Sensory Systems/Other Animals
Rodents: Somatosensory Perception of Whiskers
[edit | edit source]Introduction
[edit | edit source]

The barrel Cortex is a specialized region in somatosensory cortex responsible for processing the tactile information from whiskers. As every other cortical region, the barrel cortex also preserves the columnar organization which plays a crucial role in information processing. Information from each whisker is represented in separate, discrete columns analogous to “barrels”, hence the name barrel cortex. Rodents use whiskers constantly to acquire sensory information from the environment. Given their nocturnal nature, tactile information carried by whisker forms the primary sensory signals to build a perceptual map of the environment. The whiskers on the snouts of mice and rats serve as arrays of highly sensitive detectors for acquiring tactile information as shown in Figure 1 A and B. By using their whiskers, rodents can build spatial representations of their environment, locate objects, and perform fine-grain texture discrimination. Somatosensory whisker-related processing is highly organized into stereotypical maps, which occupy a large portion of the rodent brain. During exploration and palpation of objects, the whiskers are under motor control, often executing rapid large-amplitude rhythmic sweeping movements, and this sensory system is therefore an attractive model for investigating active sensory processing and sensory-motor integration. In these animals, a large part of the neocortex is dedicated to the processing of information from the whiskers. Since rodents are nocturnal, visual information is relatively poor and they rely heavily on the tactile information from whiskers. Perhaps the most remarkable specialization of this sensory system is the primary somatosensory ‘‘barrel’’ cortex, where each whisker is represented by a discrete and well-defined structure in layer 4.
These layer 4 barrels are somatotopically arranged in an almost identical fashion to the layout of the whiskers on the snout i.e. bordering whiskers are represented in adjacent cortical areas [1]. Sensorimotor integration of whisker related activity leads to pattern discrimination and enables rodents to have a reliable map of the environment. This is an interesting model to study because rodents use whisker to “see” and this cross modality sensory information processing could help us to improve the life of humans, who are deprived of one sensory modality. Specifically, blind people can be trained to use somatosensory information to build a spatial map of the environment [2].
Pathways carrying whisker information to Barrel Cortex
[edit | edit source]|
Pathways carrying whisker information to Barrel Cortex


The sensory neurons make excitatory glutamatergic synapses in the trigeminal nuclei of the brain stem. Trigemino-thalamic neurons in the principal trigeminal nucleus are organized into somatotopically arranged ‘‘barrelettes,’’ each receiving strong input from a single whisker as shown in (Figure 3). The principal trigeminal neurons project to the ventral posterior medial (VPM) nucleus of the thalamus, which is also somatotopically laid out into anatomical units termed ‘‘barreloids’’ VPM neurons respond rapidly and precisely to whisker deflection, with one ‘‘principal’’ whisker evoking stronger responses than all others. The axons of VPM neurons within individual barreloids project to the primary somatosensory neocortex forming discrete clusters in layer 4, which form the basis of the ‘‘barrel’’ map as shown in Figure 3.
|
Whisker information processing in Barrel Cortex with specialized local microcircuit
[edit | edit source]The deflection of a whisker is thought to open mechano-gated ion channels in nerve endings of sensory neurons innervating the hair follicle (although the molecular signalling machinery remains to be identified). The resulting depolarization evokes action potential firing in the sensory neurons of the infraorbital branch of the trigeminal nerve. The transduction through mechanical deformation is similar to the hair cells in the inner ear; in this case the contact of whiskers with the objects causes the mechano-gated ion channels to open. Cation-permeable ion channels let positively charged ions into the cells and causes depolarization, eventually leading to generation of action potentials. A single sensory neuron only fires action potentials to deflection of one specific whisker. The innervation of the hair follicle shows a diversity of nerve endings, which may be specialized for detecting different types of sensory input [3].
The layer 4 barrel map is arranged almost identically to the layout of the whiskers on the snout of the rodent. There are several recurrent connections in layer 4 and it sends axons to layer 2/3 neurons, which integrates information from other cortical regions like primary motor cortex. These intra-cortical and inter-cortical connections enable the rodents to achieve stimulus discrimination capabilities and to extract optimal information from the incoming tactile stimulus. Also, these projections play a crucial role in integrating somatosensory information with motor output. Information from whiskers is processed in the barrel cortex with specialized local microcircuits formed to extract optimal information about the environment. These cortical microcircuits are composed of excitatory and inhibitory neurons as shown in Figure 4.

Learning whisker based object discrimination & texture differentiation
[edit | edit source]Rodents move their sensors to collect information, and these movements are guided by sensory input. When action sequences are required to achieve success in novel tasks, interactions between movement and sensation underlie motor control [4] and complex learned behaviours [5]. The motor cortex has important roles in learning motor skills [6-9], but its function in learning sensorimotor associations is unknown. The neural circuits underlying sensorimotor integration are beginning to be mapped. Different motor cortex layers harbour excitatory neurons with distinct inputs and projections [10-12]. Outputs to motor centres in the brain stem and spinal cord arise from pyramidal tract-type neurons in layer 5B (L5B). Within motor cortex, excitation descends from L2/3 to L5 [13, 14]. Input from somatosensory cortex impinges preferentially onto L2/3 neurons. L2/3 neurons [10] therefore directly link somatosensation and control of movements. In one of the recent studies [15], mice were trained head fixed in a vibrissa-based object-detection task while imaging populations of neurons [16]. Following a sound, a pole was moved to one of several target positions within reach of the whiskers (the ‘go’ stimulus) or to an out-of-reach position (the ‘no-go’ stimulus). Target and out-of-reach locations were arranged along the anterior–posterior axis; the out-of reach position was most anterior. Mice searched for the pole with one whisker row, the C row, and reported the pole as ‘present’ by licking, or ‘not present’ by withholding licking. Licking on go trials (hit) was rewarded with water, whereas licking on no-go trials (false alarm) was punished with a time-out during which the trial was stopped for 2 seconds. Trials without licking (no-go, correct rejection, go, and miss) were not rewarded or punished. All mice showed learning within the first two or three sessions. Performance reached expert levels after three to six training sessions. Learning the behavioural task was directly dependent on the motor related behaviour. Naive mice whisked occasionally in a manner unrelated to trail structure. Thus, object detection relies on a sequence of actions, linked by sensory cues. An auditory cue triggers whisking during the sampling period. Contact between whisker and object causes licking for a water reward during a response period. Silencing vM1 indicates that this task requires the motor cortex; with vM1 silenced, task-dependent whisking persisted, but was reduced in amplitude and repeatability, and task performance dropped.
Neural Correlates of Sensorimotor learning mechanism
[edit | edit source]Coding of touch in the motor cortex is consistent with direct input from vS1 to the imaged neurons. A model based on population coding of individual behavioural features also predicted motor behaviours. Accurate decoding of whisking amplitude, whisking set-point and lick rate suggests that vM1 controls these slowly varying motor parameters, as expected from previous motor cortex and neurophysiological experiments.
References
[edit | edit source]1 Feldmeyer D, Brecht M, Helmchen F, Petersen CCH, Poulet JFA, Staiger JF, Luhmann HJ, Schwarz C."Barrel cortex function" Progress in Neurobiology 2013, 103 : 3-27.
2 Lahav O, Mioduser D. "Multisensory virtual environment for supporting blind persons' acquisition of spatial cognitive mapping, orientation, and mobility skills." Academia.edu 2002.
3 Alloway KD. "Information processing streams in rodent barrel cortex: The differential functions of barrel and septal circuits." Cereb Cortex 2008, 18(5):979-989.
4 Scott SH. "Inconvenient truths about neural processing in primary motor cortex." The Journal of physiology 2008, 586(5):1217-1224.
5 Wolpert DM, Diedrichsen J, Flanagan JR. "Principles of sensorimotor learning." Nature reviews Neuroscience 2011, 12(12):739-751.
6 Wise SP, Moody SL, Blomstrom KJ, Mitz AR. "Changes in motor cortical activity during visuomotor adaptation." Experimental brain research Experimentelle Hirnforschung Experimentation cerebrale 1998, 121(3):285-299.
7 Rokni U, Richardson AG, Bizzi E, Seung HS. "Motor learning with unstable neural representations." Neuron 2007, 54(4):653-666.
8 Komiyama T, Sato TR, O'Connor DH, Zhang YX, Huber D, Hooks BM, Gabitto M, Svoboda K. "Learning-related fine-scale specificity imaged in motor cortex circuits of behaving mice." Nature 2010, 464(7292):1182-1186.
9 Hosp JA, Pekanovic A, Rioult-Pedotti MS, Luft AR. "Dopaminergic projections from midbrain to primary motor cortex mediate motor skill learning." The Journal of neuroscience : the official journal of the Society for Neuroscience 2011, 31(7):2481-2487.
10 Keller A. "Intrinsic synaptic organization of the motor cortex." Cereb Cortex 1993, 3(5):430-441.
11 Mao T, Kusefoglu D, Hooks BM, Huber D, Petreanu L, Svoboda K. "Long-range neuronal circuits underlying the interaction between sensory and motor cortex." Neuron 2011, 72(1):111-123.
12 Hooks BM, Hires SA, Zhang YX, Huber D, Petreanu L, Svoboda K, Shepherd GM. "Laminar analysis of excitatory local circuits in vibrissal motor and sensory cortical areas." PLoS biology 2011, 9(1):e1000572.
13 Anderson CT, Sheets PL, Kiritani T, Shepherd GM. "Sublayer-specific microcircuits of corticospinal and corticostriatal neurons in motor cortex." Nature neuroscience 2010, 13(6):739-744.
14 Kaneko T, Cho R, Li Y, Nomura S, Mizuno N. "Predominant information transfer from layer III pyramidal neurons to corticospinal neurons." The Journal of comparative neurology 2000, 423(1):52-65.
15 O'Connor DH, Clack NG, Huber D, Komiyama T, Myers EW, Svoboda K. "Vibrissa-based object localization in head-fixed mice." The Journal of neuroscience : the official journal of the Society for Neuroscience 2010, 30(5):1947-1967.
16 O'Connor DH, Peron SP, Huber D, Svoboda K. "Neural activity in barrel cortex underlying vibrissa-based object localization in mice." Neuron 2010, 67(6):1048-1061.
17 Shaner NC, Campbell RE, Steinbach PA, Giepmans BN, Palmer AE, Tsien RY. "Improved monomeric red, orange and yellow fluorescent proteins derived from Discosoma sp. red fluorescent protein." Nature biotechnology 2004, 22(12):1567-1572.
18 Tian L, Hires SA, Mao T, Huber D, Chiappe ME, Chalasani SH, Petreanu L, Akerboom J, McKinney SA, Schreiter ER. "Imaging neural activity in worms, flies and mice with improved GCaMP calcium indicators." Nature methods 2009, 6(12):875-881.
Snakes: Sensing of Infrared Radiation
[edit | edit source]Introduction
[edit | edit source]
When seeing or sometimes even thinking of snakes, many people feel uncomfortable or even scared. There is a reason that they are considered being mythical. Snakes are different when compared to other animals: they do not have legs, they are long and move elegantly and without a noise, some of them are venomous and they steadily use their forked tongue to smell. Some of them are fast and effective killers even by night. Something that definitely makes them special is their „sixth sense“: the ability to detect infrared radiation. Similar to night viewers, snakes are capable of detecting heat changes in their surroundings and thus obtaining a detailed picture of it. There are at least two different groups of snakes which have separately developed this ability: in the first are the pit vipers, and in the second boas and pythons (those two are often classified into one group called “boids”). However, snakes are not the only species which have evolved this sense: vampire bats and some groups of insects have also developed it. Even at night pit vipers, boas and pythons can make out rodents due to the heat they emit. It can be detected by a sensory system that allows them to „see“ electromagnetic radiation with long wavelengths ranging from 750 nm to 1 mm. The organs which make that possible are called “pit organs”, and are located under their eyes, inside two hollows of the maxilla bone. They are immensely sensitive as they can even detect changes in temperature of as little as 0.003K.
Anatomy of Sensing Organ
[edit | edit source]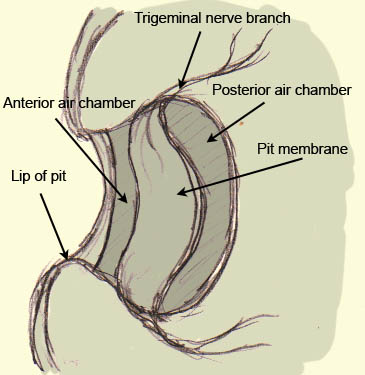
The infrared-sensing organs of vipers and boids are similar in their physiological structure but differ in their number, location and morphology. The anatomy is quite simple and will be explained in the example of the crotalus, a venomous pit viper found only in the Americas from southern Canada to northern Argentina. It consists of a hollow space that is separated into two air-filled chambers by a thin membrane of the thickness of 0.01 mm. It is filled with sensory cells of the trigeminal nerve (TNM). Roughly 7000 in number, they transduce the heat through heat sensitive ion channels, and increase their firing rate when a positive change in temperature occurs and decrease in the opposite case. They are very sensitive due to the spatial proximity of these thermoreceptors to the outside and also because of the air-filled chamber that lies underneath. This air-filled chamber works as an insulator in separating tissues that would otherwise quickly exchange heat energy. Thus, the absorbed thermal energy is used exclusively by the sensory system and is not lost to lower-lying tissues. This simple but sophisticated anatomy is the reason for the unique sensitivity of the pit organs. The pit organs’s physique allows even the detection of the radiation’s direction. The external opening is roughly half as large as the membrane. Thus, the whole organ works according to the optics of a pinhole camera: the position of the irradiated spot provides information about the object’s location. The heat itself is detected by the activation of heat sensitive ion channels called TRPA1. In other animals these channels also exist but have other functions like detecting chemical irritants or cold. Pit vipers and boids seem to have evolved the infrared-sensing independently. Since the heat sensitive ion channels have different thermal thresholds in different snakes, the temperature sensitivity differs among the snakes. Crotalus have the most sensitive channels. Snakes that are not able to detect infrared radiation also possess those channels, but their thermal threshold is too high to detect infrared radiation.
Brain’s Anatomy
[edit | edit source]Every sensory organ has a dedicated brain region to process the collected information. Snakes evaluate infrared sensory input from the pit organs in the nucleus of the lateral descending trigeminal tract (“LTTD”), a unique region in their metencephalon which has not been found in other animals. The LTTD is linked to the tectum opticum via the reticularis caloris (“RC”). Its function is still unknown. In the tectum opticum visual and infrared stimuli are connected, in order to provide a detailed idea of the animal’s surrounding.
Physiology
[edit | edit source]Experiments have shown that the detection of heat targets must be quite accurate as snakes hit thermal sources with a low error even without the help of vision. Measurements have determined that the opening angle of an infrared beam falling onto the pit organ is 45 to 60 degrees. Depending on where the heat source is relatively to the snake, the beam hits the pit’s membrane on a different spot. The receptive field of the infrared sensing system on the tectum opticum is similarly represented as the visual receptive field. The front-end of the tectum opticum receives its input from the back part of the pit membrane and the retina, and thus processes stimuli from the front part of the visual field. Similarly, the back and the sides of the visual field are represented in the back part of the tectum opticum and the front part of the pit membrane and the retina. The receptive fields of the visual and infrared sensory systems overlap almost perfectly within the tectum opticum, such that the neurons there receive and process sensory information from two senses, from more or less the same direction. While crotalus only have two pit organs, the anatomy of the temperature sensors is much more complicated in boas and pythons. They possess 13 pit organs on each side of the head. Every one of those also works like a pinhole camera that reverses the picture. The information of the front part of the visual field is again processed in the front part of the tectum opticum but now, the receptive field of every pit organ is projected onto a different part of it. The front pit organs are represented in the front part of the tectum opticum and the back parts in the back. In addition, the receptive fields of the different pit organs overlap, and thus provide a more or less continuous projective field that matches the visual one. It is curious that the front part of every pit organ is projected to the back part of the receptive field in the tectum opticum, an organization that is quite complicated and unique. The tectum opticum contains six different kinds of neurons which fire for infrared and/or visual stimuli. Some cell types respond only if there is a visual and an infrared stimulus, while others respond for any kind of stimulus. There are cells that respond for one of the sensory input if it comes alone, but increases its firing rate for simultaneous input from both systems. The last group of cells works the other way around. Some of them respond strongly for visual stimuli and stop firing when stimuli from the pit organs also arrive or vice versa. What do snakes with pit organs need these different kinds of neurons for? The processing in their brain has to help the snakes with different tasks: first of all, the snake should be able to detect and locate stimuli. Second, they have to be identified and reacted to appropriately. The cells that respond to both visual and infrared stimuli independently from each other could be responsible for the first task. Cells that only respond if they get both stimuli at the same time could work as detectors for living, moving objects. Moreover, cells that stop firing as soon as the visual stimuli is completed with an infrared signal could be especially important for detecting the cool surrounding like leaves or trees. The interaction between the different types of cells are important for correctly identifying the stimuli. They are not only used for identifying warm-blooded prey, but also for identifying predators and the snake’s thermoregulation.

Infrared Sensing in Vampire Bats
[edit | edit source]Vampire bats are the only mammals that are able to detect infrared radiation. To do so they have three hollows in their nose which contain the sensing organs. While they also use ion channels to detect heat, it is a different type of ion channels than in snakes. In other mammals and even everywhere in their own body except for the nose this type of molecule is responsible for sensing pain and burning. However, in the nose the threshold is much lower. The channel already detects changes in temperature from 29 °C on. This allows vampire bats to locate heat sources at a distance of 20 cm and helps them to find blood-rich spots on their prey.
References
[edit | edit source]Newman, E.A., and Hartline, P.H. (1982) Infrared "vision" in snakes. Scientific American 246(3):116-127 (March).
Gracheva et al.: Molecular Basis of Infrared Detection by Snakes. Nature. 2010 April 15; 464(7291): 1006-1011
Campbell et. Al.: Biological infrared Imaging and sensing. Micron 33 (2002) 211-225
Gracheva et al.: Ganglion-specific splicing of TRPV1 underlies infrared sensation in vampire bats. Nature.476, 88-91 (04.08.2011)
The Sensory System of Bats
[edit | edit source]Introduction
[edit | edit source]The order of bats (chiroptera), which is made up by the suborders of microbats (microchiroptera) and megabats (megachiroptera), are the only mammals that possess the ability to actively fly.[1][2] This aerial lifestyle, coupled with the fact that bats are mostly active during the night, brings with it the necessity for a sensory system that is very different from that of mammals living and walking on land. It is important that bats can spot objects that are in their way to not fly into them. Additionally, bats must be able to find the prey they hunt and predators that could become dangerous to them in the dark. This means that while most mammals rely mostly on their vision for movement and navigation, bats do not rely only on their eyes to scan their surroundings. Many bats navigate relying heavily on their auditory system, as they use echolocation, or biosonar, to detect objects.[3] But these bats do not only rely on their auditory sense. In fact, they understand their environment through a combination of echolocation and vision, as well as other senses.[4]

This combination of echolocation and vision for movements is not used in all bats, as all megabats (with the exception of Rousettus) and some microbats do not possess the ability to echolocate, but rely on vision and other senses, as well as their memory for navigation and object recognition.[2][5]
Animals use echolocation by producing sounds and interpreting the returning echoes that occur when the sound waves hit objects.[3][6] Most animals use echolocation to locate other animals, such as prey or predators. Additionally, animals can detect objects in their surroundings with the help of echolocation and move around these objects. [5]
But echolocation is not only used by bats. Many birds, such as cave swiftlets, also possess the ability to echolocate.[2][3][5] Echolocation is not only useful for animals that can fly, but is also used by many animals living in the sea and by some nocturnal land mammals, such as shrews and rats.[2] Some mammals and fish living in water, for example dolphins and whales, echolocate, as sound travels efficiently and over long distances in water, and relying on vision is often not possible in the darkness and vastness of the ocean.[3][5]
The Auditory System of Bats: Echolocation
[edit | edit source]As explained in the introduction, echolocation is mostly used when vision is not effective, for example in the dark or in water. The animal produces a sound, and then receives and interprets the echoes that return from objects. The outgoing pulse is then compared with the returning echo, and this information is used by the brain to produce an image of the objects in the animal’s surroundings.[3] As there are many different bat species, there are many different echolocation strategies, although all strategies have similar underlying mechanisms.[2][7]
Very good hearing is necessary to be able to process sounds during echolocation. Bats even hear frequencies in the ultrasonic range above 20kHz, which is far beyond the human range. This is also the range in which they emit the sounds used for echolocation. The very high ultrasonic frequencies are reflected not only by big obstacles, but also by small targets such as insects. This makes them very useful for bats when hunting.[3]
A Short History of Echolocation
[edit | edit source]
In 1793, Lazzaro Spallanzani found out that if he took away the sense of vision by blinding bats, they could still move around obstacles. A year later, Charles Jurine observed that if the auditory sense was taken away, bats flew into objects.[3][5] These two experiments led to the believe that bats somehow “see” with their ears instead of their eyes. The term “echolocation” was first used by Donald Griffin in 1938, when he detected the ultrasonic sounds produced by bats via a microphone.[3][5] In the following decades, the principle of echolocation was found to be used in other animals, for example in dolphins.[3][6]
Ear Anatomy
[edit | edit source]The ears of echolocating bats have a similar anatomy to those in most other mammals. Their outer ear has a very large pinna to be able to detect incoming sounds from echolocation, and some bats even use the pinna to actively listen for sounds emitted by predators or prey.[7] The tragus, a flap of skin in the external ear, is used to interpret the direction of an incoming echo during echolocation.[3][7] The ear canal of bats, the tympanic membrane, the three middle ear bones and their cochlea have a similar structure to most mammals, including humans.[7] The cochlea in the bat’s inner ear is covered by sensory cells, which connect to the brain via the auditory nerve. When sound waves enter the cochlea, the sensory cells convert the resulting vibration into neural signals and pass them to the auditory neurons to be transmitted to the brain. A recent study has found the anatomy of the inner ear of bats, namely the wall structure of the ganglion canal around the ganglion neurons, to be connected to their ability to use echolocation and to the frequencies they use for echolocation.[8]
Sound Production and Propagation
[edit | edit source]Bats produce sounds via a continuous emission of high-frequency, ultrasonic sound waves produced in their larynx.[3][5] These sounds are then emitted through the animal’s mouth or nose, depending on the bat species.[7] Many bats can vary their calling rate depending on what they are calling out for, and different species of bats produce signals with different frequencies across the duration of the sound.[1] The produced sound waves are then carried in all directions through the air in the form of sound pulses, as is shown by the orange wave in Figure 3.

Echo Reception and Sound Processing in Ear and Brain
[edit | edit source]If the sound waves released by the bat come into contact with an object, some of the waves are reflected, as is shown by the green wave in Figure 3. The incoming echo is received by the bat via their relatively big ear flaps, the pinnae. When receiving the echo, bats can extract information about the size, distance, shape, and texture of the object.[3]
The time delay between the production of a call and the reception of the returning signal is measured to find the distance to an object.[7] The vertical direction of an object is found via interpretation of vertical angles, which in bats is done using the tragus. The horizontal direction is determined by differences in the sound intensity received in each ear. The size of an object can be determined via the strength of the echo, and the peaks and troughs in the frequency spectrum of the echo give cues about surface texture.[3] Effects such as the Doppler effect, as well as different pitches during sound production, also help bats detect the distance between them and a detected object. The bat’s auditory nervous system processes the information received by the ears and a mental map of the bat’s surroundings is constructed in the bat’s brain.[1] The bat can thus avoid obstacles and predators and locate prey using echolocation.
During their evolution, bats have developed a “send-receive-switch” system. When they produce their very loud calling sounds, the receiver function is disconnected momentarily to make sure it is not damaged by the loud sound, and then connected again to receive the returning signal, which separates the call and echo in time. This switching is done via middle ear muscles attached to the bones of the inner ear. When a bat emits a loud sound, these muscles contract, and the bones cannot transmit sounds well, leading to sounds not being received while the muscles are contracted, which is an active reflex in humans as well.[2][3][5] Most bats only make another call once echoes from the last call have been received, as this makes sure the outgoing and incoming waves do not disrupt each other.[7]
The Role of Echolocation in Hunting
[edit | edit source]As mentioned before, bats continuously emit sound pulses and interpret echoes in real time, to be able to change their flight speed and trajectory very fast to move around objects.[9] But this is not all echolocation is helpful for. While echolocation is mostly used for orientation during movement, it can additionally be useful for detection and classification of prey and predators.[3][6] Some bats can even adjust the frequency of their calls depending on the circumstances, for greater hunting efficiency and adaptability.
When on a routine flight, bats produce about 5 to 10 calls per second, and when prey, such as an insect, is located, the calling rate will increase to find the exact location of the insect, and can reach up to 200 calls per second.[5] A noctule bat hunting insects for example will thus use relatively long narrowband signals to detect prey, and once an insect has been detected, will switch to shorter broadband signals with an increasing calling rate to find the exact localization of the insect, leading to a so-called “feeding buzz”.[3] If an animal or object is still far away, a too high calling rate is not preferable, as outgoing signals can mix with returning signals and lead to wrong interpretation of the object’s location.[5] Additionally, calling also needs a lot of energy, so a lower calling rate is better when no near prey or object has been located.[5]
Other Senses
[edit | edit source]Vision
[edit | edit source]Even though most bats use echolocation to understand their surroundings, they also possess the ability to see. Bats are said to have high spatial acuity, sensitivity and even possess the ability for depth perception. They have the best vision in low-light conditions, as they mostly move in the dark. Their vision is mostly black and white, and they only possess limited colour vision.[4]
Touch
[edit | edit source]While research about the sensory system of bats mostly focuses on the role of echolocation, some studies show that bats have hairs on their flight membranes that act as airflow sensors.[9] This means that bats feel air flow when they fly and can respond to changes in aerodynamic conditions as well as to echoes returning from objects. The hair cells on the wings seem to play a role especially in the adaptation of flight speed.[9][10] Additionally, bats can use the sense of touch for object detection, which can help them capture prey in situations where echolocation is not effective.
Smell and Taste
[edit | edit source]The olfactory and gustatory senses are the least important senses for bats. The sense of taste, like in humans, is mostly active when tasting food. The sense of smell is used for identification of possible mates, as well as prey and predator detection. Additionally, bats make use of their sense of smell to communicate.[4]
Multisensory Integration
[edit | edit source]The combination of vision and echolocation bats use during their flight makes them ideal animals to study multisensory integration.[4] Although this might not be possible for a long time anymore, as bat populations have declined over the last years, leading many bat species to be classified as endangered.[11] Danilovich et al. found bats to use their sense of vision to learn the three-dimensional shapes of objects in their surroundings and when deciding where to fly. Meanwhile, echolocation seems to be more important when approaching an obstacle or classifying an object. They have additionally found bats to be able to translate information received via echolocation into a visual representation in their brain.[4] Not only vision and hearing are important, as the sense of touch seems to play a role in flight navigation, although the importance of the tactile sense in comparison to the visual and auditory sense is debated.[9]
References
[edit | edit source][1] Simmons JA and Stein RA (1980) "Acoustic imaging in bat sonar: Echolocation signals and the evolution of echolocation". Journal of Comparative Physiology. 135: 61-84.
[2] Suga N (2009). "Echolocation II: neurophysiology". Elsevier: 801-802.
[3] Jones G (2005). "Echolocation". Current Biology. 15 (13): 484-488
[4] Danilovich S and Yovel Y (2019). "Integrating vision and echolocation for navigation and perception in bats". Science Advances. 5 (6): eaaw6503.
[5] "Bats: Sensory Systems and Echolocation". science.jrank.org. 2023. Retrieved 20 July 2023.
[6] Au WWL (2018). "Echolocation". Encyclopedia of Marine Mammals. Elsevier: 289-299.
[7] Erbe, Christina; Thomas Jeannette A. (2022). Exploring Animal Behaviour Through Sound: Volume 1 (Methods). Springer AG. p. 419-431.
[8] Sulser RB et al. (2022). "Evolution of inner ear neuroanatomy of bats and implications for echolocation". Nature. 602: 449-454.
[9] Jones G (2011). "Sensory Biology: Bats Feel The Air Flow". Current Biology. 21 (17): 666-667.
[10] Sterbing-D'Angelo S et al. (2011). "Bat wing sensors support flight control". Proceedings of the National Academy of Sciences. 108 (27): 11291-11296.
[11] "Conservation and Biodiversity: Why American bats are declining". earthday.org. 2019. Retrieved 8 August 2023.
- ↑ a b c d Simmons JA and Stein RA (1980). "Acoustic imaging in bat sonar: Echolocation signals and the evolution of echolocation". Journal of Comparative Physiology. 135: 61–84.
- ↑ a b c d e f g Suga N (2009). "Echolocation II: neurophysiology". Elsevier: 801–812.
- ↑ a b c d e f g h i j k l m n o p q Jones G (2005). "Echolocation". Current Biology. 15 (13): 484–488.
- ↑ a b c d e f Danilovich S and Yovel Y (2019). "Integrating vision and echolocation for navigation and perception in bats". Science Advances. 5 (6): eaaw6503.
- ↑ a b c d e f g h i j k l "Bats: Sensory Systems and Echolocation". science.jrank.org. 2023. Retrieved 20 July 2023.
- ↑ a b c d Au WWL (2018). "Echolocation". Encyclopedia of Marine Mammals. Elsevier: 289–299.
- ↑ a b c d e f g h Erbe, Christina; Thomas, Jeannette A. (2022). Exploring Animal Behavior Through Sound: Volume 1 (Methods). Springer AG. p. 419-431.
- ↑ a b Sulser RB and Patterson BD and Urban DJ and Neander AI and Luo Z (2022). "Evolution of inner ear neuroanatomy of bats and implications for echolocation". Nature. 602: 449–454.
- ↑ a b c d e Jones G (2011). "Sensory Biology: Bats Feel The Air Flow". Current Biology. 21 (17): 666–667.
- ↑ a b Sterbing D'Angelo S and Chadha M and Chiu C and Falk B and Xian W and Barcelo J and Zook JM and Moss CF (2011). "Bat wing sensors support flight control". Proceedings of the National Academy of Sciences, 108(27), 11291-11296. 108 (27): 11291–11296.
- ↑ a b "Conservation and Biodiversity: Why American bats are declining". earthday.org. 2019. Retrieved 8 August 2023.