Sensorische Systeme/ Auditorisches System
Einleitung
[Bearbeiten]Das auditorische System (auditive System) ist ein Sinnessystem, das zum Hören (auditive Wahrnehmung) dient. Dieser Wikibook Eintrag beschäftigt sich mit der Physiologie des auditorischen Systems und der erfolgreichsten neurosensorischen Prothese, dem Cochlea Implantat. Die zugrundeliegende Physik und Technik zum Thema Akustik wird in einem separaten Wikibook behandelt, Acoustics. Eine exzellente Quelle für Bilder und Animationen findet man auf der Webseite "Journey into the world of hearing" [1].
Im Reich der Tiere sind Sinne wie fühlen, schmecken und reichen weit mehr verbreitet als die Fähigkeit zu hören. Die Fähigkeit zu hören ist hauptsächlich auf Wirbeltiere und Insekten beschränkt. Innerhalb der Wirbeltiere haben Vögel und Säugetiere den am weitesten Entwickelten Gehörsinn. Die folgende Tabelle zeigt die Frequenzbereiche von Menschen und einigen anderen Tieren:
| Menschen | 20-20'000 Hz |
|---|---|
| Wale | 20-100'000 Hz |
| Fledermäuse | 1'500-100'000 Hz |
| Fische | 20-3'000 Hz |
Das Organ mit welchem wir Schall wahrnehmen ist das Ohr. Es fungiert als Empfänger bei der Erfassung akustischer Informationen und deren Weiterleitung durch das Nervensystem ins Gehirn. Das Ohr enthält sowohl Strukturen für den Gehörsinn, als auch für den Gleichgewichtssinn und ist somit ein wichtiger Bestandteil des Gehörsinnes, des Gleichgewichtssinnes und der Propriozeption.
 |
 |
 |
 |
Menschen haben zwei Ohren, welche symmetrisch auf beiden Seiten des Kopfes angeordnet sind. Das ermöglicht uns, Schallquellen zu lokalisieren. Das Gehirn extrahiert und verarbeitet verschiedene Formen von Daten, um Schall zu lokalisieren. Im Folgenden einige Beispiele:
- Die Form des Schallspektrums an der Membrana timpani (Trommelfell)
- Unterschied in der Lautstärke zwischen den beiden Ohren
- Unterschied in der Ankunftszeit des Schalles zwischen den beiden Ohren
- Art der Verzerrung des Schalles durch die Ohrmuschel. (Die Ohrmuschel wirkt als Schallfänger und Filter. Schallsignale werden von der Ohrmuschel auf unterschiedliche Art und Weise linear verzerrt. Die Art der Verzerrung hängt von der Einfallsrichtung des Schalles und der Entfernung der Schallquelle ab.)
Gesunde, junge Menschen sind in der Lage, Töne in einem Frequenzbereich von 20 Hz bis 20 kHz zu hören. Unser Gehörsinn ist am sensibelsten im Frequenzbereich von gesprochenen Worten, welcher zwischen 2000 und 4000 Herz liegt. Die Frequenzauflösung, welche der Mensch noch wahrnehmen kann, liegt bei 0.2%. Das bedeutet, dass man zwischen einem Ton von 1000 Hz und 1002 Hz unterscheiden kann. Ein Schall von 1 kHz kann erkannt werden, wenn er die Trommelfellmembran um weniger als 1 Angström ablenkt, was kleiner ist als der Durchmesser eines Wasserstoffatoms. Diese extreme Empfindlichkeit des Ohres kann veranschaulichen, warum der kleinste Knochen des menschlichen Körpers, der Steigbügel, im Gehör zu finden ist. Er ist 0,25 bis 0,33 cm lang und wiegt zwischen 1,9 und 4,3 mg.
Anatomie des Auditiven Systems
[Bearbeiten]
Dieser Abschnitt erklärt die Anatomie des menschlichen auditorischen Systems. Das Kapitel erläutert die Bestandteile des Hörorgans in der gleichen Reihenfolge, wie die akustische Information bei der Schallwahrnehmung das Hörorgan passiert und zum Gehirn gelangt. Schallwellen sind Druckänderungen in der Luft. Das auditive System kann diese Druckänderungen erfassen und in ein elektrisches Signal umwandeln. Diese Signale können dann vom Hirn prozessiert, analysiert und interpretiert werden. Erst werden wir uns mit der Struktur und den Bestandteilen des auditiven Systems beschäftigen. Zum Hörsystem gehören Aussen-, Mittel- und Innenohr sowie der Hörnerv und die Umschalt- und Verarbeitungsstationen im zentralen Nervensystem.
Das Ohr
[Bearbeiten]Das Ohr ist das Organ, in dem die erste Schallverarbeitung stattfindet und in dem sich die Sinnesrezeptoren befinden. Es besteht aus drei Teilen:
- Aussenohr
- Mittelohr
- Innenohr

Aussenohr
[Bearbeiten]Funktion: Sammeln von Schallenergie und verstärken des Schalldrucks Der sichtbare Teil des Ohres ist die Ohrmuschel, bestehen aus elastischem Knorpel.
Schallwellen werden reflektiert und gedämpft, wenn sie auf die Ohrmuschel treffen. Diese Veränderungen enthalten für das Gehirn Informationen über die Richtung, aus welcher die Schallwelle gekommen ist. Die Schallwellen wandern weiter in den Gehörgang. Schallwellen, welche zwischen 3 und 12 kHz sind, werden hier amplifiziert. Anschliessend an den Gehörgang findet man das Trommelfell (Membrana tympanica), welches zugleich der Beginn des Mittelohrs ist.
Mittelohr
[Bearbeiten]
Funktion: Das Mittelohr ist eine Schallbrücke, um den hohen Schallwellenwiderstand des Innenohrs zu überwinden. Es überträgt die akustische Energie aus der Luft auf die Cochlea.
Im Mittelohr ist der Hammer in das Trommelfell eingelassen und über den Amboss mit dem Steigbügel verbunden. Die Fussplatte des Steigbügels sitzt beweglich im ovalen Fenster zum Innenohr. Die Schallenergie wird im Mittelohr nicht durch Luftdichtschwankungen, sondern durch Schwingungen des Trommelfells und der Gehörknöchelchen fortgeleitet. Dadurch wird die Reflexion von Schall verringert und mehr Schallenergie kann auf das Innenohr übertragen werden. Somit wird im Mittelohr die Impedanz der Luft an die Impedanz der Flüssigkeit im Innenohr angepasst. Dies geschieht über Hebelwirkung, Druckerhöhung und Herabsetzten der Geschwindigkeit. Der Schall wird nicht gleichmäßig über die gesamte Gehörknöchelkette verstärkt. Der Stapediusreflex der Mittelohrmuskulatur schützt das Innenohr vor Schäden. Das Mittelohr enthält die Schallinformation also noch immer in Wellenform; erst in der Cochlea wird die Information in Nervenimpulse umgewandelt.
Innenohr
[Bearbeiten]| Structural diagram of the cochlea | Cross section of the cochlea |
|---|---|
 |

|
Funktion: Umwandeln der mechanischen Wellen (Schall) in elektrische Signale (Nervensignale).
Das Innenohr besteht aus der Cochlea und mehreren nicht auditiven Strukturen. Die Cochlea ist ein aus mehreren Schläuchen aufgebautes schneckenförmiges Organ. Sie besteht aus drei sogenannten Skalen und dem Corti-Organ. Die Scala tympani (Paukentreppe) und Scala vestibuli (Vorhoftreppe) sind mit Perilymphe gefüllt, eine Flüssigkeit, die sich ähnlich wie andere extrazelluläre Flüssigkeiten zusammensetzt. Unterhalb der Scala vestibuli liegt die Scala media (Schneckengang). Diese wird durch die Reissner-Membran und das Corti-Organ begrenzt. Die Scala media ist mit einer Endolymphe gefüllt, deren Zusammensetzung intrazellulären Flüssigkeiten ähnelt. Die chemischen Unterschiede der Perilymphe und Endolymphe sind wichtig für die Funktion des Innenohres.
Das Corti-Organ
[Bearbeiten]Das Gewebe zwischen Scala media und Scala tympani heisst Corti-Organ. Es enthält die Hörsinneszellen (Haarzellen). Über diese Haarzellen werden die mechanischen Wellen in elektrische Nervensignale umgewandelt. Die Grenzmembran zur Scala tympani heisst Basilarmembran.
Übergang vom Ohr zum Hörnervensystem
[Bearbeiten]
Haarzellen
[Bearbeiten]Haarzellen sind Rezeptoren des Innenohrs, die mechanische Reize in elektrische Aktivität umwandeln. Man unterscheidet zwischen inneren und äusseren Haarzellen. Sie befinden sich beide im Corti-Organ und tragen haarähnliche, submikroskopische Fortsätze, die Stereozilien, welche für die eigentliche Reizaufname zuständig sind. Die inneren Haarzellen sind die "Tonaufnehmer", sie wandeln die Vibration der Schallwellen in elektrische Aktivität um und befinden sich in einer Zellreihe an der Innenseite des Corti-Organs. Die äusseren Haarzellen befinden sich auf der Aussenseite des Corti-Organs. Sie dienen als aktive Verstärker im Innenohr. Über den Stereozilien befindet sich die Tektorialmembran, welche die Spitzen der längsten Stereozilien der äusseren Haarzellen gerade noch berührt. Zwischen Haarzellen und Tektorialmembran befindet sich ein schmaler mit Endolymphe gefüllter Spalt. Tritt der Schall ins Innenohr verursacht er eine Auf- und Abwärtsbewegung von Scala media und Corti-Organ. Diese Auslenkung wiederum führt zu einer Relativbewegung zwischen Corti-Organ und Tektorialmembran. Da die Tektorialmembran die Spitzen der längsten Stereozilien der äusseren Haarzellen berührt, können diese Stereozilien bei dieser Relativbewegung umbiegen und dadurch die Sinneszelle reizen. Die Stereozilien der inneren Haarzellen haben keinen Kontakt zur Tektorailmembran. Sie werden ausgelenkt, in dem der endolymphische Flüssigkeitsfilm zwischen Tektorialmembran und Haarzellen aufgrund der Scherbewegung unter der Tektorialmembran hin- und hergleitet. Die Funktion der Haarzellen ist noch nicht vollständig verstanden. Das Wissen um die Funktion der Haarzellen erlaubt es derzeit, die Zellen bei Hörverlust durch Cochlea-Implantate zu ersetzen. Doch mehr Forschung über die Funktion der Haarzellen könnte es eines Tages sogar ermöglichen, die Zellen zu reparieren. In einem aktuellen Modell sind die Spitzen der Stereozilien über "Tip-Links" miteinander verbunden. Durch Dehnen und Stauchen öffnen die Tip-Links einen Ionenkanal und erzeugen ein Rezeptorpotential in der Haarzelle. Man sollte aber im Kopf behalten, dass eine Ablenkung von 100 Nanometern bereits 90% des vollen Rezeptorpotentials auslöst.
Neuronen
[Bearbeiten]Das Nervensystem unterscheidet zwischen Nervenfasern, die Informationen zum zentralen Nervensystem transportieren, und Nervenfasern, die Informationen vom zentralen Nervensystem wegleiten:
- Afferente Neuronen (auch Sinnes- oder Rezeptorneuronen) tragen Nervenimpulse von Rezeptoren (Sinnesorganen) zum zentralen Nervensystem.
- Efferene Neuronen (auch Motor- oder Effektorneuronen) leiten Nervenimpulse vom zentralen Nervensystem weg zu Effektoren wie Muskeln oder Drüsen (und auch zu den Flimmerzellen des Innenohrs).
Die Ablenkung von Stereozilien der äusseren Haarzellen bewirkt einen Ionenstrom in die Haarzelle und somit eine Potentialänderung der Haarzelle. Durch diese Potentialänderung wird am unteren Ende der Haarzelle der Neurotransmitter Glutamat freigesetzt. Die afferenten Synapsen des Hörnervs befinden sich dort und es entsteht somit ein Aktionspotential. Das Nervenaktionspotential wird über den Hörnerv zum Hirnstamm geführt. Efferente Projektionen vom Gehirn zur Cochlea spielen ebenfalls eine Rolle bei der Wahrnehmung von Klang. Efferente Innervation dient zur selektiven Verstärkung von Frequenzen. Dies soll die Wahrnehmung gewisser Töne und Geräusche zusätzlich verstärken, wenn man sich darauf konzentriert.
Das Auditorische Nervensystem
[Bearbeiten]Die nun in Form von elektrischen Signalen neu codierten Schallinformationen wandern durch den Hörnerv (Hörnerv, vestibulocochlearer Nerv, VIII. Hirnnerv) und passieren verschiedene Zwischenstationen wie die Cochlea-Kerne, den oberen Olivenkernkomplex des Hirnstamms und die Colliculi inferiores des Mittelhirns, an denen sie weiterverarbeitet werden. Die Information erreicht schließlich den Thalamus und wird von dort an den Kortex weitergeleitet. Im menschlichen Gehirn befindet sich der primäre auditive Cortex im Temporallappen.
Primärer Auditiver Cortex
[Bearbeiten]Der primäre auditeve Cortex ist die erste Region der Großhirnrinde, die akustische Inputs erhält.
Die Schallwahrnehmung ist mit dem rechten posterior superior Temporalen-Gyrus (STG) verbunden. Der superiore temporale Gyrus enthält mehrere wichtige Strukturen des Gehirns, einschließlich der Brodmann-Areale 41 und 42. Diese markieren die Region des primären auditiven Kortex, die für die Wahrnehmung grundlegender Klangeigenschaften wie Tonhöhe und Rhythmus verantwortlich ist. Der auditive Assoziationsbereich befindet sich innerhalb des Temporallappens, in einem Bereich, der als Wernicke-Areal oder Areal 22 bezeichnet wird. Dieses Areal, in der Nähe des lateralen Hirnsulkus, ist wichtig für die Verarbeitung akustischer Signale, so dass sie als Sprache, Musik oder Lärm unterschieden werden können.
Auditorische Signalverarbeitung
[Bearbeiten]Nachdem wir die Anatomie des Gehörs angeschaut haben, werden wir uns nun mit den physiologischen Prozessen beschäftigen, welche bei der Wahrnehmung akustischer Informationen und deren Verarbeitung involviert sind. Das Hören beginnt mit Druckwellen, die auf den Gehörgang treffen und endet mit der akustischen Wahrnehmung im Gehirn. Dieser Abschnitt beschreibt den Prozess der Umwandlung von Schwingungen in Wahrnehmung.
Der Effekt des Kopfes
[Bearbeiten]Schallwellen mit einer Wellenlänge kürzer als der Kopf erzeugen einen Schallschatten auf dem Ohr, welcher weiter von der Schallquelle entfernt ist. Ist die Wellenlänge kürzer als der Kopf, führt die Beugung des Schalls zu annähernd gleichen Schallintensitäten auf beiden Ohren.

Klangwahrnehmung an der Ohrmuschel
[Bearbeiten]Schallwellen, welche von hinten oder von vorne auf die Ohrmuschel treffen werden wegen der Form der Ohrmuschel verschieden beeinflusst. Die Schallwellen werden reflektiert und gedämpft oder verstärkt. Diese Änderungen helfen später bei der Schallquellen-Lokalisierung. Im äußeren Gehörgang werden Töne zwischen 3 und 12 kHz - ein für die menschliche Kommunikation entscheidender Bereich - verstärkt. Er dient als Resonator, der die eingehenden Frequenzen verstärkt. Schalleitung in der Cochlea Der Schall, der in Form von Wellen in die Ohrmuschel eindringt, wandert entlang des Gehörgangs bis zum Beginn des Mittelohrs, das durch das Trommelfell gekennzeichnet ist. Da das Innenohr mit Flüssigkeit gefüllt ist, ist das Mittelohr eine Art Impedanzanpassungsgerät, um das Problem der Schallenergie-Reflexion beim Übergang von Luft zur Flüssigkeit zu lösen. Beispielsweise werden beim Übergang von Luft zu Wasser 99,9 % der einfallenden Schallenergie reflektiert. Diese kann mit der Intensität des reflektierten Schalls (Ir), der Intensität des einfallenden Schalls (li) und dem Wellenwiderstand der beiden Medien (Zk) ( ZLuft = 414 kg m-2 s-1 und ZWasser = 1,48*106 kg m-2 s-1) berechnet werden.
wobei Ir die Intensität der reflektieren Schallwelle ist, Ii die Intensität der einlaufenden Schallwelle, und Zk der Wellenwiederstand der beiden Medien( Zair = 414 kg m-2 s-1 and Zwater = 1.48*106 kg m-2 s-1).
Drei Faktoren, die zur Impedanzanpassung beitragen, sind:
- der relative Größenunterschied zwischen Trommelfell und Ovalem Fenster
- die Hebelwirkung der Mittelohrknöchelchen und
- die Form des Trommelfells.

Die Luftdruckschwankungen der Schallwellen lassen das Trommelfell vibrieren, was wiederum die drei verketteten Gehörknöchelchen Hammer, Incus und Steigbügel synchron schwingen lässt. Diese Knochen schwingen als Einheit und heben die Energie vom Trommelfell zum ovalen Fenster. Zusätzlich wird die Schallenergie durch den räumlichen Unterschied zwischen dem Trommelfell und der Steigbügel-Fußplatte noch verstärkt. Das Mittelohr wirkt wie ein Impedanzwandler, indem es die vom Trommelfell aufgenommene Schallenergie in größere Kraft und weniger Auslenkung umwandelt. Dieser Mechanismus erleichtert die Übertragung von Schallwellen in der Luft in Schwingungen der Flüssigkeit in der Cochlea. Die Umwandlung erfolgt durch die kolbenartige Ein- und Auswärtsbewegung des Steigbügels, der sich im ovalen Fenster befindet. Diese Bewegung der Fußplatte setzt die Flüssigkeit in der Kochlea in Bewegung.
Durch den Stapedius-Muskel, den kleinsten Muskel im menschlichen Körper, hat das Mittelohr eine Anpassungsfunktion: Die Kontraktion dieses Muskels verändert die Impedanz des Mittelohrs und schützt so das Innenohr vor Schäden durch laute Geräusche.
Frequenzanalyse in der Cochlea
[Bearbeiten]Die drei flüssigkeitsgefüllten Bereiche der Cochlea (Scala vestibuli, Scala media, Scala tympani) sind durch die Basilarmembran und die Reissner-Membran getrennt. Die Funktion der Cochlea ist es, Klänge nach ihrem Spektrum zu trennen und in einen neuronalen Code umzuwandeln. Wenn die Fußplatte des Steigbügels in die Perilymphe der Scala vestibuli drückt, biegt sich die Reissner-Membran in die Scala media. Diese Dehnung der Reissner-Membran bewirkt eine Bewegung der Endolymphe innerhalb der Scala media und eine Verschiebung der Basilarmembran. Die Trennung der Schallfrequenzen in der Cochlea ist auf die besonderen Eigenschaften der Basilarmembran zurückzuführen. Die Flüssigkeit in der Cochlea vibriert (durch Ein- und Auswärtsbewegung der Steigbügel-Fußplatte) und setzt die Membran wie eine Wanderwelle in Bewegung. Die Welle beginnt an der Basis und verbreitet sich bis zum Apex der Cochlea. Die Transversalwellen in der Basilarmembran breiten sich aus mit:
Mit dem Schubmodul μ und der Dichte ρ des Materials. Da sich Breite und Spannung der Basilarmembran ändern, ändert sich die Geschwindigkeit der sich entlang der Membran ausbreitenden Wellen von etwa 100 m/s in der Nähe des ovalen Fensters auf 10 m/s in der Nähe des Apex.
Es gibt einen Punkt entlang der Basilarmembran, an dem die Amplitude der Welle abrupt abnimmt. An diesem Punkt erzeugt die Schallwelle in der Cochlea-Flüssigkeit die maximale Verschiebung (Spitzenamplitude) der Basilarmembran. Die Entfernung, die die Welle zurücklegt, bevor sie diesen charakteristischen Punkt erreicht, hängt von der Frequenz des ankommenden Schalls ab. Daher entspricht jeder Punkt auf der Basilarmembran einem bestimmten Wert der Stimulationsfrequenz. Ein niederfrequenter Schall legt eine längere Strecke zurück als ein hochfrequenter Schall, bevor er seinen charakteristischen Punkt erreicht. Die Frequenzen sind entlang der Basilarmembran skaliert, hohe Frequenzen an der Basis und tiefe Frequenzen an der Spitze der Cochlea.

Sensorische Transduktion in der Cochlea
[Bearbeiten]Die meisten Alltagsgeräusche bestehen aus mehreren Frequenzen. Das Gehirn verarbeitet die verschiedenen Frequenzen, nicht die gesamten Töne. Aufgrund ihrer inhomogenen Eigenschaften führt die Basilarmembran eine Annäherung an eine Fourier-Transformation durch. Der Schall wird dabei in seine verschiedenen Frequenzen aufgeteilt, und jede Haarzelle auf der Membran entspricht einer bestimmten Frequenz. Die Lautstärke der Frequenzen wird durch die Feuergeschwindigkeit der entsprechenden Afferenz kodiert.

Die Sinneszellen des Gehörs, die so genannten Haarzellen, befinden sich entlang der Basilarmembran im Corti-Organ. Jedes Corti-Organ enthält etwa 16.000 solcher Zellen, die durch etwa 30.000 afferente Nervenfasern angeregt werden. Es gibt zwei anatomisch und funktionell unterschiedliche Typen von Haarzellen: die inneren und die äußeren Haarzellen. Beide Typen von Haarzellen sind entlang der Basilarmembran angeordnet. Eine Reihe von inneren Haarzellen und drei bis fünf Reihen von äußeren Haarzellen. Der größte Teil der afferenten Innervation kommt von den inneren Haarzellen, während der größte Teil der efferenten Innervation an die äußeren Haarzellen geht. Die inneren Haarzellen beeinflussen die Entladungsrate der einzelnen Hörnervenfasern, die mit diesen jeweiligen Haarzellen verbunden sind. Deshalb übertragen innere Haarzellen Schallinformationen an höhere auditorische Nervenzentren. Die äußeren Haarzellen hingegen verstärken die Bewegung der Basilarmembran, indem sie Energie in die Bewegung der Membran injizieren und Reibungsverluste reduzieren. Sie tragen aber nicht zur Übertragung von Schallinformationen bei. Die Bewegung der Basilarmembran lenkt die Stereozillen (Haare auf den Haarzellen) ab und bewirkt, dass die intrazellulären Potentiale der Haarzellen je nach Richtung der Ablenkung abnehmen (Depolarisation) oder zunehmen (Hyperpolarisation). Wenn sich die Stereozilien in Ruhestellung befinden, fließt ein stationärer Strom durch die Kanäle der Zellen. Die Bewegung der Stereozilien moduliert also den Stromfluss um diesen stationären Strom.
Betrachten wir die Wirkungsweise der beiden verschiedenen Haarzelltypen getrennt:
- Innere Haarzellen:
Die Ablenkung der Stereozilien der Haarzellen öffnet mechanisch geschlossene Ionenkanäle. Nun können kleine, positiv geladene Kalium Ionen (K+) in die Zelle eindringen und sie depolarisieren. Im Gegensatz zu vielen anderen elektrisch aktiven Zellen löst die Haarzelle selbst kein Aktionspotential aus. Stattdessen depolarisiert der Zustrom von positiven Ionen aus der Endolymphe in der Scala media die Zelle, was zu einem Rezeptorpotential führt. Dieses Rezeptorpotential öffnet spannungsgesteuerte Kalziumkanäle; Kalzium Ionen (Ca2+) gelangen dann in die Zelle und lösen die Freisetzung von Neurotransmittern am basalen Ende der Zelle aus. Die Neurotransmitter diffundieren über den engen Spalt zwischen der Haarzelle und einem Nervenende, wo sie dann an Rezeptoren binden und so Aktionspotenziale im Nerv auslösen. Auf diese Weise erhöht der Neurotransmitter die Feuerrate im VIII. Hirnnerv und das mechanische Schallsignal wird in ein elektrisches Nervensignal umgewandelt. Die Repolarisation in der Haarzelle erfolgt in besonderer Weise. Die Perilymphe in der Scala tympani hat eine sehr niedrige Konzentration an positiven Ionen. Der elektrochemische Gradient lässt die positiven Ionen durch Kanäle zur Perilymphe fließen.
- Äußere Haarzellen:
In den äußeren Haarzellen des Menschen löst das Rezeptorpotential aktive Schwingungen des Zellkörpers aus. Diese mechanische Reaktion auf elektrische Signale wird als somatische Elektromobilität bezeichnet und treibt Schwingungen in der Länge der Zelle an, die bei der Frequenz des eingehenden Schalls auftreten und eine mechanische Rückkopplungsverstärkung bewirken. Äußere Haarzellen haben sich nur bei Säugetieren entwickelt. Ohne funktionierende äußere Haarzellen nimmt die Empfindlichkeit um ca. 50 dB ab (durch größere Reibungsverluste, die die Bewegung der Basilarmemran dämpfen würden). Die äusseren Haarzellen haben auch die Frequenzselektivität (Frequenzdiskriminierung) verbessert, was besonders für den Menschen von Vorteil ist, da es anspruchsvolle Sprache und Musik ermöglicht.
Ohne externe Stimulation leiten die Hörnervenfasern Aktionspotenziale in einer zufälligen zeitlichen Abfolge ab. Dieses zeitlich zufällige Feuern wird als spontane Aktivität bezeichnet. Die Spontanentladungsraten der Fasern variieren von sehr langsamen bis hin zu sehr hohen Raten von bis zu 100 pro Sekunde. Die Fasern werden in drei Gruppen eingeteilt, je nachdem, ob sie spontan mit hoher, mittlerer oder niedriger Geschwindigkeit feuern. Fasern mit hohen Spontanraten (> 18 pro Sekunde) neigen dazu, empfindlicher auf Schallstimulationen zu reagieren als andere Fasern.
Hörbahn der Nervenimpulse
[Bearbeiten]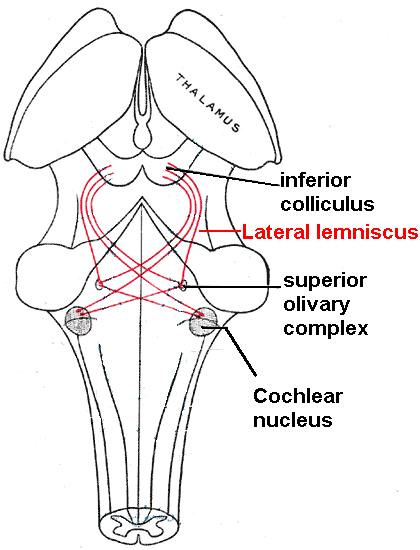
In den inneren Haarzellen wird das mechanische Schallsignal schließlich in elektrische Nervensignale umgewandelt. Die inneren Haarzellen sind mit Hörnervenfasern verbunden, deren Kerne das Ganglion cochleare bilden. Die zentralen Fortsätze des Ganglion cochleares bilden die cochleare Wurzel des Nervus vestibulocochlearis (Nervus VIII). Die elektrischen Signale (elektrische Spikes, Aktionspotentiale) werden hier weitergeleitet bis zu den Nuclei cochleares (Schneckenkerne) im Hirnstamm.
Von dort aus werden die akustischen Informationen in mindestens zwei Ströme aufgeteilt:
- Ventraler Nucleus cochlearis:
Der ventrale Nucleus cochlearis wird weiter in den posteroventralen Nucleus cochlearis und den anteroventralen Nucleus cochlearis aufgeteilt. Die Zellen des ventralen Nucleus cochlearis projizieren zu einer Ansammlung von Kernen, der so genannten Nucleus olivari superior (obere Olivenkerne).
Nucleus olivari superior: Klanglokalisierung
[Bearbeiten]Der Nucleus olivari superior - eine kleine Masse an grauer Substanz - soll an der Lokalisation von Tönen in der azimutalen Ebene (d.h. ihrem Grad nach links oder rechts) beteiligt sein. Es gibt zwei wichtige Anhaltspunkte für die Tonlokalisierung: Interaurale Pegeldifferenzen (ILD) und interaurale Zeitdifferenzen (ITD). Das ILD misst Unterschiede in der Schallintensität zwischen den Ohren. Dies funktioniert bei hohen Frequenzen (über 1,6 kHz), bei denen die Wellenlänge kürzer ist als der Abstand zwischen den Ohren, wodurch ein Kopfschatten entsteht. Das bedeutet, dass hochfrequente Töne mit geringerer Intensität auf das abgewandte Ohr treffen. Niederfrequente Töne werfen keinen Schatten. Da die Wellenlänge jedoch größer ist als der Abstand zwischen den Ohren, gibt es eine Phasendifferenz zwischen den Schallwellen, die in die Ohren eindringen - die vom ITD gemessene Zeitdifferenz. Dies funktioniert sehr präzise bei Frequenzen unter 800 Hz, bei denen der Ohrabstand kleiner als die Hälfte der Wellenlänge ist. Die Schallortung in der Mittelebene (vorne, oben, hinten, unten) wird durch das Außenohr unterstützt, das ein richtungsselektiver Filter bildet.
In den oberen Olivenkernen werden die Zeit- und Lautstärkeunterschiede der Schallinformationen beider Ohren verglichen. Unterschiede in der Schallintensität werden in den Zellen des lateralen Nucleus olivari superior und zeitliche Unterschiede (Laufzeitverzögerungen) im medialen Nuchleus olivari superior verarbeitet. Der Mensch kann Zeitunterschiede zwischen dem linken und rechten Ohr von bis zu 10 µs erkennen, entsprechend einem Schallortungsunterschied von einem Grad. Dieser Vergleich der Schallinformationen beider Ohren ermöglicht die Bestimmung der Richtung, aus der der Schall kommt. Die obere Olive ist der erste Knoten, an dem die Signale beider Ohren zusammenlaufen und verglichen werden können. Als nächster Schritt werden die Informationen vom Nucleus olivari superior über den Lemniscus lateralis zum Colliculus inferior geleitet. Die Funktion des Colliculus inferior besteht darin, Informationen zu integrieren, bevor sie an den Thalamus und den auditiven Kortex gesendet werden. Es ist interessant zu wissen, dass der obere Colliculus superior, welcher gleich in der Nähe ist, ein Zusammenspiel von akustischen und visuellen Reizen zeigt.
- Dorsaler Nucleus cochlearis:
Der dorsale Nucleus cochlearis (DCN) analysiert die Klangqualität und projiziert direkt über den Lemniscus lateralis in den Colliculus inferior.
Vom Colliculus inferior geht die auditive Information sowohl vom ventralen als auch vom dorsalen Nucleus cochlearis in den Hörkern des Thalamus über, der Corpus geniculatum mediale (mediale Kniehöcker) hiesst. Der mediale Kniehöcker überträgt Informationen weiter an den primären auditiven Kortex, welcher sich auf dem Temporallappen befindet. Der primäre auditive Kortex ist die Region des menschlichen Gehirns, die für die Verarbeitung der auditiven Informationen verantwortlich ist und ist das erste Relais, das an der bewussten Wahrnehmung von Schall beteiligt ist.
Primärer auditiver Kortex und höhere auditive Zentren
[Bearbeiten]Der primäre auditive Kortex (Brodmann Areal 41 und 42) ist das erste Relais, das an der bewussten Wahrnehmung von Schall beteiligt ist. Er ist tonotopisch organisiert und vermittelt die Grundlagen des Hörens: Tonhöhe und Lautstärke. Je nach Art des Klangs (Sprache, Musik, Lärm), wird die Information weiter in höhere auditive Gebiete geleitet. Worte werden im Wernickes Areal (Brodmann Areal 22) verarbeitet. Dieser Bereich beschäftigt sich mit dem Verstehen der geschriebenen und gesprochenen Sprache (verbales Verstehen). Die Produktion von Klang (verbaler Ausdruck) findet im Broca Areal (Brodmann Areal 44 und 45) statt. Die Muskeln, die beim Sprechen den gewünschten Klang erzeugen, werden durch den Gesichtsbereich des motorischen Kortex angesteuert. Diese Bereiche der Großhirnrinde sind an der Planung, Steuerung und Ausführung von freiwilligen motorischen Funktionen beteiligt. Bild