principle of an organism was transmitted through semen (which he considered to be a purified form of blood) and the mother's menstrual blood, which interacted in the womb to direct an organism's early development.[1] For both Hippocrates and Aristotle—and nearly all Western scholars through to the late 19th century—the inheritance of acquired characters was a supposedly well-established fact that any adequate theory of heredity had to explain. At the same time, individual species were taken to have a fixed essence; such inherited changes were merely superficial.[2] The Athenian philosopher Epicurus observed families and proposed the contribution of both males and females of hereditary characters ("sperm atoms"), noticed dominant and recessive types of inheritance and described segregation and independent assortment of "sperm atoms".[3]
In 1000 CE, the Arab physician, Abu al-Qasim al-Zahrawi (known as Albucasis in the West) was the first physician to describe clearly the hereditary nature of haemophilia in his Al-Tasrif.[4] In 1140 CE, Judah HaLevi described dominant and recessive genetic traits in The Kuzari.[5]
Preformation theory
[edit]
The preformation theory is a developmental biological theory, which was represented in antiquity by the Greek philosopher Anaxagoras. It reappeared in modern times in the 17th century and then prevailed until the 19th century. Another common term at that time was the theory of evolution, although "evolution" (in the sense of development as a pure growth process) had a completely different meaning than today. The preformists assumed that the entire organism was preformed in the sperm (animalkulism) or in the egg (ovism or ovulism) and only had to unfold and grow. This was contrasted by the theory of epigenesis, according to which the structures and organs of an organism only develop in the course of individual development (Ontogeny). Epigenesis had been the dominant opinion since antiquity and into the 17th century, but was then replaced by preformist ideas. Since the 19th century epigenesis was again able to establish itself as a view valid to this day.[6][7]
Plant systematics and hybridisation
[edit]In the 18th century, with increased knowledge of plant and animal diversity and the accompanying increased focus on taxonomy, new ideas about heredity began to appear. Linnaeus and others (among them Joseph Gottlieb Kölreuter, Carl Friedrich von Gärtner, and Charles Naudin) conducted extensive experiments with hybridisation, especially hybrids between species. Species hybridisers described a wide variety of inheritance phenomena, include hybrid sterility and the high variability of back-crosses.[8]
Plant breeders were also developing an array of stable varieties in many important plant species. In the early 19th century, Augustin Sageret established the concept of dominance, recognising that when some plant varieties are crossed, certain characteristics (present in one parent) usually appear in the offspring; he also found that some ancestral characteristics found in neither parent may appear in offspring. However, plant breeders made little attempt to establish a theoretical foundation for their work or to share their knowledge with current work of physiology,[9] although Gartons Agricultural Plant Breeders in England explained their system.[10]
Mendel
[edit]
Between 1856 and 1865, Gregor Mendel conducted breeding experiments using the pea plant Pisum sativum and traced the inheritance patterns of certain traits. Through these experiments, Mendel saw that the genotypes and phenotypes of the progeny were predictable and that some traits were dominant over others.[11] These patterns of Mendelian inheritance demonstrated the usefulness of applying statistics to inheritance. They also contradicted 19th-century theories of blending inheritance, showing, rather, that genes remain discrete through multiple generations of hybridisation.[12]
From his statistical analysis, Mendel defined a concept that he described as a character (which in his mind holds also for "determinant of that character"). In only one sentence of his historical paper, he used the term "factors" to designate the "material creating" the character: " So far as experience goes, we find it in every case confirmed that constant progeny can only be formed when the egg cells and the fertilising pollen are off like the character so that both are provided with the material for creating quite similar individuals, as is the case with the normal fertilisation of pure species. We must, therefore, regard it as certain that exactly similar factors must be at work also in the production of the constant forms in the hybrid plants."(Mendel, 1866).

Mendel's work was published in 1866 as "Versuche über Pflanzen-Hybriden" (Experiments on Plant Hybridisation) in the Verhandlungen des Naturforschenden Vereins zu Brünn (Proceedings of the Natural History Society of Brünn), following two lectures he gave on the work in early 1865.[13]
Post-Mendel, pre-rediscovery
[edit]Pangenesis
[edit]
Mendel's work was published in a relatively obscure scientific journal, and it was not given any attention in the scientific community. Instead, discussions about modes of heredity were galvanised by Darwin's theory of evolution by natural selection, in which mechanisms of non-Lamarckian heredity seemed to be required. Darwin's own theory of heredity, pangenesis, did not meet with any large degree of acceptance.[14][15] A more mathematical version of pangenesis, one which dropped much of Darwin's Lamarckian holdovers, was developed as the "biometrical" school of heredity by Darwin's cousin, Francis Galton.[16]
Germ plasm
[edit]
In 1883 August Weismann conducted experiments involving breeding mice whose tails had been surgically removed. His results — that surgically removing a mouse's tail had no effect on the tail of its offspring — challenged the theories of pangenesis and Lamarckism, which held that changes to an organism during its lifetime could be inherited by its descendants. Weismann proposed the germ plasm theory of inheritance, which held that hereditary information was carried only in sperm and egg cells.[17]
Rediscovery of Mendel
[edit]Hugo de Vries wondered what the nature of germ plasm might be, and in particular he wondered whether or not germ plasm was mixed like paint or whether the information was carried in discrete packets that remained unbroken. In the 1890s he was conducting breeding experiments with a variety of plant species and in 1897 he published a paper on his results that stated that each inherited trait was governed by two discrete particles of information, one from each parent, and that these particles were passed along intact to the next generation. In 1900 he was preparing another paper on his further results when he was shown a copy of Mendel's 1866 paper by a friend who thought it might be relevant to de Vries's work. He went ahead and published his 1900 paper without mentioning Mendel's priority. Later that same year another botanist, Carl Correns, who had been conducting hybridisation experiments with maize and peas, was searching the literature for related experiments prior to publishing his own results when he came across Mendel's paper, which had results similar to his own. Correns accused de Vries of appropriating terminology from Mendel's paper without crediting him or recognising his priority. At the same time another botanist, Erich von Tschermak was experimenting with pea breeding and producing results like Mendel's. He too discovered Mendel's paper while searching the literature for relevant work. In a subsequent paper de Vries praised Mendel and acknowledged that he had only extended his earlier work.[17]
Emergence of molecular genetics
[edit]After the rediscovery of Mendel's work there was a feud between William Bateson and Pearson over the hereditary mechanism, solved by Ronald Fisher in his work "The Correlation Between Relatives on the Supposition of Mendelian Inheritance".
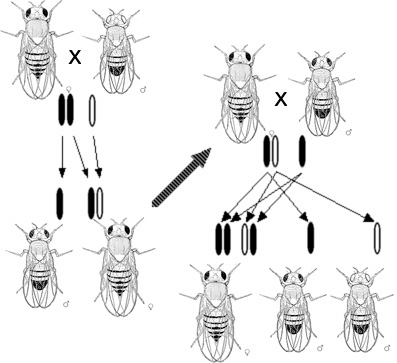
In 1910, Thomas Hunt Morgan showed that genes reside on specific chromosomes. He later showed that genes occupy specific locations on the chromosome. With this knowledge, Alfred Sturtevant, a member of Morgan's famous fly room, using Drosophila melanogaster, provided the first chromosomal map of any biological organism. In 1928, Frederick Griffith showed that genes could be transferred. In what is now known as Griffith's experiment, injections into a mouse of a deadly strain of bacteria that had been heat-killed transferred genetic information to a safe strain of the same bacteria, killing the mouse.
A series of subsequent discoveries (e.g.[18]) led to the realization decades later that the genetic material is made of DNA (deoxyribonucleic acid) and not, as was widely believed until then, of proteins. In 1941, George Wells Beadle and Edward Lawrie Tatum showed that mutations in genes caused errors in specific steps of metabolic pathways.[19] This showed that specific genes code for specific proteins, leading to the "one gene, one enzyme" hypothesis.[20] Oswald Avery, Colin Munro MacLeod, and Maclyn McCarty showed in 1944 that DNA holds the gene's information.[21] In 1952, Rosalind Franklin and Raymond Gosling produced a strikingly clear x-ray diffraction pattern indicating a helical form. Using these x-rays and information already known about the chemistry of DNA, James D. Watson and Francis Crick demonstrated the molecular structure of DNA in 1953.[22] [23] Together, these discoveries established the central dogma of molecular biology, which states that proteins are translated from RNA which is transcribed by DNA. This dogma has since been shown to have exceptions, such as reverse transcription in retroviruses.
In 1947, Salvador Luria discovered the reactivation of irradiated phage[24] leading to many further studies on the fundamental processes of repair of DNA damage (for review of early studies, see [25]). In 1958, Meselson and Stahl demonstrated that DNA replicates semiconservatively, leading to the understanding that each of the individual strands in double-stranded DNA serves as a template for new strand synthesis.[26] In 1960, Jacob and collaborators discovered the operon which consists of a sequence of genes whose expression is coordinated by operator DNA.[27] In the period 1961 – 1967, through work in several different labs, the nature of the genetic code was determined (e.g. [28]).
In 1972, Walter Fiers and his team at the University of Ghent were the first to determine the sequence of a gene: the gene for bacteriophage MS2 coat protein.[29] Richard J. Roberts and Phillip Sharp discovered in 1977 that genes can be split into segments. This led to the idea that one gene can make several proteins. The successful sequencing of many organisms' genomes has complicated the molecular definition of the gene. In particular, genes do not always sit side by side on DNA like discrete beads. Instead, regions of the DNA producing distinct proteins may overlap, so that the idea emerges that "genes are one long continuum".[30][31] It was first hypothesised in 1986 by Walter Gilbert that neither DNA nor protein would be required in such a primitive system as that of a very early stage of the earth if RNA could serve both as a catalyst and as genetic information storage processor.
The modern study of genetics at the level of DNA is known as molecular genetics, and the synthesis of molecular genetics with traditional Darwinian evolution is known as the modern evolutionary synthesis.
See also
[edit]- List of sequenced eukaryotic genomes
- History of molecular biology
- History of RNA Biology
- History of evolutionary thought
- One gene-one enzyme hypothesis
- Phage group
References
[edit]- ^ a b Leroi, Armand Marie (2010). "Function and Constraint in Aristotle and Evolutionary Theory". In Föllinger, S. (ed.). Was ist 'Leben'? Aristoteles' Anschauungen zur Entstehung und Funktionsweise von Leben. Franz Steiner Verlag. pp. 261–284. ISBN 978-3-515-09244-9. OCLC 276334688.
- ^ Mayr, Ernst (1982). The growth of biological thought : diversity, evolution, and inheritance. Belknap Press. pp. 635–640. ISBN 0674364457. OCLC 7875904.
- ^ Yapijakis, C. (2017). "Ancestral Concepts of Human Genetics and Molecular Medicine in Epicurean Philosophy". In Petermann, H.; Harper, P.; Doetz, S. (eds.). History of Human Genetics. Springer. pp. 41–57. doi:10.1007/978-3-319-51783-4_3. ISBN 978-3-319-51783-4.
- ^ Cosman, Madeleine Pelner; Jones, Linda Gale (2008). Handbook to life in the medieval world. Infobase Publishing. pp. 528–529. ISBN 978-0-8160-4887-8.
- ^ HaLevi, Judah, translated and annotated by N. Daniel Korobkin. The Kuzari: In Defense of the Despised Faith, p. 38, I:95: "This phenomenon is common in genetics as well—often we find a son who does not resemble his father at all, but closely resembles his grandfather. Undoubtedly, the genetics and resemblance were dormant within the father even though they were not outwardly apparent. Hebrew by Ibn Tibon, p.375: ונראה כזה בענין הטבעי, כי כמה יש מבני האדם שאינו דומה לאב כלל אך הוא דומה לאבי אביו ואין ספק כי הטבע ההוא והדמיון ההוא היה צפון באב ואף על פי שלא נראה להרגשה
- ^ Jacob, François (1972). Die Logik des Lebenden. Von der Urzeugung zum genetischen Code. Frankfurt am Main: Fischer. ISBN 3-10-035601-2. OCLC 1200295370.
- ^ Jahn, Ilse; Löther, Rolf; Senglaub, Konrad (1985). Geschichte der Biologie: Theorien, Methoden, Institutionen, Kurzbiographien (2nd ed.). Jena: VEB Fischer. ISBN 3437203290. OCLC 19780133.
- ^ Mayr 1982, pp. 640–9
- ^ Mayr 1982, pp. 649–651
- ^ For example, Explanatory Notes, Gartons Seed Catalogue for Spring 1901
- ^ Pierce, Benjamin A. (2020). Genetics A Conceptual Approach (7th ed.). W.H. Freeman. pp. 49–56. ISBN 978-1-319-29714-5.
- ^ Mukherjee, Siddartha (2016). "Chapter 4". The Gene: An intimate history. Scribner. ISBN 978-1-4767-3353-1. OCLC 949803912.
- ^ Alfred, Randy (8 February 2010). "Feb. 8, 1865: Mendel Reads Paper Founding Genetics". Wired. ISSN 1059-1028. Retrieved 11 November 2019.
- ^ Darwin, C.R. (27 April 1871). "Pangenesis". Nature. 3 (78): 502–3. Bibcode:1871Natur...3..502D. doi:10.1038/003502a0.
- ^ Geison, G. L. (1969). "Darwin and heredity: The evolution of his hypothesis of pangenesis". J Hist Med Allied Sci. XXIV (4): 375–411. doi:10.1093/jhmas/XXIV.4.375. PMID 4908353.
- ^ Bulmer, M. G. (2003). Francis Galton: Pioneer of Heredity and Biometry. Johns Hopkins University Press. pp. 116–8. ISBN 978-0-801-88140-4.
- ^ a b Mukherjee 2016, Chapter 5
- ^ Hershey AD, Chase M (May 1952). "Independent functions of viral protein and nucleic acid in growth of bacteriophage". J Gen Physiol. 36 (1): 39–56. doi:10.1085/jgp.36.1.39. PMC 2147348. PMID 12981234.
- ^ Beadle GW, Tatum EL (November 1941). "Genetic Control of Biochemical Reactions in Neurospora". Proc Natl Acad Sci U S A. 27 (11): 499–506. Bibcode:1941PNAS...27..499B. doi:10.1073/pnas.27.11.499. PMC 1078370. PMID 16588492.
- ^ Gerstein MB, Bruce C, Rozowsky JS, Zheng D, Du J, Korbel JO, Emanuelsson O, Zhang ZD, Weissman S, Snyder M (June 2007). "What is a gene, post-ENCODE? History and updated definition". Genome Research. 17 (6): 669–681. doi:10.1101/gr.6339607. PMID 17567988.
- ^ Steinman RM, Moberg CL (February 1994). "A triple tribute to the experiment that transformed biology". The Journal of Experimental Medicine. 179 (2): 379–84. doi:10.1084/jem.179.2.379. PMC 2191359. PMID 8294854.
- ^ Pierce, Benjamin A. (2020). Genetics A Conceptual Approach (7th ed.). New York: W.H. Freeman. pp. 299–300. ISBN 978-1-319-29714-5.
- ^ Watson JD, Crick FH (25 April 1953). "Molecular structure of nucleic acids; a structure for deoxyribose nucleic acid". Nature. 171 (4356): 737–8. Bibcode:1953Natur.171..737W. doi:10.1038/171737a0. PMID 13054692.
- ^ Luria SE (September 1947). "Reactivation of Irradiated Bacteriophage by Transfer of Self-Reproducing Units". Proc Natl Acad Sci U S A. 33 (9): 253–264. Bibcode:1947PNAS...33..253L. doi:10.1073/pnas.33.9.253. PMC 1079044. PMID 16588748.
- ^ Bernstein C (March 1981). "Deoxyribonucleic acid repair in bacteriophage". Microbiol Rev. 45 (1): 72–98. doi:10.1128/mr.45.1.72-98.1981. PMC 281499. PMID 6261109.
- ^ Meselson M, Stahl FW (July 1958). "The replication of DNA in Escherichia Coli". Proc Natl Acad Sci U S A. 44 (7): 671–682. Bibcode:1958PNAS...44..671M. doi:10.1073/pnas.44.7.671. PMC 528642. PMID 16590258.
- ^ Jacob F, Perrin D, Sánchez C, Monod J, Edelstein S (2005) [1960]. "The operon: a group of genes with expression coordinated by an operator". C R Biol. 328 (6): 514–520. doi:10.1016/j.crvi.2005.04.005. PMID 15999435.
- ^ Crick FH, Barnett L, Brenner S, Watts-Tobin RJ (December 1961). "General nature of the genetic code for proteins". Nature. 192 (4809): 1227–32. Bibcode:1961Natur.192.1227C. doi:10.1038/1921227a0. PMID 13882203.
- ^ Min Jou W, Haegeman G, Ysebaert M, Fiers W (May 1972). "Nucleotide sequence of the gene coding for the bacteriophage MS2 coat protein". Nature. 237 (5350): 82–8. Bibcode:1972Natur.237...82J. doi:10.1038/237082a0. PMID 4555447. S2CID 4153893.
- ^ Pearson, H. (May 2006). "Genetics: what is a gene?". Nature. 441 (7092): 398–401. Bibcode:2006Natur.441..398P. doi:10.1038/441398a. PMID 16724031. S2CID 4420674.
- ^ Pennisi E (June 2007). "Genomics. DNA study forces rethink of what it means to be a gene". Science. 316 (5831): 1556–7. doi:10.1126/science.316.5831.1556. PMID 17569836. S2CID 36463252.
Further reading
[edit]- Elof Axel Carlson, Mendel's Legacy: The Origin of Classical Genetics (Cold Spring Harbor Laboratory Press, 2004.) ISBN 0-87969-675-3
External links
[edit]This section's use of external links may not follow Wikipedia's policies or guidelines. (January 2023) |
- Olby's "Mendel, Mendelism, and Genetics," at MendelWeb
- Andrei, A. (2013). ""Experiments in Plant Hybridization" (1866), by Johann Gregor Mendel". Embryo Project Encyclopedia. Arizona State University.
- http://www.accessexcellence.org/AE/AEPC/WWC/1994/geneticstln.html
- http://www.sysbioeng.com/index/cta94-11s.jpg
- http://www.esp.org/books/sturt/history/
- http://cogweb.ucla.edu/ep/DNA_history.html
- "The history of genetics". BBC. 2000.
- https://web.archive.org/web/20120323085256/http://www.hchs.hunter.cuny.edu/wiki/index.php?title=Modern_Science&printable=yes
- Avery, Oswald T.; MacLeod, Colin M.; McCarty, Maclyn (February 1944). "Studies on the chemical nature of the substance inducing transformation of Pneumococcal types: Induction of transformation by a DEsoxyribonucleic acid fraction isolated from Pneumococcus Type III ". J Exp Med. 79 (2): 137–158. doi:10.1084/jem.79.2.137. PMC 2135445. PMID 19871359.
- http://www.nature.com/physics/looking-back/crick/Crick_Watson.pdf
- Todd, AR (1954). "Chemical Structure of the Nucleic Acids". Proc. Natl. Acad. Sci. U.S.A. 40 (8): 748–55. Bibcode:1954PNAS...40..748T. doi:10.1073/pnas.40.8.748. PMC 534157. PMID 16589553.
- http://www.genomenewsnetwork.org/resources/timeline/1960_mRNA.php
- https://web.archive.org/web/20120403041525/http://www.molecularstation.com/molecular-biology-images/data/503/MRNA-structure.png
- http://www.genomenewsnetwork.org/resources/timeline/1973_Boyer.php
- Struhl, Kevin (1 October 2008). "The hisB463 Mutation and Expression of a Eukaryotic Protein in Escherichia coli". Genetics. 180 (2): 709–714. doi:10.1534/genetics.104.96693. PMC 2567374. PMID 18927256.
- Sanger, F; Nicklen, S; Coulson, AR (December 1977). "DNA sequencing with chain-terminating inhibitors". Proc. Natl. Acad. Sci. U.S.A. 74 (12): 5463–7. Bibcode:1977PNAS...74.5463S. doi:10.1073/pnas.74.12.5463. PMC 431765. PMID 271968.
- Jeffreys, AJ; Wilson, V; Thein, SL (1985). "Individual-specific 'fingerprints' of human DNA". Nature. 316 (6023): 76–79. Bibcode:1985Natur.316...76J. doi:10.1038/316076a0. PMID 2989708. S2CID 4229883.
- Cech, T. R.; Bass, B. L. (1986). "Biological Catalysis by RNA". Annual Review of Biochemistry. 55: 599–629. doi:10.1146/annurev.bi.55.070186.003123. PMID 2427016.
- "A sheep cloning how-to, more or less". CNN. February 1997.
- "Human Genome Project" (Fact Sheet). National Human Genome Research Institute.
| Fields, disciplines | |
|---|---|
| Theories, concepts | |
| Related | |
| Histories of basic sciences |  | |
|---|---|---|
| Histories of medical specialties | ||
| Medicine in ancient societies | ||
| History of methods in medicine | ||
| Disasters and plagues | ||