과도기 화석
Transitional fossil| 다음에 대한 시리즈 일부 |
| 고생물학 |
|---|
 |
| 고생물학 포털 카테고리 |
과도기적 화석은 조상 집단과 그 파생된 후손 집단 모두에게 공통적인 특성을 보여주는 생명체의 화석화된 잔해다.[1]이것은 특히 후손 집단이 총체적 해부학 및 조상 집단과 생활 방식에 의해 극명하게 구별되는 경우에 중요하다.이 화석들은 분류학적 구분이 연속적인 변동에 대해 사후판단에 부과된 인간의 구성물이라는 것을 상기시켜주는 역할을 한다.화석 기록의 불완전성 때문에, 보통 과도기 화석이 분화점에 얼마나 가까운지 정확히 알 수 있는 방법은 없다.따라서 과도기 화석이 그러한 조상들의 모델로 자주 이용되고 있지만, 보다 최근의 집단의 직접적인 조상이라고 가정할 수는 없다.[2]
1859년 찰스 다윈의 '종의 기원'이 처음 출판되었을 때, 화석 기록은 잘 알려져 있지 않았다.다윈은 과도기적 화석의 부족을 "내 이론에 대해 촉구할 수 있는 가장 명백하고 중대한 반대"라고 설명했지만, 그는 그것을 지질학적 기록의 극도의 불완전성과 연관시켜 설명했다.[3]그는 당시 이용할 수 있는 제한된 수집품에 주목했지만, 이용 가능한 정보는 자연 선택을 통한 수정과 함께 그의 혈통 이론에 따른 패턴을 보여주는 것이라고 설명했다.[4]실제로, 아르카이옵테릭스는 불과 2년 후인 1861년에 발견되었으며, 이전의 비조류 공룡과 조류 사이의 전형적인 과도기적 형태를 나타낸다.그 이후로 더 많은 과도기 화석이 발견되었고, 현재 많은 과도기 화석을 포함하여 척추동물의 모든 부류가 어떻게 연관되어 있는지에 대한 증거가 풍부하다.[5]계급 수준의 전환의 구체적인 예로는 테트라포드와 물고기, 새와 공룡, 포유류, 그리고 "매머럴과 같은 파충류"가 있다.
인류 진화에 관한 대중적 저술에서 '연계 누락'이라는 용어는 호민관 진화 기록의 인식된 차이를 지칭하기 위해 광범위하게 사용되어 왔다.그것은 새로운 과도기적인 화석 발견을 언급하기 위해 가장 일반적으로 사용된다.그러나 과학자들은 진화 이전의 자연관을 지칭하기 때문에 이 용어를 사용하지 않는다.
진화론적 및 유전학적 분류학
계통 명명법의 전환

진화 분류학에서, 20세기 대부분 동안 지배적인 분류학의 형태로서 여전히 비전문학 교과서에 사용되고 있는 형태학적 유사성에 기초한 분류학에서는, 종종 서로 분기하여 진화 나무를 형성하는 "거품" 또는 "거품"으로 그려진다.[6]과도기적 형태는 해부학적으로 보면 여러 집단 사이에 떨어지는 것으로 보이며, 새로 갈라진 쇄골 안과 바깥에서 나오는 특성이 혼재되어 있다.[7]
1990년대에 클래드리스틱스가 확립되면서, 관계는 일반적으로 막대기와 같은 수치로 진화적 선형의 분기를 나타내는 클래도그램으로 표현되었다.서로 다른 소위 "자연적" 또는 "단조적" 집단은 중첩된 단위를 형성하며, 이것들만이 유전학적 이름을 부여받는다.전통적인 분류에서는 테트라포드(tetrapod)와 물고기가 서로 다른 두 집단으로 보이는 반면, 계통생성 테트라포드(tetrapod)는 물고기의 한 가지로 간주된다.따라서 클래지컬리티와 함께 더 이상 기성 집단 간의 전환이 없으며, 경과 화석이라는 용어는 잘못된 명칭이다.분화는 클라도그램에서 분기로 표현되는 그룹 내에서 발생한다.[8]
피복형적 맥락에서 과도기적 유기체는 한 가지의 초기 예를 나타내는 것으로 볼 수 있는데, 이 가지는 이전에 알려진 자손의 특징 중 일부가 아직 진화하지 않았다.[9]그러한 그룹의 초기 대표자들은 보통 화석 생물이 딸 클래드에 속하는지 여부에 따라 "[10]바살세아" 또는 "자매세아"라고 불린다.[8]
과도기 대 조상
혼란의 근원은 두 개의 서로 다른 분류학 그룹 사이의 과도기적 형태가 한 그룹 또는 두 그룹의 직접적인 조상이어야 한다는 개념이다.진화 분류법의 목표 중 하나가 다른 세자의 조상이었던 세자를 식별하는 것이라는 사실 때문에 어려움이 가중된다.그러나 진화는 사다리와 같은 진행을 만들어 내는 선형적인 과정보다는 관련 종의 복잡한 덤불 무늬를 만들어 내는 분쇄 과정이고, 화석 기록의 불완전성 때문에 화석 기록에 나타난 어떤 특정한 형태도 다른 어떤 것의 직접적인 조상일 가능성은 낮다.클래지스틱스는 한 분류학 집단이 다른 집단의 조상이라는 개념을 강조하고, 그 대신 다른 집단보다 더 최근의 공통 조상을 서로 공유하는 자매 택사의 식별을 강조한다.몇몇 해양 플랑크톤 마이크로포셀과 같이 몇 가지 예외적인 경우가 있는데, 화석 기록은 어떤 화석이 실제로 다른 종의 후대의 개체군에게 조상이 된 개체군을 나타낸다고 자신 있게 암시할 수 있을 정도로 충분히 완성되어 있다.[11]그러나 일반적으로 과도기적 화석은 실제 조상이라기보다는 서로 다른 세자의 실제 공통적인 조상들의 과도기적 해부학적 특징을 보여주는 특징을 가진 것으로 간주된다.[2]
두드러진 예
아르케오프테릭스

아크로옵테릭스는 조류와 밀접한 관련이 있는 공룡의 속이다.19세기 후반부터 팔래온학자들에게 받아들여져 왔으며, 2011년 한 연구는 이 평가에 의문을 제기해 왔으나, 가장 오래된 것으로 알려진 새로 알려져 있으며, 대신 새의 기원과 밀접한 관련이 있는 비아비안 공룡임을 시사했다.[12]
그것은 약 1억 5천만년 전 후기 쥬라기 시대에 지금의 독일 남부에 살았는데, 당시 유럽은 지금보다 훨씬 더 적도에 가까운 얕은 따뜻한 열대 바다에 있는 군도였다.유럽의 까치와 비슷한 모양으로, 가장 큰 개체가 까마귀의 크기를 가질 수 있는,[13] 아크로옵테릭스는 길이가 약 0.5미터(1.6피트)까지 자랄 수 있다.아크로옵테릭스는 작은 크기와 넓은 날개, 유추된 비행 능력에도 불구하고 현대 조류보다 다른 작은 중생대 공룡들과 공통점이 더 많다.특히, 그것은 날카로운 이빨을 가진 턱, 발톱을 가진 손가락 세 개, 긴 뼈 꼬리, 과대증식 가능한 두 번째 발가락("킬링 발톱"), 깃털(홈오테미를 암시하는 것), 그리고 다양한 골격의 특징과 같은 특징을 공유한다.[14]이러한 특징들은 아크로옵테릭스를 공룡과 새들 사이의 과도기적 화석의 확실한 후보자로 만들어 공룡과 새의 기원을 연구하는 데 모두 중요한 역할을 하게 한다.[15]
최초의 완전한 표본은 1861년에 발표되었고, 그 이후로 10개의 고고학 화석이 더 발견되었다.알려진 11개의 화석은 대부분 깃털의 인상을 포함하고 있는데, 이는 그러한 구조물에 대한 가장 오래된 직접적인 증거 중 하나이다.게다가, 이 깃털들은 비행깃털의 진보된 형태를 취하고 있기 때문에, 아크로옵테릭스 화석은 깃털이 후기 쥬라기 이전에 진화하기 시작했다는 증거다.[16]
오스트랄로피테쿠스 아파렌시스

오스트랄로피테쿠스 아파렌시스라는 호민관은 현대의 두발 달린 인간과 그들의 네발 달린 유인원 조상 사이의 진화적 전환을 나타낸다.일부 연구자들이 A. A. A. A. A. A. A. A. A. A. A. A. A. A. A. A. A. A. A. A.의 많은 특징들은 두발성을 강하게 반영하고 있다.[17]전체 해부학에서 골반은 유인원보다 훨씬 인간적이다.장골날은 짧고 넓으며 천골은 넓고 엉덩이 관절 바로 뒤에 위치하며, 무릎 확장기에 강한 애착이 있다는 명백한 증거가 있어 곧은 자세를 암시한다.[17]: 122
골반이 완전히 사람의 그것과 같지는 않지만, 이러한 특징들은 상당히 많은 양의 두발성을 수용하도록 획기적으로 리모델링된 구조를 가리킨다.대퇴골은 엉덩이에서 무릎 쪽으로 비스듬히 들어간다.이 특성은 발이 신체의 중간선 가까이 떨어질 수 있게 하며, 습관적인 두발 운동을 강하게 나타낸다.오늘날 인간, 오랑우탄, 거미원숭이는 이와 같은 특징을 가지고 있다.발에는 큰 발가락을 기댄 것이 특징으로, 뒷다리로 나뭇가지를 잡는 것이 불가능하지 않다면 곤란하다.A. 아파렌시스는 운동 외에도 현대의 침팬지[18](인간과 가장 가까운 친척)보다 약간 큰 뇌를 가지고 있었고 유인원보다 더 인간적인 치아를 가지고 있었다.[19]
파키세티데스, 앰뷸런스코투스


고래, 돌고래, 고슴도치는 육지 포유류의 해양 포유류 후손이다.파키세티드는 가장 초기 고래인 발굽이 달린 포유류의 멸종된 종으로, 이들의 가장 가까운 자매 집단은 라오엘레과의 인도히우스다.[20][21]그들은 약 5천 3백만년 전에 Early Eocene에서 살았다.그들의 화석은 1979년 파키스탄 북에서 처음 발견되었는데, 옛 테티스 해의 해안에서 멀지 않은 강이었다.[22][page needed]파키세티드는 대부분의 육지 포유동물처럼 고엽막에 의존하기 보다는 강화된 뼈 전도를 사용하여 물속에서 들을 수 있었다.이 배열은 물밑에서 방향 청력을 제공하지 않는다.[23]
약 4900만년 전에 살았던 구급차 나탄은 1994년 파키스탄에서 발견되었다.그것은 아마도 수륙양용이었고, 악어처럼 보였다.[24]Eocene에서는 구급차들이 파키스탄 북부의 Tethys 해양의 만과 하원에 서식했다.[25]구급차의 화석은 풍부한 해양 식물 화석과 연골 연체동물과 관련된 근해의 얕은 해양 퇴적물에서 항상 발견된다.[25]비록 해양 퇴적물에서만 발견되지만, 그들의 산소 동위원소 값은 염도 범위의 물을 소비했음을 나타내며, 어떤 표본은 해수 소비의 증거가 없고, 다른 표본들은 치아가 화석화된 당시 신선한 물의 소비량이 전혀 없었다.구급차들이 광범위한 염분 농도를 용인했다는 것은 분명하다.[26]그들의 식단은 아마도 물을 마시기 위해 접근한 육지 동물이나 강에 사는 민물 수생 생물들을 포함했을 것이다.[25]따라서, 구급차는 민물과 해양 서식지 사이의 고래 조상들의 이동 단계를 나타낸다.
틱타알릭


틱타알릭은 후기 데본기 시대의 멸종된 사랍어(지느러미)[27]의 속성으로, 테트라포드(네발 동물)와 비슷한 특징이 많다.그것은 그 시대의 산소가 부족한 얕은 물 서식지에 적응하는 것을 발전시키기 위한 고대 사복류 동물들 중 하나인데, 이것은 사복류의 진화를 이끈 적응이다.[28]잘 보존된 화석은 2004년 캐나다 누나부트의 엘레스미어 섬에서 발견되었다.[29]
틱타알릭은 약 3억 7천 5백만년 전에 살았다.고생물학자들은 이것이 3억 8천만년 된 화석으로부터 알려진 판데르이치스와 같은 비테트라포드 척추동물과 약 3억 6천 5백만년 된 화석으로부터 알려진 아칸토스테가와 이크티오스테가와 같은 초기 테트라포드 사이의 전환을 대표한다고 제안한다.원시 물고기와 파생된 4각류 특성이 뒤섞여 발견자 중 한 명인 닐 슈빈(Neil Shubin)이 틱타알릭을 "피시파드"로 특징 짓게 되었다.[30][31]이전의 많은 물고기 같은 과도기 화석과 달리 틱타알릭의 '핀'은 기본적인 손목뼈와 손가락을 연상시키는 단순한 광선을 가지고 있다.그들은 체중을 견디고 있었는지도 모른다.현대의 모든 사다리꼴과 마찬가지로 갈비뼈, 가슴띠가 분리된 이동식 목, 물고기의 아가미, 비늘, 지느러미를 가지고 있었지만 폐를 가지고 있었다.[27]그러나 2008년 Boisvert의 논문 al.Panderichthys는 더 파생된 원위부 때문에 Tiktaalik보다 테트라포드에 더 가까울 수 있으며, 이는 수렴 진화에 의해 테트라포드와 독립적으로 유사성을 발전시켰을 수 있다.[32]
네발 동물 발자국이 그를 폴란드에서 자연에서 2010년 1월 보고 발견된 10백만 년들 가장 오래된 것으로 알려진 elpistostegids[33](그 중에게 '틱타알릭'이라는 한 예입니다)보다 나이가 많에서 직접 tr이 아니라에게 '틱타알릭'이라는 같은 동물들에 400만년 전에 진화되는 기능이 왔다late-surviving 유물을 내포한다"안전하게 데이트" 있었다.ansi그리고 그것들은 우리가 육지 척추동물의 초기 역사에 대해 얼마나 잘 알지 못하는지를 강조한다."[34]
Amphistium


회오리목(flattornectiform)은 광어류(bright-fined fish의 주문이다.현대 광어의 가장 분명한 특징은 두 눈이 어른 물고기의 머리와 같은 쪽에 있는 비대칭이다.어떤 집안에서는 눈이 항상 몸의 오른쪽에 있고(등쪽 또는 오른쪽 눈 광어) 다른 가족에서는 항상 왼쪽에 있다(불신 또는 왼쪽 눈 광어).원시 가시가 있는 터빈은 우와 좌의 눈이 같은 개인을 포함하며, 일반적으로 다른 가족에 비해 비대칭성이 낮다.그 외에 돌기가 있는 눈의 존재, 해저에 사는 것에 대한 또 다른 적응(벤트호스), 등지느러미를 머리로 확장하는 것이 그 특징이다.[35]
암피스티움은 광어의 초기 친척으로 확인된 5천만년 된 화석 어류로, 암피스티움에서는 척추동물의 전형적인 대칭 머리로부터의 전환이[36] 불완전하며, 한쪽 눈은 머리 중앙 부근에 위치한다.[37]고생물학자들은 "연구자들이 한때 믿을 수밖에 없었던 것처럼 갑자기 일어난 것이 아니라 자연 선택을 통한 진화와 일치하는 방식으로 점차적으로 변화가 일어났다"[36]고 결론지었다.
암피스티움은 루테티아 이탈리아의 몬테 볼카 라거스테테로부터 알려진 많은 화석 물고기 종들 중 하나이다.헤테로네테스는 프랑스의 조금 전의 층에서 나온 매우 유사한 화석이다.[37]
룬캐리아

벨기에에서 종자식물의 미들 데보니아 전구체가 확인되어 초기 종자식물을 약 2천만년 앞당겼다.Runcaria는 작고 방사상 대칭으로, 큐플로 둘러싸인 정합 메가스포르랑기움이다.메가스포르랑기움은 다층계 위에 돌출된 미개봉 원위 확장체를 가지고 있다.그 연장은 혐기성 수분작용에 관여했다는 의심을 받고 있다.런캐리아는 단단한 종자 외투와 꽃가루를 씨앗으로 인도하는 시스템을 제외한 씨앗 식물의 모든 자질을 갖추면서 씨앗으로 이어지는 인물 획득의 순서를 새롭게 조명한다.[38]
화석 기록
화석 기록이 완전하지 않기 때문에 모든 과도기적 형태가 화석 기록에 나타나는 것은 아니다.유기체는 최상의 환경 속에서 화석으로 거의 보존되지 않을 뿐이며, 그러한 화석의 극히 일부만이 발견되었다.고생물학자 도널드 프로테로는 화석 기록을 통해 알려진 종의 수가 알려진 생물 종 수의 5% 미만이라는 사실에서 이것이 설명된다고 언급하면서 화석을 통해 알려진 종의 수가 지금까지 살아왔던 모든 종의 1% 미만임에 틀림없음을 시사했다.[39]
생물학적 구조가 화석을 만드는 데 필요한 전문적이고 드문 환경 때문에, 논리는 알려진 화석이 지금까지 존재했던 모든 생명체의 극히 일부분만을 나타내며, 각각의 발견은 진화의 스냅숏만을 나타낸다는 것을 지시한다.과도기적 화석은 전이적 화석을 통해서만 설명되고 확증될 수 있는데, 이것은 결코 분명히 다른 형태들 사이의 정확한 중간 지점을 증명하지 못한다.[40]
화석 기록은 매우 고르지 않고 거의 예외 없이 단단한 부분이 있는 유기체 쪽으로 심하게 기울어져 있어 대부분의 부드러운 체질의 유기체 그룹은 화석 기록이 거의 또는 전혀 없다.[39]전통적인 집단들 사이의 많은 과도기적 화석을 포함하여, 좋은 화석 기록을 가지고 있다고 여겨지는 집단은 척추동물, 에치노데름, 브라치오포드와 몇몇 절지동물 집단들이다.[41]
역사
포스트 다윈


동물과 식물의 종은 일정하지 않고 시간이 흐르면서 변한다는 생각은 18세기까지 제안되었다.[42]1859년에 출판된 다윈의 "종의 기원"은 다윈에게 확고한 과학적 근거를 주었다.그러나 다윈의 작업의 약점은 다윈 자신이 지적한 팔래온학적 증거의 부족이었다.자연 선택이 일반과 가족 내에서 보이는 변화를 만들어 내는 것을 상상하는 것은 쉽지만, 상위 범주 간의 변화는 상상하기 더 어려웠다.다윈의 작품이 출판된 지 불과 2년 후인 1861년 런던 시료의 극적인 발견은 고도로 파생된 새들의 종류와 보다 기초적인 파충류들의 종류 사이의 관계를 처음으로 제공했다.[43]다윈에게 보낸 편지에서, 팔레온테리학자 휴 팔콘거는 다음과 같이 썼다.
솔른호펜 채석장들이 8월 명령으로 다윈이라는 낯선 존재로 밝혀지기 위해 임관되었다면, 그것은 아르케오프테릭스보다 더 훌륭하게 가장 많은 것을 처형할 수 없었을 것이다.[44]
따라서, 아크로옵테릭스와 같은 과도기적 화석은 다윈의 이론을 확증할 뿐만 아니라, 그 자체로 진화의 아이콘으로 여겨지게 되었다.[45]예를 들어, 1904년의 스웨덴 백과사전 노디스크 패밀제복은 "ettaf de betydelsefullaste polontologiska fynd, som nggonsin gjorts" ("지금까지 행해진 가장 중요한 고생물학적 발견 중 하나") 화석의 부정확한 Arcogopteryx 재구성을 보여주었다.[46]
식물의 상승
과도기적 화석은 동물의 화석만이 아니다.20세기 초에 식물의 분열을 지도화하면서, 혈관 식물의 조상에 대한 탐색이 시작되었다.1917년 로버트 키드스톤과 윌리엄 헨리 랭은 스코틀랜드 애버딘셔의 라이니 체르트에서 극히 원시적인 식물의 잔해를 발견하여 라이니아라고 이름지었다.[47]
라이니아 식물은 작고 막대기처럼 생겼는데, 잎이 없는 줄기는 단순하게 이분법적으로 가지를 쳐 각각 포자낭에 의해 기울어져 있었다.단순한 형태는 이끼의 산발적인 형태와 반향되며, 리니아는 세대를 번갈아 가며, 작은 줄기의 혼잡한 형태의 해당 생식세포와 높이가 불과 몇 밀리미터에 불과한 것으로 나타났다.[48]따라서 라이니아는 양치류나 클럽모스와 같은 초기 혈관 식물과 이끼 사이에 중간쯤에 떨어진다.이끼와 같은 생식기의 카펫에서, 더 큰 라이니아 산발물은 단순한 몽둥이와 비슷하게 성장했고, 식물을 기질에 고정시킨, 수평으로 자라는 줄기의 성장한 줄기에 의해 퍼졌다.이끼와 같은 특징과 혈관의 특이한 혼합과 식물의 극단적인 구조적 단순성은 식물학적 이해에 커다란 영향을 미쳤다.[49]
누락된 링크

모든 생명체가 어떤 종류의 전이 과정을 통해 연관되어 있다는 생각은 다윈의 진화론을 앞서는 것이다.장바티스트 라마크는 생명체가 가장 단순한 생물체의 형태로 끊임없이 생성되는 것을 상상했고, 점진적인 일련의 낮은 형태를 통해 복잡성과 완벽함(즉 인간)을 향해 노력했다.[50]그의 견해로는 하등 동물들은 진화 현장의 신참에 불과했다.[51]
'종의 기원' 이후, 에른스트 해켈의 인간 혈통에서 보여지듯이 진화 초기 단계를 대표하는 "하위 동물"에 대한 생각은 여전히 남아 있었다.[52]그 후 척추동물이 일종의 진화적 서열을 형성하는 것으로 여겨졌지만, 다양한 계층은 구별되는 반면, 발견되지 않은 중간 형태는 "연계 누락"이라고 불린다.
이 용어는 찰스 리엘이 지질학 칼럼의 누락된 부분과 관련하여 저서 『지질학의 요소들』 제3판(1851)에서 과학적인 맥락에서 처음 사용하였으나, 1863년 저서 『인간 고대의 지질학적 증거들』의 xi 페이지에 나타나면서 현재의 의미로 대중화되었다.그 무렵에, 일반적으로 마지막 빙하기의 끝이 인류 최초의 모습을 나타낸다고 생각되었다; 리엘은 인간의 기원을 훨씬 더 뒤로 돌려놓기 위해 인간 고대의 새로운 발견들을 그렸다.Lyell은 인간과 짐승의 거대한 괴리가 어떻게 연결될 수 있는지 깊은 미스터리로 남아있다고 썼다.[53]리엘의 생생한 글씨는 대중의 상상력을 불러일으키며 쥘 베른의 지구 중심 여행(1864년)과 루이 피구에의 1867년 초판("홍수 앞의 지구")을 감동시켰는데, 이 책은 짐승 가죽을 입고 돌도끼를 휘두르는 야만인 남녀의 극적인 삽화를 담고 있었다.1863년 판에 나타난 소굴.[54]
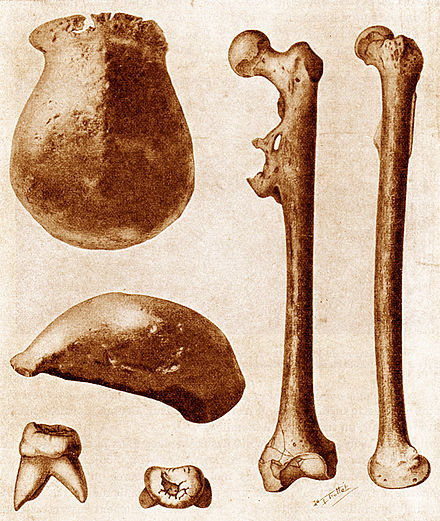
그러나 1891년 네덜란드의 젊은 지질학자 외젠 두부아가 자바 솔로 강둑에서 두개골 캡, 어금니, 대퇴골을 발견하기 전까지는 유인원과 인간 사이의 과도기적 특성을 보여주는 화석을 찾는 것은 아무런 성과가 없었다.이 발견은 낮은 유인원 모양의 두개골 지붕과 침팬지와 성인 인간의 두개골 중간인 약 1000cc로 추정되는 뇌를 결합시켰다.단 하나의 어금니는 현대의 어떤 사람의 치아보다 컸지만 대퇴골은 길고 곧았으며 무릎 각도로 '자바 맨'이 똑바로 걸었다는 것을 알 수 있었다.[55]피테칸트로푸스 에렉투스("정확한 유인원")라는 이름으로 보아, 그것은 현재 인류 진화 화석의 긴 목록 중 첫 번째가 되었다.그 당시 그것은 많은 사람들에 의해 "실종 고리"라는 찬사를 받았으며, 비록 그것이 공룡과 새의 중간 매개체인 아르케오옵테릭스 같은 다른 매개체들에게 사용되기도 하지만, 인간의 화석을 위해 주로 사용되는 용어로서 이 용어를 설정하는 것을 도왔다.[56][57]

'연계 누락'은 여전히 대중에게 잘 알려져 있고 대중 매체에서 자주 사용되는 인기 용어지만, 과학 출판물에서는 이 용어를 피한다.[58][5]일부 블로거들은 그 연결고리가 더 이상 "실종"되지 않기 때문에 그리고 인간의 진화가 더 이상 단일 선형 진행 측면에서 발생했다고 믿지 않기 때문에 그것을 "부적절하다"[59]라고 불렀다.[5][60]
구두점 평형
스테판 제이 굴드와 나일스 엘드레지에 의해 개발되어[61] 1972년에 처음 제시된 구두점 평형 이론은 종종 과도기 화석의 논의에 잘못 끌려들어간다.[62]그러나 이 이론은 지질학적으로 짧은 기간에 걸쳐 세자 내 또는 밀접하게 연관된 세자 사이의 잘 문서화된 전환에만 관련된다.일반적으로 동일한 지질학적 아웃크로 추적 가능한 이러한 전환은 형태학적 안정성의 연장기간의 형태학에서 작은 점프를 보여준다.이러한 점프를 설명하기 위해 Gould와 Eldredge는 비교적 긴 기간 동안 급속한 진화의 기간으로 분리된 유전적 안정성을 예상하였다.굴드는 과도기적 화석의 존재를 부정하기 위해 창조론자의 작품 오용에 대해 다음과 같은 관찰을 했다.
우리가 추세를 설명하기 위해 구두점 평형을 제안했기 때문에, 화석 기록에 과도기적인 형태가 없다는 것을 인정하는 것과 같이, 디자인을 통해서든 어리석음을 통해서든 창조론자들에 의해 계속해서 인용되는 것은 분개하고 있다.종단부는 종 수준에서 발생하며, 주요 집단 내에서 더 높은 수준의 전환에서 (계단 모델의) 방향 추세가 만연한다.
참고 항목
참조
- ^ 프리먼 & 헤론 2004 페이지 816
- ^ a b 프로테로 2007, 페이지 133-135
- ^ 다윈 1859, 페이지 279–280
- ^ 다윈 1859, 페이지 341–343
- ^ a b c Prothero, Donald R. (1 March 2008). "Evolution: What missing link?". New Scientist. 197 (2645): 35–41. doi:10.1016/s0262-4079(08)60548-5. ISSN 0262-4079.
- ^ 예를 들어, 벤튼 1997을 참조하십시오.
- ^ 프로테로 2007, 페이지 84.
- ^ a b Kazlev, M. Alan. "Amphibians, Systematics, and Cladistics". Palaeos. Retrieved 9 May 2012.
- ^ 프로테로 2007, 페이지 127.
- ^ 프로테로 2007, 263페이지.
- ^ Prothero, Donald R.; Lazarus, David B. (June 1980). "Planktonic Microfossils and the Recognition of Ancestors". Systematic Biology. 29 (2): 119–129. doi:10.1093/sysbio/29.2.119. ISSN 1063-5157.
- ^ Xing Xu; Hailu You; Kai Du; Fenglu Han (28 July 2011). "An Archaeopteryx-like theropod from China and the origin of Avialae". Nature. 475 (7357): 465–470. doi:10.1038/nature10288. ISSN 0028-0836. PMID 21796204. S2CID 205225790.
- ^ Erickson, Gregory M.; Rauhut, Oliver W. M.; Zhonghe Zhou; et al. (9 October 2009). "Was Dinosaurian Physiology Inherited by Birds? Reconciling Slow Growth in Archaeopteryx". PLOS One. 4 (10): e7390. Bibcode:2009PLoSO...4.7390E. doi:10.1371/journal.pone.0007390. ISSN 1545-7885. PMC 2756958. PMID 19816582.