Arqueus
| Archaea | |
|---|---|
 | |
| Dades | |
| Mitjà de locomoció | rodolament |
| Descobridor o inventor | Carl Richard Woese i George Edward Fox |
| Període | |
| Taxonomia | |
| Domini | Archaea Woese, Kandler i Wheelis, 1990 |
| Tipus taxonòmic | Methanobacterium |
| Nomenclatura | |
| Sinònims |
|
| Significat | ‘[Organismes] arcaics’ |
| Clades | |

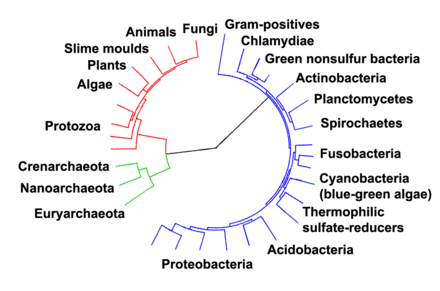
Els arqueus (Archaea), anteriorment coneguts com a arqueobacteris (Archaeobacteria),[2] són un domini d'organismes unicel·lulars. Els arqueus, com els eubacteris, són procariotes que manquen de nucli cel·lular o qualsevol altre orgànul dins les cèl·lules. En el passat, se'ls considerà un grup inusual de bacteris, però com que tenen una història evolutiva independent i presenten moltes diferències en la seva bioquímica respecte a la resta de formes de vida, actualment se'ls classifica com un domini distint en el sistema de tres dominis. En aquest sistema, presentat per Carl Woese, les tres branques evolutives principals són els arqueus, els bacteris i els eucariotes. Els arqueus estan subdividits en quatre fílums, dels quals dos, els crenarqueots i els euriarqueots, són estudiats més intensivament. La classificació dels arqueus roman dificultosa, car la immensa majoria d'aquests organismes no han sigut mai estudiats al laboratori i només se'ls ha detectat per mitjà de l'anàlisi dels seus àcids nucleics en mostres del medi ambient.
En general, els arqueus i bacteris són bastant semblants en forma i en mida, tot i que alguns arqueus tenen formes molt inusuals, com ara les cèl·lules planes i quadrades de Haloquadra walsbyi. Malgrat aquesta semblança visual amb els bacteris, els arqueus tenen gens i diverses rutes metabòliques que són més properes a les dels eucariotes; notablement, els enzims implicats en la transcripció i la traducció. Altres aspectes de la bioquímica dels arqueus són únics, com ara els èters lípids de les seves membranes cel·lulars. Els arqueus exploten una varietat de recursos molt més grans que els eucariotes; des de compostos orgànics familiars com ara els sucres, fins a l'ús d'amoníac, ions de metalls o fins i tot hidrogen com nutrients. Els arqueus tolerants de la sal utilitzen la llum solar com a font d'energia, i altres espècies d'arqueus fixen carboni; tanmateix, a diferència de les plantes i els cianobacteris, no es coneix cap espècie d'arqueu que sigui capaç d'ambdues coses. Els arqueus es reprodueixen asexualment i es divideixen per fissió binària, fragmentació o gemmació; a diferència dels bacteris i els eucariotes, no es coneix cap espècie d'arqueu que formi espores.
Originalment, els arqueus eren vists com a extremòfils que vivien en ambients severs, com ara aigües termals i llacs salats, però posteriorment se'ls ha observat en una gran varietat d'hàbitats, com ara sòls, oceans i aiguamolls. Els arqueus són especialment nombrosos als oceans, i els que es troben al plàncton podrien ser un dels grups d'organismes més abundants del planeta. Aquests procariotes són actualment considerats una part important de la vida a la Terra i podrien tenir un paper important tant en el cicle del carboni com en el cicle del nitrogen. No es coneixen exemples clars de patògens o paràsits que siguen arqueus, però solen ser mutualistes o comensals. En són exemples els arqueus metanògens que viuen a l'intestí dels humans i els remugants, on estan presents en grans quantitats i contribueixen a digerir l'aliment. Els arqueus tenen la seva importància en la tecnologia; hi ha metanògens que són utilitzats per produir biogàs i com a part del procés de depuració, i els enzims d'arqueus extremòfils capaços de resistir a temperatures elevades i solvents orgànics són explotats en la biotecnologia.
El 2012 es va demostrar per primera vegada, que els arqueus són consumits per organismes vius, en concret per part de cucs poliquets dins la família dels dorvil·lèids. L'estudi es va publicar al Society for Microbial Ecology Journal, de la revista Nature.[3]
Classificació
[modifica]Un nou domini
[modifica]A principis del segle xx, els procariotes eren considerats un únic grup d'organismes i classificats segons la seva bioquímica, morfologia i metabolisme. Per exemple, els microbiòlegs intentaven classificar els microorganismes segons l'estructura de la seva paret cel·lular, la seva forma, i les substàncies que consumeixen.[4] Tanmateix, el 1965 es proposà un nou sistema,[5] utilitzant les seqüències genètiques d'aquests organismes per esbrinar quins procariotes estan realment relacionats entre ells. Aquest mètode, conegut com a filogènia, és el mètode principal utilitzat actualment.

Els arqueus foren classificats per primer cop com un grup separat de procariotes el 1977 per Carl Woese i George E. Fox en arbres filogenètics basats en les seqüències de gens d'ARN ribosòmic (ARNr).[6] Aquests dos grups foren originalment denominats arqueobacteris i eubacteris i tractats com regnes o subregnes, que Woese i Fox denominaren «regnes originals». Woese argumentà que aquest gup de procariotes és un tipus de vida fonamentalment distint. Per emfatitzar aquesta diferència, aquests dos dominis foren posteriorment anomenats Archaea i Eubacteria.[7] El nom científic Archaea prové del grec antic ἀρχαῖα, que vol dir 'els antics'.[8] El terme «arqueobacteri» prové de la combinació d'aquesta arrel i del terme grec baktērion, que significa 'bastonet'.
Originalment, només es classificaren els metanògens en aquest nou domini, i els arqueus eren considerats extremòfils que només viuen en hàbitats com ara aigües termals i llacs salats. A finals del segle xx, els microbiòlegs s'adonaren que els arqueus són un grup d'organismes gran i divers àmpliament distribuïts a la natura, i que són comuns en hàbitats molt menys extrems, com ara sòls i oceans.[9] Aquesta nova presa de consciència de la importància i l'omnipresència vingué de l'ús de la reacció en cadena de la polimerasa per detectar procariotes en mostres d'aigua o sòl a partir únicament dels seus àcids nucleics. Això permet detectar i identificar organismes que no poden ser cultivats al laboratori, que sol ser difícil.[10][11]
Classificació actual
[modifica]La classificació dels arqueus, i dels procariotes en general, és un camp en constant moviment i controvertit. Els sistemes actuals de classificació intenten organitzar els arqueus en grups d'organismes que comparteixen trets estructurals i avantpassats comuns.[12] Aquestes classificacions es basen significativament en l'ús de seqüències de gens d'ARN ribosòmic per revelar les relacions entre els organismes (filogenètica molecular).[13] La majoria d'espècies d'arqueus cultivables i ben investigades són membres de dos fílums principals, els euriarqueots i crenarqueots. S'han proposat temptativament altres grups. Per exemple, a la peculiar espècie Nanoarchaeum equitans, que fou descoberta el 2003, se li ha atribuït el seu propi fílum, el dels nanoarqueots.[14] També s'ha proposat el nou fílum dels korarqueots, que conté un nombre reduït d'inusuals espècies termòfiles que comparteixen trets dels dos fílums principals, però que són més properes als crenarqueots.[15][16] Altres espècies d'arqueus recentment descobertes només tenen una relació distant amb qualsevol d'aquests grups, com ara els nanoorganismes arquobacterians acidòfils de Richmond Mine (ARMAN), descoberts el 2006.[17]

La classificació dels arqueus en espècies també és controvertida. En biologia, una espècie és un grup d'organismes relacionats. Una definició popular d'espècie entre els animals és un conjunt d'organismes que poden reproduir-se entre ells i que estan reproductivament aïllats d'altres grups d'organismes (és a dir, no poden reproduir-se amb altres espècies).[18] Tanmateix, els esforços per classificar els procariotes tals com els arqueus en espècies es compliquen pel fet que són asexuals i que presenten un alt nivell de transferència horitzontal de gens entre llinatges. Aquest tema és controvertit; per exemple, algunes dades suggereixen que en arqueus com ara Ferroplasma, es poden agrupar cèl·lules individuals en poblacions de genoma molt similar i que rarament transfereixen gens a grups més divergents de cèl·lules.[19] Alguns argumenten que aquests grups de cèl·lules són anàlegs a espècies. D'altra banda, estudis d'Halorubrum descobriren un intercanvi genètic significatiu entre aquestes poblacions.[20] Aquests resultats han portat a pensar que classificar aquests grups d'organismes com espècies tindria poc sentit pràctic.[21]
El coneixement actual sobre la diversitat dels arqueus és fragmentari, i no es pot estimar amb cap mena de precisió el nombre total d'espècies d'arqueus.[13] Fins i tot les estimacions del nombre total de fílums arquobacterians varien entre 18 i 23, dels quals només vuit tenen representants que s'han cultivat i estudiat directament. Molts d'aquests grups hipotètics són coneguts únicament a partir d'una sola seqüència d'ARNr, cosa que indica que la diversitat d'aquests organismes roman completament desconeguda.[22] El problema de com estudiar i classificar microbis no cultivats també es dona en els bacteris.[23]
Origen i evolució
[modifica]Els arqueus són una forma de vida molt antiga. S'han datat fòssils probables d'aquestes cèl·lules de fa gairebé 3.500 milions d'anys,[24] i s'han detectat restes de lípids que podrien ser d'arqueus o d'eucariotes en esquists de fa 2.700 milions d'anys.[25] Com que la majoria de procariotes no tenen una morfologia característica, la forma dels fòssils no serveix per identificar-los com arqueus. En canvi, fòssils químics, en forma dels lípids exclusius dels arqueus, ofereixen més informació car aquests compostos no apareixen en altres grups d'organismes.[26] Aquests lípids han sigut detectats en roques que daten del Precambrià. Les restes més antigues conegudes d'aquests lípids d'isoprè daten del cinturó d'Isua, a l'oest de Groenlàndia, que contenen sediments formats fa 3.800 milions d'anys que són els més antics del món.[27] L'origen dels arqueus sembla extremament remot en el temps, i el llinatge dels arqueus podria ser el més antic de la Terra.[28]
Woese argumentà que els bacteris, arqueus i eucariotes representen cadascun un llinatge evolutiu distint que divergí fa molts milions d'anys d'una colònia ancestral d'organismes.[30][31] Tanmateix, alguns biòlegs han argumentat que els arqueus i eucariotes sorgiren d'un grup de bacteris.[32] Thomas Cavalier-Smith proposà el clade dels neomurs per representar aquesta teoria. Neomura vol dir 'parets noves' i es refereix a la teoria que els arqueus i els eucariotes haurien derivat de bacteris que (entre altres adaptacions) havien substituït les parets de peptidoglicà per altres glicoproteïnes. És possible que l'últim avantpassat comú dels bacteris i els arqueus fos un termòfil, cosa que presenta la possibilitat que les temperatures baixes siguin «ambients extrems» per als arqueus, i que els organismes que viuen en ambients més freds apareguessin més tard en la història de la vida a la Terra.[33] Com que els arqueus i els bacteris no estan relacionats més estretament els uns amb els altres que amb els eucariotes, s'ha argumentat que el terme «procariota» no té un sentit evolutiu autèntic i que caldria descartar-lo completament.[34]
La relació entre arqueus i eucariotes roman un problema important. A part de les semblances en l'estructura i les funcions cel·lulars que es discuteixen més avall, molts arbres genètics agrupen els dos clades. Algunes de les primeres anàlisis fins i tot suggerien que la relació entre els eucariotes i el fílum d'arqueus dels euriarqueots és més propera que la relació entre els euriarqueots i els crenarqueots.[35] Tanmateix, actualment es considera més versemblant que l'avantpassat dels eucariotes divergís bastant aviat dels arqueus.[36][37] El descobriment de gens similars als dels arqueus en alguns grups de bacteris, com ara Thermotoga maritima, fa que aquestes relacions siguin difícils de determinar, car ha tingut lloc una transferència horitzontal de gens.[38] Alguns han suggerit que els eucariotes sorgiren de la fusió d'un arqueu i un bacteri, que esdevingueren el nucli i el citoplasma; això explica diverses semblances genètiques però presenta dificultats a l'hora d'explicar l'estructura cel·lular.[39]
Filogènesi
[modifica]Les anàlisis filogenètiques recents han obtingut les següents relacions entre els grups de arqueus.[40][41][42]
| Cytota |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Morfologia
[modifica]
La mida dels arqueus varia entre 0,1 micròmetres (μm) i més de 15 μm de diàmetre, i existeixen en diverses formes, habitualment esferes, bastons, espirals o plaques.[43] Altres morfologies dels crenarqueots inclouen cèl·lules lobulades de forma irregular en Sulfolobus, filaments prims semblants a agulles de menys d'un micròmetre de diàmetre en Thermofilum, i barres gairebé perfectament rectangulars a Thermoproteus i Pyrobaculum.[44] Fins i tot existeix una espècie d'arqueu pla i quadrada anomenada Haloquadra walsbyi, que viu en basses hipersalines.[45] Aquestes formes inusuals són suportades probablement tant per les seves parets cel·lulars com per un citoesquelet procariota. En els arqueus existeixen proteïnes relacionades amb els components del citoesquelet d'altres organismes,[46] i es formen filament dins de les seves cèl·lules,[47] però a diferència d'altres organismes, aquestes estructures cel·lulars no estan ben compreses en els arqueus.[48]
Algunes espècies d'arqueus formen agregats o filaments de cèl·lules de fins a 200 μm de llargada,[43] i aquests organismes poden ser membres prominents de les comunitats microbials que produeixen biofilms.[49] N'és un exemple extrem Thermococcus coalescens, car agregats d'aquestes cèl·lules es fusionen en cultiu, formant cèl·lules úniques gegants.[50] Una forma especialment elaborada de colònia pluricel·lular és la dels arqueus del gènere Pyrodictium. Les seves cèl·lules produeixen agrupacions de llargs i prims tubs buits que reben el nom de cànules, que sobresurten de la superfície de les cèl·lules i les connecten en una densa colònia semblant a un arbust.[51] Es desconeix la funció d'aquestes cànules, però podrien permetre a les cèl·lules comunicar-se o intercanviar nutrients amb les seves veïnes.[52] També es poden formar colònies per associació entre espècies diferents. Per exemple, en una comunitat que fou descoberta el 2001 en un aiguamoll alemany, hi havia colònies blanques i rodones d'una nova espècie d'arqueu del fílum dels euriarqueots escampades al llarg de filaments prims que poden mesurar fins a 15 cm de llarg; aquests filaments es componen d'una espècie particular de bacteris.[53]
Estructura cel·lular
[modifica]Els arqueus són semblants als bacteris en la seva estructura cel·lular general, però la composició i organització d'algunes d'aquestes estructures diferencien els arqueus. Com els bacteris, els arqueus manquen de membranes interiors, de manera que les seves cèl·lules no contenen orgànuls.[34] També s'assemblen als bacteris en què la seva membrana cel·lular està habitualment delimitada per una paret cel·lular i en què neden per mitjà d'un o més flagels.[54] En la seva estructura general, els arqueus s'assemblen especialment als bacteris grampositius, car la majoria tenen una única membrana plasmàtica i paret cel·lular, i manquen d'espai periplàsmatic; l'excepció d'aquesta regla general és l'arqueu Ignicoccus, que té un espai periplasmàtic particularment gran que conté vesícules limitades per membranes, i que queda tancat per una membrana exterior.[55]
Membrana cel·lular
[modifica]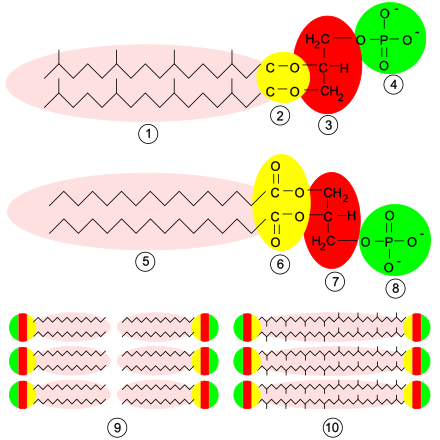
Les membranes dels arqueus es componen de molècules que difereixen molt de les que es troben en altres formes de vida, cosa que és una prova que els arqueus només tenen una relació distant amb els bacteris i eucariotes.[56] En tots els organismes, les membranes cel·lulars es componen de molècules conegudes com a fosfolípids. Aquestes molècules tenen una part polar que es dissol en l'aigua (el «cap» de fosfats), i una part «greixosa» no polar que no es dissol en l'aigua (la «cua» de lípids). Aquestes parts diferents queden connectades per un grup glicerol. Dins l'aigua, els fosfolípids s'aglomeren, amb els caps de fosfats polars vers l'aigua i les cues lipídiques no polars lluny de l'aigua. Això fa que s'estructurin en capes. L'estructura principal de la membrana cel·lular és una doble capa d'aquests fosfolípids, que rep el nom de bicapa lipídica.
Els fosfolípids de les membranes dels arqueus són inusuals en quatre coses. Primerament, els bacteris tenen membranes compostes principalment de lípids èster amb glicerol, mentre que els arqueus les tenen compostes de lípids èter amb glicerol.[57] La diferència entre aquests dos tipus de fosfolípids és el tipus d'enllaç que uneix els lípids al grup glicerol; aquests dos tipus d'enllaç apareixen en groc a la imatge de la dreta. En els lípids èster hi ha un enllaç èster, mentre que en la resta de lípids hi ha un enllaç èter. Els enllaços èter tenen una resistència química superior a la dels enllaços èster, cosa que podria contribuir a la capacitat d'alguns arqueus de sobreviure a temperatures extremes o en ambients molt àcids o alcalins.[58] Els eubacteris i eucariotes també contenen alguns lípids èter, però a diferència dels arqueus, aquests lípids no formen una part important de les seves membranes.
Segonament, els lípids arquobacterians són únics perquè l'estereoquímica del grup glicerol és la inversa que la que s'observa en altres organismes. El grup glicerol pot existir en dues formes que són la imatge inversa l'una de l'altra, i que es poden denominar formes dretana i esquerrana; en llenguatge químic se les anomena enantiòmers. De la mateixa manera que una mà dreta no entra fàcilment dins un guant per la mà esquerra, una molècula de glicerol dretana generalment no pot ser utilitzada o creada per enzims adaptats per la forma esquerrana. Això suggereix que els arqueus utilitzen enzims completament diferents per sintetitzar els seus fosfolípids dels que utilitzen els bacteris i eucariotes; com que aquests enzims es desenvoluparen molt al principi de la història de la vida, això suggereix al seu torn que els arqueus se separaren molt aviat dels altres dos dominis.[56]
Tercerament, les cues lipídiques dels fosfolípids dels arqueus tenen una composició química diferent de les d'altres organismes. Els lípids arquobacterians es basen en la cadena lateral isoprenoide i són llargues cadenes amb múltiples branques laterals i, a vegades, fins i tot anells de ciclopropà o ciclohexà.[59] Això contrasta amb els àcids grassos que hi ha a les membranes d'altres organismes, que tenen cadenes rectes sense branques ni anells. Tot i que els isoprenoides tenen un paper important en la bioquímica de molts organismes, només els arqueus els utilitzen per produir fosfolípids. Aquestes cadenes amb branques podrien ajudar a evitar que les membranes dels arqueus tinguin fuites a altes temperatures.[60]
Finalment, en alguns arqueus la bicapa lipídica és substituïda per una única monocapa. De fet, els arqueus fusionen les cues de dues molècules fosfolipídiques independents en una única molècula amb dos caps polars; aquesta fusió podria fer la seva membrana més rígida i més apta per resistir ambients severs.[61] Per exemple, tots els lípids de Ferroplasma són d'aquest tipus, cosa que es creu que ajuda aquest organisme a sobreviure en els medis extraordinàriament àcids en què habita.[62]
Paret cel·lular i flagels
[modifica]La majoria d'arqueus tenen una paret cel·lular; les excepcions són Thermoplasma i Ferroplasma.[63] En la majoria d'arqueus , la paret es compon de proteïnes de capa de superfície, que formen una capa S.[64] Una capa S és una agrupació rígida de molècules proteíniques que cobreixen l'exterior de la cèl·lula com una cota de malla.[65] Aquesta capa ofereix una protecció química i física, i pot servir de barrera, impedint que entrin en contacte macromolècules amb la membrana cel·lular.[66] A diferència dels bacteris, la majoria d'arqueus no tenen peptidoglicà a la paret cel·lular.[67] L'excepció és el pseudopeptidoglicà, que es troba en els metanobacterials, però aquest polímer és diferent del peptidoglicà bacterià, ja que manca de D-aminoàcids i àcid N-acetilmuràmic.[66]
Els arqueus també tenen flagels, que funcionen d'una manera semblant als flagels bacterians —són llargs palets moguts per motors rotatoris situats a la base dels flagels. Aquests motors són impulsats pel gradient de protons de la membrana. Tanmateix, els flagels dels arqueus són notablement diferents en la seva composició i el seu desenvolupament.[54] Cada tipus de flagel evolucionà d'un avantpassat diferent; el flagel bacterià evolucionà d'un sistema de secreció de tipus III, mentre que els flagels dels arqueus semblen haver evolucionat dels pili bacterians de tipus IV.[68] A diferència del flagel bacterià, que és un palet buit i que està format per subunitats que es mouen per la cavitat central i després s'afegeixen a la punta del flagel, els flagels dels arqueus se sintetitzen per mitjà de l'addició de subunitats a la seva base.[69]
Metabolisme
[modifica]Els arqueus presenten una gran varietat de reaccions químiques en el seu metabolisme, i utilitzen moltes fonts d'energia diferents. Aquestes formes de metabolisme es classifiquen en grups nuricionals, segons la font de l'energia i del carboni. Alguns arqueus obtenen l'energia de composts inorgànics com ara el sofre o l'amoníac (són litòtrofs). Aquests arqueus inclouen nitrificants, metanògens i oxidants anaeròbics de metà.[70] En aquestes reaccions, un compost passa electrons a l'altre (en una reacció redox), alliberant energia que és utilitzada per alimentar les activitats de les cèl·lules. Un compost actua com donador d'electrons i l'altre com acceptor d'electrons. Una característica comuna de totes aquestes reaccions és que l'energia alliberada és utilitzada per generar trifosfat d'adenosina (ATP) per mitjà de la quimiosmosi, que és el mateix procés bàsic que té lloc als mitocondris de les cèl·lules animals.[71]
Altres grups d'arqueus utilitzen la llum solar com font d'energia (són fotòtrofs). Tanmateix, cap d'aquests organismes no presenta una fotosíntesi generadora d'oxigen.[71] Moltes de les rutes metabòliques bàsiques són compartides per totes les formes de vida; per exemple, els arqueus utilitzen una forma modificada de la glicòlisi (la via Entner-Doudoroff), i un cicle de Krebs complet o parcial.[72] Aquestes semblances amb la resta d'organismes probablement reflecteixen tant l'evolució primerenca d'aquestes parts del metabolisme en la història de la vida, com el seu alt nivell d'eficiència.[73]
| Tipus nutricional | Font d'energia | Font de carboni | Exemples |
|---|---|---|---|
| Fotòtrofs | Llum solar | Compostos orgànics | Halobacteria |
| Litòtrofs | Compostos inorgànics | Compostos inorgànics o fixació del carboni | Ferroglobus, Methanobacteria o Pyrolobus |
| Organòtrofs | Compostos orgànics | Compostos orgànics o fixació del carboni | Pyrococcus, Sulfolobus o Methanosarcinales |
Alguns euriarqueots són metanògens i produeixen gas metà en ambients anaeròbics com ara aiguamolls. Aquest tipus de metabolisme evolucionà aviat, i fins i tot és possible que el primer organisme de vida lliure fos un metanogen.[74] Una reacció típica d'aquests organismes implica l'ús de diòxid de carboni com receptor d'electrons per oxiditzar hidrogen. La metanogènesi implica una varietat de coenzims que són únics a aquests arqueus, com ara el coenzim M o el metanofuran.[75] Altres compostos orgànics com ara alcohols, àcid acètic o àcid fòrmic són utilitzats com acceptors d'electrons pels metanògens. Aquestes reaccions són habituals en els arqueus intestinals. L'àcid acètic també és descompost en metà i diòxid de carboni directament, pels arqueus acetòtrofs. Aquests acetòtrofs són arqueus de l'ordre dels metanosarcinals, i són una part important de les comunitats de microorganismes productores de biogàs.[76]
Altres arqueus utilitzen el CO₂ de l'atmosfera com font de carboni, en un procés anomenat fixació del carboni (són autòtrofs). En els arqueus, aquest procés implica o bé una forma molt modificada del cicle de Calvin,[78] o una ruta metabòlica recentment descoberta coneguda com a cicle del 3-hidroxipropionat/4-hidroxibutirat.[79] Els crenarqueots també utilitzen el cicle de Krebs invers i els euriarqueots també utilitzen la via reductora acetil-CoA.[80] En aquests organismes, la fixació del carboni és alimentada per fonts inorgàniques d'energia, en lloc de ser alimentada per la llum solar com és el cas en les plantes i cianobacteris. No es coneixen arqueus que puguin dur a terme la fotosíntesi, que és quan la llum és utilitzada pels fotoautòtrofs com font d'energia a més de font d'aliment per la fixació del diòxid de carboni.[81] Les fonts d'energia utilitzades pels arqueus per fixar el carboni són extremament diverses, i van de l'oxidació de l'amoníac (per part dels nitrosopumilals en un metabolisme anammox)[82][83] fins a l'oxidació d'àcid sulfhídric o sofre elemental per part d'espècie de Sulfolobus, utilitzant oxigen o ions metàl·lics com acceptors d'electrons.[71]
Els arqueus fotòtrofs utilitzen llum per produir energia química en forma d'ATP. En els halobacteris hi ha bombes d'ions que s'activen per la llum, com ara la bacteriorodopsina i l'halorodopsina, que generen gradients d'ions per mitjà del bombament d'ions vers l'exterior de la cèl·lula a través de la membrana plasmàtica. L'energia emmagatzemada en aquests gradients electroquímics és posteriorment convertida en ATP per l'ATP sintasa.[43] Aquest procés és una forma de fotofosforilació. L'estructura i el funcionament d'aquestes bombes activades per la llum han sigut estudiats en gran detall, cosa que ha revelat que la seva capacitat de moure ions a través de les membranes depèn en uns canvis produïts per la llum en l'estructura d'un cofactor de retinol incorporat al centre de la proteïna.[84]
Genètica
[modifica]Els arqueus solen tenir un únic nucleoide, mal anomenat cromosoma circular,[85] la mida del qual pot arribar a 5.751.492 parells de bases en Methanosarcina acetivorans,[86] que té el genoma d'arqueu més gran seqüenciat fins al present. El genoma de Nanoarchaeum equitans és el genoma d'arqueu més petit conegut amb només 490.885 parells de bases, i es calcula que només conté 537 gens codificadors de proteïnes.[87] En els arqueus també s'observen petits fragments independents d'ADN, anomenats plasmidis. Els plasmidis poden moure's d'una cèl·lula a l'altra pel contacte físic, en un procés que pot ser similar a la conjugació bacteriana.[88][89]

Els arqueus poden ser infectats per virus d'ADN bicatenari, que no estan relacionats a cap altra forma de virus i que tenen una varietat de formes inusuals; alguns semblen ampolles, barres ganxudes o llàgrimes.[91] Aquests virus han sigut estudiats amb més detall en els arqueus termòfils, especialment en els ordres dels sulfolobals i els termoproteals.[92] Les defenses contra aquests virus poden implicar l'ús d'ARN interferent de seqüències d'ADN repetitiu dels genomes arquobacterians relacionades amb els gens dels virus.[93][94]
Els arqueus són genèticament distints dels bacteris i eucariotes, i fins a un 15% de les proteïnes que codifica un genoma propi dels arqueus són únics als arqueus, tot i que la majoria d'aquests gens únics manquen de funció coneguda.[95] De la resta de gens únics als arqueus que tenen una funció identificada, la majoria treballen en la metanogènesi. Els gens compartits pels arqueus, bacteris i eucariotes formen un nucli comú de funcions cel·lulars, implicat principalment en la transcripció, la traducció, i el metabolisme dels nucleòtids.[96] Altres característiques distintives dels genomes arquobacterians són l'organització de gens amb funcions relacionades – com ara enzims que catalitzen fases de la mateixa ruta metabòlica – en nous operons, així com grans diferències en els seus gens d'ARNt i les seves aminoacil ARNt sintetases.[96]
La transcripció i la traducció en els arqueus són més similars als processos dels eucariotes que els dels bacteris, i l'ARN polimerasa i els ribosomes arquobacterians són molt propers als seus homòlegs eucariotes.[85] Tot i que els arqueus només tenen un tipus d'ARN polimerasa, la seva estructura i funció en la transcripció sembla propera a la de l'ARN polimerasa II dels eucariotes, amb assemblatges proteics similars (els factors de transcripció generals) que dirigeixen la unió de l'ARN polimerasa al promotor d'un gen.[97] Tanmateix, altres factors de transcripció arquobacterians són més propers als que tenen els bacteris.[98] La modificiació posttranscripcional és més simple que en els eucariotes, car la majoria de gens arquobacterians manquen d'introns, tot i que hi ha molts introns en elseus gens de l'ARN de transferència i ARN ribosòmic,[99] i pot haver-hi introns en alguns dels seus gens codificadors de proteïnes.[100][101]
Reproducció
[modifica]Els arqueus es reprodueixen asexualment per fissió binària o múltiple, fragmentació, o gemmació; no experimenten meiosi, de manera que si una espècie d'arqueu existeix en més d'una forma, totes elles tindran el mateix material genètic.[43] La divisió cel·lular és controlada en els arqueus en un cicle cel·lular; un cop el cromosoma de la cèl·lula s'ha replicat i els dos cromosomes fills han quedat separats, la cèl·lula es divideix.[102] Els detalls del cicle cel·lular dels arqueus només han sigut investigats en el gènere Sulfolobus, però aquest té característiques que són similars tant als sistemes bacterians com als eucariotes. En aquest arqueu, els cromosomes es repliquen a partir de múltiples punts d'inici (orígens de replicació) utilitzant ADN polimerases que s'assemblen als enzims eucariotes equivalents.[103] Tanmateix, les proteïnes que controlen la divisió cel·lular, com ara la proteïna FtsZ, que forma un anell contraent al voltant de la cèl·lula, i els components del septe que es forma a través del centre de la cèl·lula, són similars als seus homòlegs bacterians.[102]
Tant els bacteris com els eucariotes produeixen espores, però aquest fet no ha sigut observat en cap arqueu conegut.[104] Algunes espècies d'haloarqueus experimenten dimorfisme fenotípic i creixen com diversos tipus diferents de cèl·lula, incloent-hi estructures amb parets gruixudes que són resistents als xocs osmòtics i que permeten als arqueus sobreviure en aigües amb concentracions baixes de sal, però aquestes no són estructures reproductives, sinó que podrien contribuir amb la dispersió a nous hàbitats.[105]
Ecologia
[modifica]Hàbitats
[modifica]Els arqueus existeixen en una gran varietat d'hàbitats, són una part important dels ecosistemes globals,[9] i podrien representar fins a un 20% del total de biomassa de la Terra.[106] Nombrosos arqueus són extremòfils, i aquest tipus d'hàbitat fou vist històricament com el seu nínxol ecològic.[70] De fet, alguns arqueus sobreviuen a altes temperatures, com la soca 121, sovint per sobre de 100 °C, com les que hi ha als guèisers, xemeneies mineralitzants, i pous de petroli. Altres viuen en hàbitats molt freds, i d'altres en aigües altament salines àcides o alcalines. Tanmateix, altres arqueus són mesòfils que viuen en condicions molt més suaus; aiguamolls, clavegueres, els oceans i el sòl.[9]

Els arqueus extremòfils són membres de quatre grups fisiològics principals. Són els halòfils, termòfils, alcalífils i acidòfils.[107] Aquests grups no són incloents ni tenen una relació amb el fílum al qual pertanyi un determinat arqueu, ni són mútuament exclusius, car alguns arqueus pertanyen a diversos d'aquests grups. Tanmateix, són útils com punt de partida per la classificació d'aquests organismes.
Els halòfils, incloent-hi el gènere Halobacterium, viuen en ambients extremament salins, com ara llacs salats, i comencen a superar els seus homòlegs bacterians a salinitats superiors al 20-25%.[70] Els termòfils prosperen a temperatures per sobre de 45 °C, en llocs com ara aigües termals; els arqueus hipertermòfils són els que prosperen a temperatures superiors a 80 °C.[108] La soca 116 d'arqueu de Methanopyrus kandleri creix a 122 °C, que és la temperatura més alta registrada a la qual prospera un organisme.[109] Altres arqueus existeixen en condicions molt àcides o alcalines.[107] Per exemple, un dels arqueus acidòfils més extrems és Picrophilus torridus, que creix a un pH 0, cosa que equival a prosperar en àcid sulfúric amb una concentració molar d'1,2.[110]
Aquesta resistència a ambients extrems ha convertit els arqueus en el centre d'especulació sobre les possibles propietats de la vida extraterrestre.[111] Aquesta s'ha centrat en la possibilitat que existeixi vida microbial a Mart,[112] i fins i tot s'ha arribat a suggerir que microbis viables podrien viatjar entre planetes en meteorits.[113]
Recentment, diversos estudis han demostrat que els arqueus no existeixen únicament en medis mesòfils i termòfils, sinó que també estan presents, a vegades en altes quantitats, a temperatures baixes. Per exemple, els arqueus són comuns en ambients oceànics freds com ara els mars polars.[114] Encara són més significants les grans quantitats d'arqueus que viuen arreu del món en la comunitat planctònica (com a part del picoplàncton).[115] Tot i que aquests arqueus poden ser presents en quantitats extremament grans (fins a un 40% de la biomassa microbiana), gairebé cap d'aquestes espècies no ha sigut aïllada i estudiada en un cultiu pur.[116] Per consegüent, la comprensió actual del paper dels arqueus en l'ecologia dels oceans és rudimentària, de manera que la seva influència completa sobre els cicles biogeoquímics globals roman en gran part sense explorar.[117] Tanmateix, un estudi recent ha demostrat que un grup de crenarqueots marins són capaços de dur a terme la nitrificació, suggerint que aquests organismes podrien ser importants en el cicle del nitrogen oceànic.[118] També es troben grans quantitats d'arqueus als sediments que cobreixen el fons marí, i aquests organismes formen la majoria de cèl·lules vivents a profunditats de més d'un metre dins aquest sediment.[119][120]
Paper en els cicles químics
[modifica]Els arqueus formen part dels sistemes de la Terra que reciclen elements com ara carboni, nitrogen i sofre dels diversos hàbitats de cada ecosistema. Tot i que aquestes activitats són vitals pel funcionament normal dels ecosistemes, els arqueus també poden contribuir als canvis que els humans causen a l'ambient, i fins i tot provocar contaminació.
Els arqueus duen a terme múltiples passos del cicle del nitrogen; això inclou tant reaccions dissimilatòries que eliminen nitrogen dels ecosistemes (com ara la respiració basada en nitrats i la desnitrificació), com processos assimilatoris que introdueixen nitrogen, com ara l'assimilació de nitrogen i la fixació de nitrogen.[121] El paper dels arqueus en les reaccions d'oxidació d'amoníac fou descobert recentment; és particularment important als oceans.[82][122] Sembla que els arqueus també tenen una especial importància en l'oxidació d'amoníac al sòl; produeixen nitrits, que són posteriorment oxidats en nitrats per altres microbis, i després absorbits per plantes i altres organismes.[123]
En el cicle del sofre, els arqueus que creixen oxidant compostos de sofre són importants car alliberen aquest element de les roques, fent-lo disponible per altres organismes. Tanmateix, els arqueus que fan això, com ara Sulfolobus, poden causar danys ambientals, car produeixen àcid sulfúric com producte residual, i el creixement d'aquests organismes en mines abandonades pot causar un drenatge miner àcid.[124]
En el cicle del carboni, els arqueus metanògens són rellevants com productors de metà. La capacitat d'aquests arqueus d'eliminar hidrogen és important en la degradació de matèria orgànica per part de les poblacions de microorganismes que actuen com desintegradors en ecosistemes anaeròbics, com ara sediment, aiguamolls o instal·lacions de tractament de residus.[125] Tanmateix, el metà és un dels gasos amb efecte d'hivernacle més abundants de l'atmosfera terrestre, representant el 18 % del total global.[126] És un gas amb efecte d'hivernacle 25 vegades més potent que el diòxid de carboni.[127] Els metanògens són la font principal de metà atmosfèric, i són els responsables de bona part de les emissions anuals de metà del món.[128] Per tant, aquests arqueus contribueixen a les emissions globals de gasos amb efecte d'hivernacle i a l'escalfament global.
Interaccions amb altres organismes
[modifica]
Les interaccions ben definides entre arqueus i altres organismes són o bé mutualistes o bé comensalistes. El 2007, encara no es coneixia cap exemple clar de patogen o paràsit arqueu.[129][130] Tanmateix, s'ha suggerit una relació entre la presència d'algunes espècies de metanògens i les infeccions a la boca,[131][132] i Nanoarchaeum equitans podria ser parasitari, car només sobreviu i es reprodueix dins les cèl·lules del crenarqueot Ignicoccus hospitalis,[133] i sembla no oferir cap benefici al seu hoste.[134]
Un exemple ben comprès de mutualisme és la interacció entre els protozous i els arqueus metanògens del sistema digestiu d'animals que digereixen cel·lulosa, com ara remugants i tèrmits.[135] En aquests ambients anaeròbics, els protozous descomponen la cel·lulosa de material vegetal per obtenir-ne energia. Aquest procés allibera hidrogen com producte residual, però els nivells alts d'hidrogen redueixen l'energia generada per aquesta reacció. Quan els metanògens converteixen l'hidrogen en metà, els protozous se'n beneficien car podran obtenir més energia de la descomposició de la cel·lulosa.[136]
Aquestes associacions entre metanògens i protozous van un pas més enllà en diverses espècies de protozous anaerobis, com ara Plagiopyla frontata; en aquest cas, els arqueus viuen dins el protozou i consumeixen l'hidrogen produït als seus hidrogenosomes.[137][138] S'estan descobrint associacions similars amb organismes més grans, amb el descobriment que l'arqueu marí Cenarchaeum symbiosum viu a dins de (és un endosimbiont de) l'esponja Axinella mexicana.[139]
Els arqueus també poden ser comensals, beneficiant-se d'una associació sense ajudar o danyar l'altre organisme. Per exemple, el metanogen Methanobrevibacter smithii és de llarg l'arqueu més comú de la flora humana, representant aproximadament un 10% de tots els procariotes de l'intestí humà.[140] Com en el cas dels tèrmits, és possible que aquests metanògens siguin en realitat mutualistes en els humans, interaccionant amb altres microbis de l'intestí per facilitar la digestió dels aliments.[141] També hi ha comunitats d'arqueus associades amb una varietat d'altres organismes, com ara a la superfície de coralls,[142] i a la part del sòl que envolta les arrels de les plantes (la rizosfera).[143][144]
Importància en la tecnologia i la indústria
[modifica]Els arqueus extremòfils, especialment els que resisteixen la calor o acideses i alcalinitats extremes, són una font d'enzims que funcionen en aquestes condicions extremes.[145][146] Aquests enzims tenen una gran varietat d'usos. Per exemple, ADN polimerases termoestables, com ara la Pfu ADN polimerasa de Pyrococcus furiosus, han revolucionat la biologia molecular permetent que es faci servir la reacció en cadena de la polimerasa com tècnica senzilla i ràpida per clonar ADN. En la indústria, amilases, galactosidases i pul·lulanases d'altres espècies de Pyrococcus que funcionen per sobre de 100 °C permeten tractar aliments a altres temperatures, com en la producció de llet i xerigot pobres en lactosa.[147] Els enzims d'aquests arqueus termòfils també tendeixen a ser molt estables en dissolvents orgànics, permetent-ne la utilització en processos respectuosos amb el medi ambient en química verda que sintetitzen compostos orgànics.[146] L'estabilitat dels enzims termòfils també fa que siguin més fàcils d'utilitzar en la biologia estructural, de manera que els enzims homòlegs d'enzims bacterians o eucariotes d'arqueus extremòfils siguin utilitzats en estudis estructurals.[148]
En contrast amb la varietat d'usos dels enzims dels arqueus, els usos dels organismes en si en la biotecnologia són més limitats. Tanmateix, els arqueus metanògens són una part vital del tractament de residus, car formen part de la comunitat de microorganismes que duen a terme la digestió anaeròbica i produeixen biogàs.[149] En la mineria mineralúrgia, els arqueus acidòfils són prometedors per l'extracció de metalls dels minerals, incloent-hi or, cobalt i coure.[150]
En els arqueus s'ha descobert una nova classe d'antibiòtics potencialment útils. S'han descrit algunes d'aquestes arqueocines, però es creu que n'existeixen centenars més, especialment en Haloarchaea i Sulfolobus.[151] Aquests compostos són importants car tenen una estructura diferent de la dels antibiòtics bacterians, de manera que poden tenir un mode d'acció diferent. A més, podrien permetre la creació de nous marcadors seleccionables per utilitzar-los en la biologia molecular d'arqueus. El descobriment de noves arqueocines depèn de la recol·lecció i cultiu reeixits de noves espècies d'arqueus del medi ambient.[152]
Referències
[modifica]- ↑ Baggott, 2018, p. 248.
- ↑ «Què cal saber? La denominació dels arqueus». Societat Catalana de Biologia, octubre 2009 (actualització gener 2021). [Consulta: 7 setembre 2024].
- ↑ ScienceDaily
- ↑ Staley JT «The bacterial species dilemma and the genomic-phylogenetic species concept». Philos. Trans. R. Soc. Lond., B, Biol. Sci., 361, 1475, 2006, pàg. 1899–909. DOI: 10.1098/rstb.2006.1914. PMID: 17062409.
- ↑ Zuckerkandl E, Pauling L «Molecules as documents of evolutionary history». J. Theor. Biol., 8, 2, 1965, pàg. 357–66. DOI: 10.1016/0022-5193(65)90083-4. PMID: 5876245.
- ↑ Woese C, Fox G «Phylogenetic structure of the prokaryotic domain: the primary kingdoms». Proc Natl Acad Sci USA, 74, 11, 1977, pàg. 5.088–5.090. DOI: 10.1073/pnas.74.11.5088. PMID: 270744.
- ↑ Woese CR, Kandler O, Wheelis ML «Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya». Proc. Natl. Acad. Sci. U.S.A., 87, 12, 1990, pàg. 4576–9. DOI: 10.1073/pnas.87.12.4576. PMID: 2112744.
- ↑ archaea. (2008). Merriam-Webster Online Dictionary [Consultat l'1 de juliol de 2008]
- ↑ 9,0 9,1 9,2 DeLong EF «Everything in moderation: archaea as 'non-extremophiles'». Curr. Opin. Genet. Dev., 8, 6, 1998, pàg. 649–54. DOI: 10.1016/S0959-437X(98)80032-4. PMID: 9914204.
- ↑ Theron J, Cloete TE «Molecular techniques for determining microbial diversity and community structure in natural environments». Crit. Rev. Microbiol., 26, 1, 2000, pàg. 37–57. DOI: 10.1080/10408410091154174. PMID: 10782339.
- ↑ Schmidt TM «The maturing of microbial ecology». Int. Microbiol., 9, 3, 2006, pàg. 217–23. Arxivat de l'original el 2008-09-11. PMID: 17061212 [Consulta: 26 setembre 2008]. Arxivat 2008-09-11 a Wayback Machine.
- ↑ Gevers D, Dawyndt P, Vandamme P, et al. «Stepping stones towards a new prokaryotic taxonomy». Philos. Trans. R. Soc. Lond., B, Biol. Sci., 361, 1475, 2006, pàg. 1911-6. DOI: 10.1098/rstb.2006.1915. PMID: 17062410.
- ↑ 13,0 13,1 Robertson CE, Harris JK, Spear JR, Pace NR «Phylogenetic diversity and ecology of environmental Archaea». Curr. Opin. Microbiol., 8, 6, 2005, pàg. 638–42. PMID: 16236543.
- ↑ Huber H, Hohn MJ, Rachel R, Fuchs T, Wimmer VC, Stetter KO. «A new phylum of Archaea represented by a nanosized hyperthermophilic symbiont». Nature, 417, 6884, 2002, pàg. 27–8. DOI: 10.1038/417063a. PMID: 11986665.
- ↑ Barns SM, Delwiche CF, Palmer JD, Pace NR «Perspectives on archaeal diversity, thermophily and monophyly from environmental rRNA sequences». Proc. Natl. Acad. Sci. U.S.A., 93, 17, 1996, pàg. 9188–93. DOI: 10.1073/pnas.93.17.9188. PMID: 8799176.
- ↑ Elkins JG, Podar M, Graham DE, et al. «A korarchaeal genome reveals insights into the evolution of the Archaea». Proc. Natl. Acad. Sci. U.S.A., 105, 23, Juny 2008, pàg. 8102–7. DOI: 10.1073/pnas.0801980105. PMID: 18535141.
- ↑ Baker, B.J., Tyson, G.W., Webb, R.I., Flanagan, J., Hugenholtz, P. and Banfield, J.F. «Lineages of acidophilic Archaea revealed by community genomic analysis. Science». Science, 314, 6884, 2006, pàg. 1933 – 1935. DOI: 10.1126/science.1132690. PMID: 17185602.
- ↑ de Queiroz K «Ernst Mayr and the modern concept of species». Proc. Natl. Acad. Sci. U.S.A., 102 Suppl 1, 2005, pàg. 6600–7. DOI: 10.1073/pnas.0502030102. PMID: 15851674.
- ↑ Eppley JM, Tyson GW, Getz WM, Banfield JF «Genetic exchange across a species boundary in the archaeal genus ferroplasma». Genetics, 177, 1, 2007, pàg. 407–16. DOI: 10.1534/genetics.107.072892. PMID: 17603112.
- ↑ Papke RT, Zhaxybayeva O, Feil EJ, Sommerfeld K, Muise D, Doolittle WF «Searching for species in haloarchaea». Proc. Natl. Acad. Sci. U.S.A., 104, 35, 2007, pàg. 14092–7. DOI: 10.1073/pnas.0706358104. PMID: 17715057.
- ↑ Kunin V, Goldovsky L, Darzentas N, Ouzounis CA «The net of life: reconstructing the microbial phylogenetic network». Genome Res., 15, 7, 2005, pàg. 954–9. DOI: 10.1101/gr.3666505. PMID: 15965028.
- ↑ Hugenholtz P «Exploring prokaryotic diversity in the genomic era». Genome Biol., 3, 2, 2002, pàg. REVIEWS0003. Arxivat de l'original el 2020-03-15. DOI: 10.1186/gb-2002-3-2-reviews0003. PMID: 11864374 [Consulta: 26 setembre 2008]. Arxivat 2020-03-15 a Wayback Machine.
- ↑ Rappé MS, Giovannoni SJ «The uncultured microbial majority». Annu. Rev. Microbiol., 57, 2003, pàg. 369–94. DOI: 10.1146/annurev.micro.57.030502.090759. PMID: 14527284.
- ↑ Schopf J «Fossil evidence of Archaean life». Philos Trans R Soc Lond B Biol Sci, 361, 1470, 2006, pàg. 869–85. DOI: 10.1098/rstb.2006.1834. PMID: 16754604.[Enllaç no actiu]
- ↑ Brocks JJ, Logan GA, Buick R, Summons RE «Archean molecular fossils and the early rise of eukaryotes». Science, 285, 5430, 1999, pàg. 1033–6. DOI: 10.1126/science.285.5430.1033. PMID: 10446042.
- ↑ Chappe B, Albrecht P, Michaelis W «Polar Lipids of Archaebacteria in Sediments and Petroleums». Science, 217, 4554, Juliol 1982, pàg. 65–66. DOI: 10.1126/science.217.4554.65. PMID: 17739984.
- ↑ Hahn, Jürgen; Pat Haug «Traces of Archaebacteria in ancient sediments». System Applied Microbiology, 7, Archaebacteria '85 Proceedings, 1986, pàg. 178–83.
- ↑ Wang M, Yafremava LS, Caetano-Anollés D, Mittenthal JE, Caetano-Anollés G «Reductive evolution of architectural repertoires in proteomes and the birth of the tripartite world». Genome Res., 17, 11, 2007, pàg. 1.572.1.585. DOI: 10.1101/gr.6454307. PMID: 17908824.
- ↑ Ciccarelli FD, Doerks T, von Mering C, Creevey CJ, Snel B, Bork P «Toward automatic reconstruction of a highly resolved tree of life». Science, 311, 5765, 2006, pàg. 1283–7. DOI: 10.1126/science.1123061. PMID: 16513982.
- ↑ Woese CR, Gupta R «Are archaebacteria merely derived 'prokaryotes'?». Nature, 289, 5793, 1981, pàg. 95–6. DOI: 10.1038/289095a0. PMID: 6161309.
- ↑ Woese C «The universal ancestor». Proc. Natl. Acad. Sci. U.S.A., 95, 12, 1998, pàg. 6854–9. DOI: 10.1073/pnas.95.12.6854. PMID: 9618502.
- ↑ Gupta RS «The natural evolutionary relationships among prokaryotes». Crit. Rev. Microbiol., 26, 2, 2000, pàg. 111–31. DOI: 10.1080/10408410091154219. PMID: 10890353.
- ↑ Gribaldo S, Brochier-Armanet C «The origin and evolution of Archaea: a state of the art». Philos. Trans. R. Soc. Lond., B, Biol. Sci., 361, 1470, 2006, pàg. 1007–22. Arxivat de l'original el 2012-06-04. DOI: 10.1098/rstb.2006.1841. PMID: 16754611 [Consulta: 26 setembre 2008]. Arxivat 2012-06-04 at Archive.is
- ↑ 34,0 34,1 Woese CR «There must be a prokaryote somewhere: microbiology's search for itself». Microbiol. Rev., 58, 1, Març 1994, pàg. 1–9. PMC: 372949. PMID: 8177167.
- ↑ Lake JA «Origin of the eukaryotic nucleus determined by rate-invariant analysis of rRNA sequences». Nature, 331, 6152, Gener 1988, pàg. 184–6. DOI: 10.1038/331184a0. PMID: 3340165.
- ↑ Gouy M, Li WH «Phylogenetic analysis based on rRNA sequences supports the archaebacterial rather than the eocyte tree». Nature, 339, 6220, Maig 1989, pàg. 145–7. DOI: 10.1038/339145a0. PMID: 2497353.
- ↑ Yutin N, Makarova KS, Mekhedov SL, Wolf YI, Koonin EV «The deep archaeal roots of eukaryotes». Mol. Biol. Evol., 25, Maig 2008, pàg. 1619. DOI: 10.1093/molbev/msn108. PMID: 18463089.
- ↑ Nelson KE, Clayton RA, Gill SR, et al. «Evidence for lateral gene transfer between Archaea and bacteria from genome sequence of Thermotoga maritima». Nature, 399, 6734, 1999, pàg. 323–9. DOI: 10.1038/20601. PMID: 10360571.
- ↑ Lake JA. «Origin of the eukaryotic nucleus determined by rate-invariant analysis of rRNA sequences». Nature, 331, 6152, 1988, pàg. 184–6. DOI: 10.1038/331184a0. PMID: 3340165.
- ↑ Williams, Tom & Cox, Cymon & Foster, Peter & Szollosi, Gergely & Embley, T.. (2020). Phylogenomics provides robust support for a two-domains tree of life. Nature Ecology & Evolution. 4. 1-10. 10.1038/s41559-019-1040-x.
- ↑ Hug, L. A. et al. 2016, A new view of the tree of life. Nature Microbiology, 1, 16048.
- ↑ Cindy J. Castelle & Jillian F. Banfield 2018, Major New Microbial Groups Expand Diversity and Alter our Understanding of the Tree of Life. PERSPECTIVE, VOL 172, Issue 6, P1181-1197, 2018, DOI:https://doi.org/10.1016/j.cell.2018.02.016
- ↑ 43,0 43,1 43,2 43,3 Krieg, Noel. Bergey's Manual® of Systematic Bacteriology. USA: Springer, 2005, p. 21-6. ISBN 978-0-387-24143-2.
- ↑ Barns, Sue i Burggraf, Siegfried. (1997) Crenarchaeota Arxivat 2012-05-02 a Wayback Machine.. Versió de l'1 gener 1997. a The Tree of Life Web Project
- ↑ Walsby, A.E. «A square bacterium». Nature, 283, 5742, 1980, pàg. 69–71. DOI: 10.1038/283069a0.
- ↑ Hara F, Yamashiro K, Nemoto N, 'et al. «An actin homolog of the archaeon Thermoplasma acidophilum that retains the ancient characteristics of eukaryotic actin». J. Bacteriol., 189, 5, 2007, pàg. 2039–45. Arxivat de l'original el 2020-05-27. DOI: 10.1128/JB.01454-06. PMID: 17189356 [Consulta: 26 setembre 2008].
- ↑ Trent JD, Kagawa HK, Yaoi T, Olle E, Zaluzec NJ «Chaperonin filaments: the archaeal cytoskeleton?». Proc. Natl. Acad. Sci. U.S.A., 94, 10, 1997, pàg. 5383–8. Arxivat de l'original el 2020-05-27. DOI: 10.1073/pnas.94.10.5383. PMID: 9144246 [Consulta: 26 setembre 2008].
- ↑ Hixon WG, Searcy DG «Cytoskeleton in the archaebacterium Thermoplasma acidophilum? Viscosity increase in soluble extracts». BioSystems, 29, 2–3, 1993, pàg. 151-60. DOI: 10.1016/0303-2647(93)90091-P. PMID: 8374067.
- ↑ Hall-Stoodley L, Costerton JW, Stoodley P «Bacterial biofilms: from the natural environment to infectious diseases». Nat. Rev. Microbiol., 2, 2, 2004, pàg. 95–108. DOI: 10.1038/nrmicro821. PMID: 15040259.
- ↑ Kuwabara T, Minaba M, Iwayama Y, 'et al. «Thermococcus coalescens sp. nov., a cell-fusing hyperthermophilic archaeon from Suiyo Seamount». Int. J. Syst. Evol. Microbiol., 55, Pt 6, Novembre 2005, pàg. 2507–14. DOI: 10.1099/ijs.0.63432-0. PMID: 16280518.[Enllaç no actiu]
- ↑ Nickell S, Hegerl R, Baumeister W, Rachel R «Pyrodictium cannulae enter the periplasmic space but do not enter the cytoplasm, as revealed by cryo-electron tomography». J. Struct. Biol., 141, 1, 2003, pàg. 34–42. DOI: 10.1016/S1047-8477(02)00581-6. PMID: 12576018.
- ↑ Horn C, Paulmann B, Kerlen G, Junker N, Huber H «In vivo observation of cell division of anaerobic hyperthermophiles by using a high-intensity dark-field microscope». J. Bacteriol., 181, 16, 1999, pàg. 5114–8. DOI: 10.1073/pnas.241636498v1. PMID: 10438790.
- ↑ Rudolph C, Wanner G, Huber R «Natural communities of novel archaea and bacteria growing in cold sulfurous springs with a string-of-pearls-like morphology». Appl. Environ. Microbiol., 67, 5, Maig 2001, pàg. 2336–44. DOI: 10.1128/AEM.67.5.2336-2344.2001. PMC: 92875. PMID: 11319120.
- ↑ 54,0 54,1 Thomas NA, Bardy SL, Jarrell KF «The archaeal flagellum: a different kind of prokaryotic motility structure». FEMS Microbiol. Rev., 25, 2, 2001, pàg. 147–74. DOI: 10.1111/j.1574-6976.2001.tb00575.x. PMID: 11250034.
- ↑ Rachel R, Wyschkony I, Riehl S, Huber H «The ultrastructure of Ignicoccus: evidence for a novel outer membrane and for intracellular vesicle budding in an archaeon». Archaea, 1, 1, Març 2002, pàg. 9–18. Arxivat de l'original el 2009-02-24. PMID: 15803654 [Consulta: 26 setembre 2008].
- ↑ 56,0 56,1 Koga Y, Morii H «Biosynthesis of ether-type polar lipids in archaea and evolutionary considerations». Microbiol. Mol. Biol. Rev., 71, 1, 2007, pàg. 97–120. DOI: 10.1128/MMBR.00033-06. PMID: 17347520.
- ↑ De Rosa M, Gambacorta A, Gliozzi A «Structure, biosynthesis, and physicochemical properties of archaebacterial lipids». Microbiol. Rev., 50, 1, 1986, pàg. 70–80. PMID: 3083222.
- ↑ Albers SV, van de Vossenberg JL, Driessen AJ, Konings WN «Adaptations of the archaeal cell membrane to heat stress». Front. Biosci., 5, Setembre 2000, pàg. D813–20. DOI: 10.2741/albers. PMID: 10966867.
- ↑ Damsté JS, Schouten S, Hopmans EC, van Duin AC, Geenevasen JA «Crenarchaeol: the characteristic core glycerol dibiphytanyl glycerol tetraether membrane lipid of cosmopolitan pelagic crenarchaeota». J. Lipid Res., 43, 10, Octubre 2002, pàg. 1641–51. DOI: 10.1194/jlr.M200148-JLR200. PMID: 12364548.
- ↑ Koga Y, Morii H «Recent advances in structural research on ether lipids from archaea including comparative and physiological aspects». Biosci. Biotechnol. Biochem., 69, 11, Novembre 2005, pàg. 2019–34. Arxivat de l'original el 2008-12-31. DOI: 10.1271/bbb.69.2019. PMID: 16306681 [Consulta: 27 setembre 2008]. Arxivat 2008-12-31 a Wayback Machine.
- ↑ Hanford MJ, Peeples TL «Archaeal tetraether lipids: unique structures and applications». Appl. Biochem. Biotechnol., 97, 1, Gener 2002, pàg. 45–62. DOI: 10.1385/ABAB:97:1:45. PMID: 11900115.
- ↑ Macalady JL, Vestling MM, Baumler D, Boekelheide N, Kaspar CW, Banfield JF «Tetraether-linked membrane monolayers in Ferroplasma spp: a key to survival in acid». Extremophiles, 8, 5, Octubre 2004, pàg. 411–9. DOI: 10.1007/s00792-004-0404-5. PMID: 15258835.
- ↑ Golyshina OV, Pivovarova TA, Karavaiko GI, et al «Ferroplasma acidiphilum gen. nov., sp. nov., an acidophilic, autotrophic, ferrous-iron-oxidizing, cell-wall-lacking, mesophilic member of the Ferroplasmaceae fam. nov., comprising a distinct lineage of the Archaea». Int. J. Syst. Evol. Microbiol., 50 Pt 3, Maig 2000, pàg. 997–1006. Arxivat de l'original el 2023-11-13. PMID: 10843038 [Consulta: 27 setembre 2008].
- ↑ Sára M, Sleytr UB «S-Layer proteins». J. Bacteriol., 182, 4, 2000, pàg. 859–68. DOI: 10.1128/JB.182.4.859-868.2000. PMID: 10648507.
- ↑ Engelhardt H, Peters J «Structural research on surface layers: a focus on stability, surface layer homology domains, and surface layer-cell wall interactions». J Struct Biol, 124, 2–3, 1998, pàg. 276–302. DOI: 10.1006/jsbi.1998.4070. PMID: 10049812.
- ↑ 66,0 66,1 Kandler, O.; König, H. «Cell wall polymers in Archaea (Archaebacteria)». Cellular and Molecular Life Sciences (CMLS), 54, 4, 1998, pàg. 305–308. DOI: 10.1007/s000180050156.[Enllaç no actiu]
- ↑ Howland, John L. The Surprising Archaea: Discovering Another Domain of Life. Oxford: Oxford University Press, 2000, p. 32. ISBN 0-19-511183-4.
- ↑ Ng SY, Chaban B, Jarrell KF «Archaeal flagella, bacterial flagella and type IV pili: a comparison of genes and posttranslational modifications». J. Mol. Microbiol. Biotechnol., 11, 3–5, 2006, pàg. 167–91. DOI: 10.1159/000094053. PMID: 16983194.
- ↑ Bardy SL, Ng SY, Jarrell KF «Prokaryotic motility structures». Microbiology (Reading, Engl.), 149, Pt 2, Febrer 2003, pàg. 295–304. DOI: 10.1099/mic.0.25948-0. PMID: 12624192.
- ↑ 70,0 70,1 70,2 Valentine DL «Adaptations to energy stress dictate the ecology and evolution of the Archaea». Nat. Rev. Microbiol., 5, 4, 2007, pàg. 316–23. DOI: 10.1038/nrmicro1619. PMID: 17334387.
- ↑ 71,0 71,1 71,2 Schäfer G, Engelhard M, Müller V «Bioenergetics of the Archaea». Microbiol. Mol. Biol. Rev., 63, 3, Setembre 1999, pàg. 570–620. PMC: 103747. PMID: 10477309.
- ↑ Zillig W «Comparative biochemistry of Archaea and Bacteria». Curr. Opin. Genet. Dev., 1, 4, Desembre 1991, pàg. 544–51. DOI: 10.1016/S0959-437X(05)80206-0. PMID: 1822288.
- ↑ Romano A, Conway T «Evolution of carbohydrate metabolic pathways». Res Microbiol, 147, 6–7, 1996, pàg. 448–55. DOI: 10.1016/0923-2508(96)83998-2. PMID: 9084754.
- ↑ Koch A «How did bacteria come to be?». Adv Microb Physiol, 40, 1998, pàg. 353–99. DOI: 10.1016/S0065-2911(08)60135-6. PMID: 9889982.
- ↑ DiMarco AA, Bobik TA, Wolfe RS «Unusual coenzymes of methanogenesis». Annu. Rev. Biochem., 59, 1990, pàg. 355–94. DOI: 10.1146/annurev.bi.59.070190.002035. PMID: 2115763.
- ↑ Klocke M, Nettmann E, Bergmann I, et al «Characterization of the methanogenic Archaea within two-phase biogas reactor systems operated with plant biomass». Syst. Appl. Microbiol., 31, Maig 2008, pàg. 190. DOI: 10.1016/j.syapm.2008.02.003. PMID: 18501543.
- ↑ Basat en PDB 1FBB. Dades publicades a Subramaniam S, Henderson R «Molecular mechanism of vectorial proton translocation by bacteriorhodopsin». Nature, 406, 6796, Agost 2000, pàg. 653–7. DOI: 10.1038/35020614. PMID: 10949309.
- ↑ Mueller-Cajar O, Badger MR «New roads lead to Rubisco in archaebacteria». Bioessays, 29, 8, Agost 2007, pàg. 722–4. DOI: 10.1002/bies.20616. PMID: 17621634.
- ↑ Berg IA, Kockelkorn D, Buckel W, Fuchs G «A 3-hydroxypropionate/4-hydroxybutyrate autotrophic carbon dioxide assimilation pathway in Archaea». Science (journal), 318, 5857, Desembre 2007, pàg. 1782–6. DOI: 10.1126/science.1149976. PMID: 18079405.
- ↑ Thauer RK «Microbiology. A fifth pathway of carbon fixation». Science (journal), 318, 5857, Desembre 2007, pàg. 1732–3. DOI: 10.1126/science.1152209. PMID: 18079388.
- ↑ Bryant DA, Frigaard NU «Prokaryotic photosynthesis and phototrophy illuminated». Trends Microbiol., 14, 11, Novembre 2006, pàg. 488–96. DOI: 10.1016/j.tim.2006.09.001. PMID: 16997562.
- ↑ 82,0 82,1 Francis CA, Beman JM, Kuypers MM «New processes and players in the nitrogen cycle: the microbial ecology of anaerobic and archaeal ammonia oxidation». ISME J, 1, 1, Maig 2007, pàg. 19–27. DOI: 10.1038/ismej.2007.8. PMID: 18043610.
- ↑ Könneke M, Bernhard AE, de la Torre JR, Walker CB, Waterbury JB, Stahl DA «Isolation of an autotrophic ammonia-oxidizing marine archaeon». Nature, 437, 7058, Setembre 2005, pàg. 543–6. DOI: 10.1038/nature03911. PMID: 16177789.
- ↑ Lanyi JK «Bacteriorhodopsin». Annu. Rev. Physiol., 66, 2004, pàg. 665–88. DOI: 10.1146/annurev.physiol.66.032102.150049. PMID: 14977418.
- ↑ 85,0 85,1 Allers T, Mevarech M «Archaeal genetics - the third way». Nat. Rev. Genet., 6, 1, 2005, pàg. 58–73. DOI: 10.1038/nrg1504. PMID: 15630422.
- ↑ Galagan JE, et al «The genome of M. acetivorans reveals extensive metabolic and physiological diversity». Genome Research, 12, 4, 4-2002, pàg. 532–42. DOI: 11932238. ISSN: 1088-9051.
- ↑ Waters E, et al «The genome of Nanoarchaeum equitans: insights into early archaeal evolution and derived parasitism». Proc. Natl. Acad. Sci. U.S.A., 100, 22, 2003, pàg. 12984–8. DOI: 10.1073/pnas.1735403100. PMID: 14566062.
- ↑ Schleper C, Holz I, Janekovic D, Murphy J, Zillig W «A multicopy plasmid of the extremely thermophilic archaeon Sulfolobus effects its transfer to recipients by mating». J. Bacteriol., 177, 15, 1995, pàg. 4417–26. Arxivat de l'original el 2012-05-29. PMID: 7635827 [Consulta: 12 octubre 2008].
- ↑ Sota M; Top EM. «Horizontal Gene Transfer Mediated by Plasmids». A: Plasmids: Current Research and Future Trends. Caister Academic Press, 2008. [1] ISBN 978-1-904455-35-6.
- ↑ Xiang X, Chen L, Huang X, Luo Y, She Q, Huang L «Sulfolobus tengchongensis spindle-shaped virus STSV1: virus-host interactions and genomic features». J. Virol., 79, 14, 2005, pàg. 8677–86. DOI: 10.1128/JVI.79.14.8677-8686.2005. PMID: 15994761.
- ↑ Prangishvili D, Forterre P, Garrett RA «Viruses of the Archaea: a unifying view». Nat. Rev. Microbiol., 4, 11, 2006, pàg. 837–48. DOI: 10.1038/nrmicro1527. PMID: 17041631.
- ↑ Prangishvili D, Garrett RA «Exceptionally diverse morphotypes and genomes of crenarchaeal hyperthermophilic viruses». Biochem. Soc. Trans., 32, Pt 2, 2004, pàg. 204–8. DOI: 10.1042/BST0320204. PMID: 15046572.
- ↑ Mojica FJ, Díez-Villaseñor C, García-Martínez J, Soria E «Intervening sequences of regularly spaced prokaryotic repeats derive from foreign genetic elements». J. Mol. Evol., 60, 2, 2005, pàg. 174–82. DOI: 10.1007/s00239-004-0046-3. PMID: 15791728.
- ↑ Makarova KS, Grishin NV, Shabalina SA, Wolf YI, Koonin EV «A putative RNA-interference-based immune system in prokaryotes: computational analysis of the predicted enzymatic machinery, functional analogies with eukaryotic RNAi, and hypothetical mechanisms of action». Biol. Direct, 1, 2006, pàg. 7. DOI: 10.1186/1745-6150-1-7. PMID: 16545108.
- ↑ Graham DE, Overbeek R, Olsen GJ, Woese CR «An archaeal genomic signature». Proc. Natl. Acad. Sci. U.S.A., 97, 7, 2000, pàg. 3304–8. DOI: 10.1073/pnas.050564797. PMID: 10716711.
- ↑ 96,0 96,1 Gaasterland T «Archaeal genomics». Curr. Opin. Microbiol., 2, 5, 1999, pàg. 542–7. DOI: 10.1016/S1369-5274(99)00014-4. PMID: 10508726.
- ↑ Werner F «Structure and function of archaeal RNA polymerases». Mol. Microbiol., 65, 6, Setembre 2007, pàg. 1395–404. DOI: 10.1111/j.1365-2958.2007.05876.x. PMID: 17697097.
- ↑ Aravind L, Koonin EV «DNA-binding proteins and evolution of transcription regulation in the archaea». Nucleic Acids Res., 27, 23, 1999, pàg. 4658–70. DOI: 10.1093/nar/27.23.4658. PMID: 10556324.
- ↑ Lykke-Andersen J, Aagaard C, Semionenkov M, Garrett RA «Archaeal introns: splicing, intercellular mobility and evolution». Trends Biochem. Sci., 22, 9, Setembre 1997, pàg. 326-31. DOI: 10.1016/S0968-0004(97)01113-4. PMID: 9301331.
- ↑ Watanabe Y, Yokobori S, Inaba T, et al «Introns in protein-coding genes in Archaea». FEBS Lett., 510, 1-2, Gener 2002, pàg. 27–30. DOI: 10.1016/S0014-5793(01)03219-7. PMID: 11755525.
- ↑ Yoshinari S, Itoh T, Hallam SJ, et al «Archaeal pre-mRNA splicing: a connection to hetero-oligomeric splicing endonuclease». Biochem. Biophys. Res. Commun., 346, 3, Agost 2006, pàg. 1024–32. DOI: 10.1016/j.bbrc.2006.06.011. PMID: 16781672.
- ↑ 102,0 102,1 Bernander R «Archaea and the cell cycle». Mol. Microbiol., 29, 4, 1998, pàg. 955–61. DOI: 10.1046/j.1365-2958.1998.00956.x. PMID: 9767564.
- ↑ Kelman LM, Kelman Z «Multiple origins of replication in archaea». Trends Microbiol., 12, 9, 2004, pàg. 399–401. DOI: 10.1016/j.tim.2004.07.001. PMID: 153371581.
- ↑ Onyenwoke RU, Brill JA, Farahi K, Wiegel J «Sporulation genes in members of the low G+C Gram-type-positive phylogenetic branch (Firmicutes)». Arch. Microbiol., 182, 2–3, 2004, pàg. 182–92. DOI: 10.1007/s00203-004-0696-y. PMID: 15340788.
- ↑ Kostrikina NA, Zvyagintseva IS, Duda VI. «Cytological peculiarities of some extremely halophilic soil archaeobacteria». Arch. Microbiol., 156, 5, 1991, pàg. 344–49. DOI: 10.1007/BF00248708.
- ↑ DeLong EF, Pace NR «Environmental diversity of bacteria and archaea». Syst. Biol., 50, 4, 2001, pàg. 470–8. DOI: 10.1080/106351501750435040. PMID: 12116647.
- ↑ 107,0 107,1 Pikuta EV, Hoover RB, Tang J «Microbial extremophiles at the limits of life». Crit. Rev. Microbiol., 33, 3, 2007, pàg. 183–209. DOI: 10.1080/10408410701451948. PMID: 17653987.
- ↑ Madigan MT, Martino JM. Brock Biology of Microorganisms. 11a ed.. Pearson, 2006, p. 136. ISBN 0-13-196893-9.
- ↑ Takai K, Nakamura K, Toki T, Tsunogai U, Miyazaki M, Miyazaki J, Hirayama H, Nakagawa S, Nunoura T, Horikoshi K «Cell proliferation at 122 °C and isotopically heavy CH4 production by a hyperthermophilic methanogen under high-pressure cultivation». Proc Natl Acad Sci U S A, 105, 2008, pàg. 10949-54. DOI: 10.1073/pnas.0712334105.
- ↑ Ciaramella M, Napoli A, Rossi M «Another extreme genome: how to live at pH 0». Trends Microbiol., 13, 2, Febrer 2005, pàg. 49–51. DOI: 10.1016/j.tim.2004.12.001. PMID: 15680761.
- ↑ Javaux EJ «Extreme life on Earth—past, present and possibly beyond». Res. Microbiol., 157, 1, 2006, pàg. 37–48. DOI: 10.1016/j.resmic.2005.07.008. PMID: 16376523.
- ↑ Nealson KH «Post-Viking microbiology: new approaches, new data, new insights». Orig Life Evol Biosph, 29, 1, Gener 1999, pàg. 73–93. DOI: 10.1023/A:1006515817767. PMID: 11536899.
- ↑ Davies PC «The transfer of viable microorganisms between planets». Ciba Found. Symp., 202, 1996, pàg. 304–14; discussion 314–7. PMID: 9243022.
- ↑ López-García P, López-López A, Moreira D, Rodríguez-Valera F «Diversity of free-living prokaryotes from a deep-sea site at the Antarctic Polar Front». FEMS Microbiol. Ecol., 36, 2–3, Juliol 2001, pàg. 193–202. PMID: 11451524.
- ↑ Karner MB, DeLong EF, Karl DM «Archaeal dominance in the mesopelagic zone of the Pacific Ocean». Nature, 409, 6819, 2001, pàg. 507–10. DOI: 10.1038/35054051. PMID: 11206545.
- ↑ Giovannoni SJ, Stingl U. «Molecular diversity and ecology of microbial plankton». Nature, 427, 7057, 2005, pàg. 343–8. DOI: 10.1038/nature04158. PMID: 16163344.
- ↑ DeLong EF, Karl DM «Genomic perspectives in microbial oceanography». Nature, 437, 7057, Setembre 2005, pàg. 336–42. DOI: 10.1038/nature04157. PMID: 16163343.
- ↑ Konneke M, Bernhard AE, de la Torre JR, Walker CB, Waterbury JB, Stahl DA. «Isolation of an autotrophic ammonia-oxidizing marine archaeon». Nature, 437, 7057, 2005, pàg. 543–6. DOI: 10.1038/nature03911. PMID: 16177789.
- ↑ Teske A, Sørensen KB «Uncultured archaea in deep marine subsurface sediments: have we caught them all?». ISME J, 2, 1, Gener 2008, pàg. 3–18. DOI: 10.1038/ismej.2007.90. PMID: 18180743.
- ↑ Lipp JS, Morono Y, Inagaki F, Hinrichs KU «Significant contribution of Archaea to extant biomass in marine subsurface sediments». Nature, Juliol 2008. DOI: 10.1038/nature07174. PMID: 18641632.
- ↑ Cabello P, Roldán MD, Moreno-Vivián C «Nitrate reduction and the nitrogen cycle in archaea». Microbiology (Reading, Engl.), 150, Pt 11, Novembre 2004, pàg. 3527–46. Arxivat de l'original el 2011-06-07. DOI: 10.1099/mic.0.27303-0. PMID: 15528644 [Consulta: 18 octubre 2008].
- ↑ Coolen MJ, Abbas B, van Bleijswijk J, et al. «Putative ammonia-oxidizing Crenarchaeota in suboxic waters of the Black Sea: a basin-wide ecological study using 16S ribosomal and functional genes and membrane lipids». Environ. Microbiol., 9, 4, Abril 2007, pàg. 1001–16. DOI: 10.1111/j.1462-2920.2006.01227.x. PMID: 17359272.
- ↑ Leininger S, Urich T, Schloter M, et al «Archaea predominate among ammonia-oxidizing prokaryotes in soils». Nature, 442, 7104, Agost 2006, pàg. 806–9. DOI: 10.1038/nature04983. PMID: 16915287.
- ↑ Baker, B. J.; Banfield, J. F. «Microbial communities in acid mine drainage». FEMS Microbiology Ecology, 44, 2, 2003, p. 139–152. DOI: 10.1016/S0168-6496(03)00028-X.
- ↑ Schimel J «Playing scales in the methane cycle: from microbial ecology to the globe». Proc. Natl. Acad. Sci. U.S.A., 101, 34, Agost 2004, pàg. 12400–1. DOI: 10.1073/pnas.0405075101. PMC: 515073. PMID: 15314221.
- ↑ «EDGAR 3.2 Fast Track 2000». Arxivat de l'original el 2008-05-21. [Consulta: 26 juny 2008].
- ↑ «Annual Greenhouse Gas Index (AGGI) Indicates Sharp Rise in Carbon Dioxide and Methane in 2007», 23-04-2008. Arxivat de l'original el 2008-05-14. [Consulta: 26 juny 2008].
- ↑ «Trace Gases: Current Observations, Trends, and Budgets». Climate Change 2001. United Nations Environment Programme. Arxivat de l'original el 2012-08-05. [Consulta: 19 octubre 2008].
- ↑ Eckburg P, Lepp P, Relman D «Archaea and their potential role in human disease». Infect Immun, 71, 2, 2003, pàg. 591-6. DOI: 10.1128/IAI.71.2.591-596.2003. PMID: 12540534.
- ↑ Cavicchioli R, Curmi P, Saunders N, Thomas T «Pathogenic archaea: do they exist?». Bioessays, 25, 11, 2003, pàg. 1119–28. DOI: 10.1002/bies.10354. PMID: 14579252.
- ↑ Lepp P, Brinig M, Ouverney C, Palm K, Armitage G, Relman D «Methanogenic Archaea and human periodontal disease». Proc Natl Acad Sci U S a, 101, 16, 2004, pàg. 6176–81. DOI: 10.1073/pnas.0308766101. PMID: 15067114.
- ↑ Vianna ME, Conrads G, Gomes BP, Horz HP «Identification and quantification of archaea involved in primary endodontic infections». J. Clin. Microbiol., 44, 4, Abril 2006, pàg. 1274–82. Arxivat de l'original el 2019-10-16. DOI: 10.1128/JCM.44.4.1274-1282.2006. PMC: 1448633. PMID: 16597851 [Consulta: 19 octubre 2008].
- ↑ Waters E, Hohn MJ, Ahel I, et al. «The genome of Nanoarchaeum equitans: insights into early archaeal evolution and derived parasitism». Proc. Natl. Acad. Sci. U.S.A., 100, 22, Octubre 2003, pàg. 12984–8. DOI: 10.1073/pnas.1735403100. PMC: 240731. PMID: 14566062.
- ↑ Jahn U, Gallenberger M, Paper W, et al. «Nanoarchaeum equitans and Ignicoccus hospitalis: new insights into a unique, intimate association of two archaea». J. Bacteriol., 190, 5, Març 2008, pàg. 1743–50. Arxivat de l'original el 2020-05-27. DOI: 10.1128/JB.01731-07. PMID: 18165302 [Consulta: 19 octubre 2008].
- ↑ Chaban B, Ng SY, Jarrell KF «Archaeal habitats—from the extreme to the ordinary». Can. J. Microbiol., 52, 2, Febrer 2006, pàg. 73–116. DOI: 10.1139/w05-147. PMID: 16541146.
- ↑ Schink B «Energetics of syntrophic cooperation in methanogenic degradation». Microbiol. Mol. Biol. Rev., 61, 2, Juny 1997, pàg. 262–80. PMC: 232610. PMID: 9184013.
- ↑ Lange, M.; Westermann, P.; Ahring, B.K. «Archaea in protozoa and metazoa». Applied Microbiology and Biotechnology, 66, 5, 2005, pàg. 465–474. DOI: 10.1007/s00253-004-1790-4.
- ↑ van Hoek AH, van Alen TA, Sprakel VS, et al. «Multiple acquisition of methanogenic archaeal symbionts by anaerobic ciliates». Mol. Biol. Evol., 17, 2, Febrer 2000, pàg. 251–8. PMID: 10677847.
- ↑ Preston, C.M.; Wu, K.Y.; Molinski, T.F.; Delong, E.F. «A psychrophilic crenarchaeon inhabits a marine sponge: Cenarchaeum symbiosum gen. nov., sp. nov». Proc Natl Acad Sci USA, 93, 13, 1996, pàg. 6241-6246. DOI: 10.1073/pnas.93.13.6241. PMID: 8692799.
- ↑ Eckburg PB, Bik EM, Bernstein CN, et al «Diversity of the human intestinal microbial flora». Science (journal), 308, 5728, Juny 2005, pàg. 1635–8. DOI: 10.1126/science.1110591. PMC: 1395357. PMID: 15831718.
- ↑ Samuel BS, Gordon JI «A humanized gnotobiotic mouse model of host-archaeal-bacterial mutualism». Proc. Natl. Acad. Sci. U.S.A., 103, 26, Juny 2006, pàg. 10011-6. DOI: 10.1073/pnas.0602187103. PMC: 1479766. PMID: 16782812.
- ↑ Wegley, L.; Yu, Y.; Breitbart, M.; Casas, V.; Kline, D.I.; Rohwer, F. «Coral-associated Archaea» (PDF). Marine Ecology Progress Series, 273, 2004, pàg. 89–96. Arxivat de l'original el 2008-09-11. DOI: 10.3354/meps273089 [Consulta: 19 octubre 2008]. Arxivat 2008-09-11 a Wayback Machine.
- ↑ Chelius MK, Triplett EW «The Diversity of Archaea and Bacteria in Association with the Roots of Zea mays L». Microb. Ecol., 41, 3, Abril 2001, pàg. 252–63. DOI: 10.1007/s002480000087. PMID: 11391463.
- ↑ Simon HM, Dodsworth JA, Goodman RM «Crenarchaeota colonize terrestrial plant roots». Environ. Microbiol., 2, 5, Octubre 2000, pàg. 495–505. DOI: 10.1046/j.1462-2920.2000.00131.x. PMID: 11233158.
- ↑ Breithaupt H «The hunt for living gold. The search for organisms in extreme environments yields useful enzymes for industry». EMBO Rep., 2, 11, 2001, pàg. 968–71. DOI: 10.1093/embo-reports/kve238. PMID: 11713183.
- ↑ 146,0 146,1 Egorova K, Antranikian G «Industrial relevance of thermophilic Archaea». Curr. Opin. Microbiol., 8, 6, 2005, pàg. 649–55. PMID: 16257257.
- ↑ Synowiecki J, Grzybowska B, Zdziebło A «Sources, properties and suitability of new thermostable enzymes in food processing». Crit Rev Food Sci Nutr, 46, 3, 2006, pàg. 197–205. DOI: 10.1080/10408690590957296. PMID: 16527752.
- ↑ Jenney FE, Adams MW «The impact of extremophiles on structural genomics (and vice versa)». Extremophiles, 12, 1, Gener 2008, pàg. 39–50. DOI: 10.1007/s00792-007-0087-9. PMID: 17563834.
- ↑ Schiraldi C, Giuliano M, De Rosa M «Perspectives on biotechnological applications of archaea». Archaea, 1, 2, 2002, pàg. 75–86. Arxivat de l'original el 2013-08-26. PMID: 15803645 [Consulta: 19 octubre 2008].
- ↑ Norris PR, Burton NP, Foulis NA «Acidophiles in bioreactor mineral processing». Extremophiles, 4, 2, 2000, pàg. 71-6. DOI: 10.1007/s007920050139. PMID: 10805560.
- ↑ O'Connor EM, Shand RF «Halocins and sulfolobicins: the emerging story of archaeal protein and peptide antibiotics». J. Ind. Microbiol. Biotechnol., 28, 1, Gener 2002, pàg. 23–31. DOI: 10.1038/sj/jim/7000190. PMID: 11938468.
- ↑ Shand RF; Leyva KJ. «Archaeal Antimicrobials: An Undiscovered Country». A: Blum P (ed.). Archaea: New Models for Prokaryotic Biology. Caister Academic Press, 2008. ISBN 978-1-904455-27-1.
Bibliografia
[modifica]- Baggott, J. Origins: The Scientific Story of Creation (en anglès). Oxford University Press, 2018. ISBN 9780192561978.