Metionin
| Metionin | |
|---|---|
| CSCCC(C(=O)O)N | |
| Općenito | |
| Hemijski spoj | Metionin |
| Molekularna formula | C=3H=7N=1O=2 |
| CAS registarski broj | 59-51-8 |
| Kratki opis | Bijeli kristalni prah |
| Osobine1 | |
| Molarna masa | 60,07 g/mol |
| Agregatno stanje | čvrsto |
| Gustoća | 1.340 g/cm3 |
| Tačka topljenja | 281 |
| Rastvorljivost | Rastvorljiv |
| 1 Gdje god je moguće korištene su SI jedinice. Ako nije drugačije naznačeno, dati podaci vrijede pri standardnim uslovima. | |
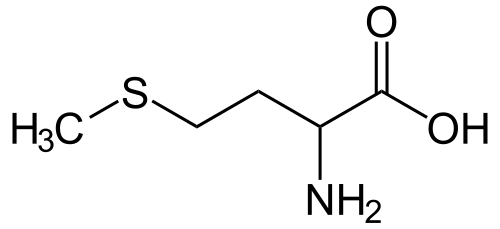

Metionin ili α-amino γ-metil tiobuterna kiselina (HO2CCH(NH2)CH2CH2SCH3) je esencijalna aminokiselina koja skupa sa cisteinom i cistinom spada i u tioaminske kiseline, tj aminokiseline koje u svom molekulu sadrže sumpor). Učestvuje u sintezi proteina, u transmetilacijskim reakcijama i sintezi aminokiselina cistein i cistin. Njegovu ugradnju u polipeptidni lanac kontrolirakodon AUG.[1][2][3]
Značaj i metabolizam
[uredi | uredi izvor]- Metionin, u transmetilacijskim procesima, je donor elektron metil grupe, koja može preći na neki akceptor (primatelj), što je neophodno u sintezi nekih biološki značajnih jedinjenja, kao što su:
- adrenalin,
- kreatin,
- anserin,
- sarkozin,
- betain,
- karnitin,
- holin,
- poliamin i drugi.
U svojoj strukturi metionin ima metil grupu koja je vezana za sumpor, koja se u ovakvim reakcijama lahko može prenositi na neku drugu molekulu. U reakcijama transmetilacije, metionin se najprije mora aktivirati veznjem za ATP, pri čemu se adenozin veže za metionin i daje S-adenozil-metionin ili aktivni metionin. Aktivacija se odvija pod uticajem enzima S-adenozinmetionin transferaze. Iz aktivnog metionina se, zatim, metil grupa može vrlo lako prenijeti na amino ili hidroksilnu grupu primatelja.
U priloženoj tabeli su prikazane značajnije transmetilacijske reakcije.
| Donator metil grupe | Primatelj metil grupe | Proizvod reakcije |
| S-adenozil-metionin | Guanidosirćetna kiselina | Kreatin |
| S-adenozil-metionin | Noradrenalin | Adrenalin |
| S-adenozil-metionin | Karnozin | Anserin |
| S-adenozil-metionin | Lizin | Karnitin |
| S-adenozil-metionin | Etanolamin | Holin |
- Odvajanjem metil grupe od S-adenozil-metionina nastaje S-adenozil homocistein, a S-adenozil-homocistein se hidrolizom razlaže na adenozin i homocistein.
- Homocistein iz prethodne reakcije se spaja sa serinom u cistation, pod katalitskim uticajem enzima cistation sintetaza. Cistation se , uz djejstvo enzima cistationaza razlaže na homoserin i cistein. Na taj način se iz metionin sintetišu i druge tioaminske aminokiseline: cistein i cistin.
- Homoserin iz prethodne reakcije se dezaminizira do α-ketobuterne kiseline, uz oslobađanje molekule vode i vodiksulfida. U daljem metabolizmu α-ketobuterne kiseline, [[propioninastaje koenzim A, koji, nakon dekarboksilacije daje metilmalonil koenzima A. Pod uticajem enzima metilmalonil koenzim A dismutaza ovo jedinjenje prelazi u sukscinil koenzim A, iz koga se može sintetizirati glukoza. Zato je metionin i glikogena aminokiselina.
Derivati metionina
[uredi | uredi izvor]
Metioninski derivat S-adenozil metionin (SAM) je kofaktor koji služi uglavnom kao metil donator. SAM je sastavljen od adenozil molekula (preko 5'uglika), vezan za sumpor metionina, zato što ga čini sulfonijem kalcija (sa tri supstituenta i pozitivan naboj). Sumpor se ponaša kao meka Lewis kiselina, odnosno donator/elektrofil koji omogućuje S-metil grupi da se prenese na kisik, dušik ili aromatični sistem. To se često odvija uz pomoć drugih kofaktora, kao što je kobalamin (vitamina B12 kod ljudi). Neki enzimi koriste SAM za pokretanje radikalne reakcije, po čemu se nazivaju radikalni SAM enzimi.
Kao posljedica transfera metil grupe, dobija se S-adenozil-homocistein. U bakterija, ovo regenerira i metilacije ili se očuva uklanjanjem adenina i homocisteina, ostavljajući spoj dihidroksipentandion da se spontano pretvori u autoinducer-2, koji se izlučuje kao otpadni produkt.
Biosnteza i metabolizam
[uredi | uredi izvor]- EC 2.1.1 SAM-ovisne metil transferaze
- EC 2.1.1.5 Betain-homocistein S-metiltransferaza
- EC 2.1.1.13 Metionin sintaze
- EC 2.3.1.30 Serin acetiltransferaza
- EC 2.3.1.46 Homoserin O-sukciniltransferaza
- EC 2.5.1.6 metionin adenosiltransferaza
- EC 2.5.1.47 Cistein sintaza
- EC 2.5.1.48 Cistationin γ-sintaza
- EC 3.3.1.1 S-adenozil hidrolaza
- EC 4.1.1.57 metionin dekarboksilaza
- EC 4.2.1.22 cistationin β-sintaza
- EC 4.4.1.1 Cistationin γ -liaza
- EC 4.4.1.8 Cistationin-β-liaza

Enzimi uključeni u biosintezu metionina su:
- Aspartokinaza
- Β-aspartat semi- aldehid dehidrogenaza
- Homoserin dehidrogenaza
- Homoserin acetiltransferaza
- Cistationin-γ-sintaza
- Cistationin-β-liaza
- Metionin sintaza (u sisara, ovaj korak obavlja homocistein metiltransferaza)
Metionin se ne sintetizira u organizmu pa se mora unositi putem hrane. Ima ga u voću, mesu, uključujući i riblje, siru, povrću, posebno u spanaću, lješniku, grahu, pistaćima.[4]
Kao esencijalna aminokiselina, metionina se, kod ljudi i drugih životinja, ne sintetizira ‘’de novo’’ koji moraju konzumirati metionin ili proteine bogate metioninom. U biljkama i mikroorganizmima, biosinteze metionina se odvija u aspartatnoj porodici, zajedno sa treoninom i lizinom (ali ne preko α-aminoadipata). Glavna osnova je asparaginska kiselina, dok sumpora može doći od cisteina, metanetiola ili sumporovodika
- Asparaginska kiselina se najprije, preko β-aspartil-semi-aldehida, pretvara u homoserin, u dva smanjenja terminalne karboksilne grupe (homoserin ima stoga γ-hidroksil, otuda i prefiks homo). Srednji aspartat-semi-aldehid se grana skupa sa lizinovim biosintetskim putem, gdje je umjesto zgusnutog piruvata. Homoserin se povezuje sa treoninskim putem, gdje se, umjesto toga, isomerizira nakon aktiviranja termainalne hidroksilne grupe fosfata; u biljkama također učestvuje i u biosintezi metionina.
- Homoserin se tada aktivira fosfatom, sukcinil ili acetil hidroksilne grupe.
- U biljkama i možda u nekim bakterijama, koriste se fosfati. Ovaj korak dijeli s biosintezom treonina.
- U većini organizama, za aktiviranje homoserina, učestvuje acetil skupina. To se u bakterija može katalizirati enzimom koji je kodiran sa metX ili metA.
- U enterobakterija i ograničenom broju ostalih organizama, koristi se sukcinat. Enzim koji katalizira reakciju je meta prirode i specifičnost je za acetil-CoA i sukcinil-CoA. Fiziološka osnova za prednost acetil-CoA ili sukcinil-CoA je nepoznata, ali takve alternativne rute su prisutne u nekim drugim putevima (npr. u biosintezi lizina i arginina).
- Hidroksil aktivirajuća grupa se tada premješta sa cisteinom i metanethiolom ili sumporvodikom. Ova reakcija je tehnički γ-eliminacijska, prema Michaelovoj adiciji. Svi uključeni enzimi su homologni i članovi Cys/Met metabolizma PLP-zavisne porodice. Oni koriste kofaktor PLP (piridoksal fosfat), koji stabilizira ugljikove međuproizvode.
- Ako se reagira sa cisteinom, nastaje cistationin, koji je rascijepljen da popusti homocistein. Uključeni enzimi su cistationin-γ-sintaze (u bakterija kodirane iz metB) i cistationina-β-liaza (metC). Cistationin je drugačije vezan na dva enzima koji omogućavaju β-γ reakciju.
- Kada se reagira slobodnim sumporovodikom, nastaje homocistei, što katalizira O-acetilhomoserin aminokarboksipropiltransferaza (ranije poznata kao O-acetilhomoserin (tiol)-liaza. U bakterija je kodirana putem ili metY ili MetZ.
- Ako se reagira sa metanetiolom, metionin nastaje direktno. Metanetiol je nusprodukt kataboličkih puteva određenih sastojaka, pa je ova ruta jneuobičajena.
- Kada se homocistein proizvodi sa tiol grupom, metionin donosi metal. Poznate su dvije metionin sintaze: jedna je kobalamin (vitamin B12) zavisna, a druga nije.
Put korištenja cisteina se naziva "transsulfuracijski put", dok se put sumporovodika (ili metanetiola) naziva "direktni sulfurilacijskim put".[5]
Proizvodi slični cisteinu, naime, mogu biti napravljeni od jednog aktiviranog serina i jednog homocisteina (obrnuti trans-sulfurilacijski put) ili sumporovodika (direktni sulfurilacijski put); aktivirani serin je uglavnom O-acetil-serin (u Escherichia coli – preko CysK ili CysM), ali u Aeropyrum pernix i nekim drugi arheama koristi se O-fosfoserin. Pritom su dysK i cisM homologni, ali pripadaju PLP putu, grani tipa III.
Također pogledajte
[uredi | uredi izvor]Reference
[uredi | uredi izvor]- ^ Graeme K. Hunter G. K. (2000): Vital Forces. The discovery of the molecular basis of life. Academic Press, London 2000, ISBN 0-12-361811-8.
- ^ Nelson D. L., Michael M. Cox M. M. (2013): Lehninger Biochemie. Springer, ISBN 978-3-540-68637-8.
- ^ Nelson D. L., Michael M. Cox M. M. (2013): Lehninger Principles of Biochemistry. W. H. Freeman, 2013.ISBN 978-1-4641-0962-1.
- ^ Bajrović K, Jevrić-Čaušević A., Hadžiselimović R., Ed. (2005): Uvod u genetičko inženjerstvo i biotehnologiju. Institut za genetičko inženjerstvo i biotehnologiju (INGEB), Sarajevo, ISBN 9958-9344-1-8.
- ^ Kornberg A. (1989): For the love of enzymes – The Odyssay of a biochemist. Harvard University Press, Cambridge (Mass.), London,ISBN 0-674-30775-5, ISBN 0-674-30776-3.
Vanjski linkovi
[uredi | uredi izvor]- Computational Chemistry Wiki
- Food Sources of Methionine Arhivirano 30. 5. 2015. na Wayback Machine
- Foods containing methionine Arhivirano 15. 8. 2015. na Wayback Machine