트리쵸메
Trichome

트리콤(/tratrakokommz, trtrkokommz/, 고대 그리스어 μα(트리코마)의 '털')은 식물, 조류, 지의류 및 특정 원생물의 미세한 성장 또는 부속물이다.그것들은 다양한 구조와 기능을 가지고 있다.예로는 털, 선모, 비늘, 유두 등이 있습니다.식물상의 어떤 종류의 털도 덮는 것은 유도체이며, 그것을 가진 표면은 사춘기라고 한다.
알갈 트리콤
특정한, 보통 필라멘트 모양의, 조류는 말단 세포가 [example needed]트리콤이라고 불리는 가늘고 긴 머리카락 같은 구조로 생산된다.스피루리나나 오발레이터리아와 같은 일부 시아노박테리아에 있는 이러한 구조에도 동일한 용어가 적용됩니다.시아노박테리아의 트리콤은 발진기에서처럼 덮이지 않거나 [1]칼로트릭스에서처럼 덮을 수 있다.이러한 구조물은 특히 추운 사막 [citation needed]기후에서 토양 침식을 방지하는 데 중요한 역할을 한다.필라멘트 모양의 칼집은 토양 구조를 유지하는 데 도움이 되는 끈적끈적한 네트워크를 형성합니다.
식물 트리콤





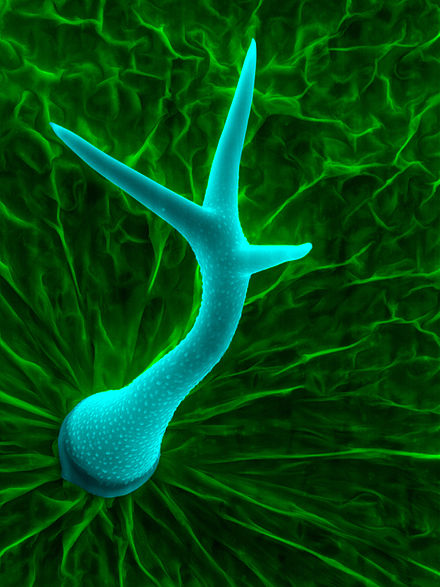


식물 트리콤은 식물의 종과 개별 식물의 장기 사이에 서로 다른 많은 다른 특징을 가지고 있다.이러한 기능은 트리콤이 배치되는 하위 카테고리에 영향을 미칩니다.정의 기능에는 다음과 같은 것이 있습니다.
- 단세포 또는 다세포
- 직선(분기가 거의 없거나 전혀 없는 오른쪽), 나선형(코르크 나사 모양) 또는 후크형(곡선 꼭대기)[2]
- 세포질 유무
- 선(분비) vs.엘란도라
- 구부러짐, 단순(가지 없는 단세포), 펠테이트(비늘 모양), 성형(별 모양)[3]
- 단축 대 단축 - 잎 또는 다른 측방 기관의 윗면(단축) 또는 아랫면(단축)에 각각 트리콤이 존재하는지 여부를 나타냅니다.
모델 생물인 C. salvifolius에서, 이 표면은 유축 [4]표면보다 더 많은 자외선, 태양 복사 강도 빛 스트레스에 시달리기 때문에 이 식물에는 더 많은 단축 트리콤이 존재합니다.
트리콤은 자외선, 곤충, 증산,[5] 동결불내증 등과 같은 다양한 유해성으로부터 식물을 보호할 수 있습니다.
공중 표면 털
다양한 종류의 식물 Trichomes 있표피는 것이다.그 조건 또는 가시로는 표피 이상을 포함하는 것이다를 참조하십시오 emergences.이 구분은 쉽게 적용되(Wait-a-minute tree[동음 이의 필요한] 보).또한, 표면에서 나왔다.nontrichomatous 표피 세포가 있다.[예가 필요했다]
trichome의 보편적인 유형의 머리.식물의 털이나 다세포, 분지나 가지로 갈라지지 않은 단세폴 수 있다.다세포 털 세포 하나 또는 여러겹을 가질 수 있다.Branched 털(나뭇가지형)캥거루 발(Anigozanthos), 박새, 또는 성상(별 모양의)에서처럼, 애기 장대 thaliana에 수지상 수 있다.
세포 일부를 직접적으로 종류의 줄기에 나는 표면 또는 감염에 부착된 한접시와 방패 모양의 집합체를 가지고 있trichome의 또 다른 보편적인 타입은 규모나 비늘 털,.그 파인애플, 진달래 속과 바다buckthorn(Hippophae rhamnoides) 같은 bromeliads의 흔한 예는 잎 규모이다.
다양한 종류의 털은 선상이고, 민트나 라미과에서 생산되는 에센셜 오일과 같은 종류의 분비물을 만들어 낼 수 있다.
줄기나 잎과 같은 식물 기관의 표면적인 모습을 묘사하기 위해 많은 용어들이 트리콤의 존재, 형태, 외관을 언급하기 위해 사용된다.예를 들어 다음과 같습니다.
- 광택이 있는, 광택이 있는, 광택이 없는, 털이나 트리콤이 없는, 표면이 매끄러운
- 히루수트 – 털이 거칠다
- 히스피드 – 뻣뻣한 털을 가진
- 관절형 – 단순 다세포 단모
- 솜털 – 긴 털로 된 거의 양털 같은 덮개를 가진
- 필로스 – 긴 털, 곧은 털, 부드러운 털, 넓게 뻗은 털 또는 곧은 털을 가진 사춘기
- 화농성 – 미세한 사춘기; 가늘고 짧으며 보통 곧은 털을 가지고 있다.
- 자생성 – 부드럽고 곧은 미세한 털로 약간 덮여 있다
- 사춘기 – 모든 종류의 털 또는 트리콤을 가진
- strigillose – 미세한 strigose
- 스트리고스 – 가장자리 또는 중간턱을 따라 거의 같은 방향을 가리키는 생머리를 가진
- 장신구 – 미세한 장신구
- 털갈이 – 촘촘하고 매트가 있는 털로 덮여 있다
- 별장 – 아주 작은 별장
- 융모 – 길고 부드러운 털을 가지며 종종 곡선을 그리지만 매트가 없는
식물에 있는 털의 크기, 형태, 밀도, 위치는 종에 따라 그리고 심지어 다른 식물 장기에 있는 종에 따라 매우 다양합니다.표면 털이 있는 것의 몇 가지 기본적인 기능 또는 장점을 열거할 수 있습니다.많은 경우 털은 적어도 일부 작은 초식동물들의 먹이에 방해가 될 수 있으며, 입천장에 대한 경직과 과민성에 따라 큰 초식동물들도 마찬가지일 수 있다.서리가 내리기 쉬운 지역에서 자라는 식물의 털은 살아있는 표면 세포로부터 서리를 막아줍니다.바람이 많이 부는 곳에서는 털이 식물 표면을 가로지르는 공기의 흐름을 방해하여 증산을 줄입니다.털의 촘촘한 코팅은 햇빛을 반사하여 뜨겁고 건조하며 개방된 서식지에서 아래의 더 섬세한 조직을 보호합니다.또한, 사용 가능한 수분의 많은 부분이 안개 방울로부터 오는 장소에서는, 머리카락이 [citation needed]물방울이 쌓일 수 있는 표면적을 증가시킴으로써 이 과정을 강화시키는 것으로 보입니다.
선삼염색체
선모양 트리콤은 식물의 약 30%에서만 발견되지만 광범위하게 연구되어 왔다.그들의 기능은 식물의 대사물을 분비하는 것이다.이러한 대사물에는 다음이 포함됩니다.
- 성장과 발달에[6] 관련된 식물에서 많은 기능을 가진 테르페노이드
- 페닐프로파노이드: 2차 대사물, 스트레스 반응과 같은 많은 식물 경로에서 역할을 하며 환경에서의[7] 식물 상호작용의 매개자 역할을 한다.
- 플라보노이드
- 메틸케톤
- 아크릴수거[8]
비구형 트리콤
비골상 트리콤은 식물을 자외선으로부터 [4]보호하는 데 중요하다.
모델 식물인 Cistus salvifolius는 지중해 연안의 고휘도 스트레스와 열악한 토양 조건 지역에서 발견된다.그것은 방사선과 식물 건조의 흡수에 영향을 미치는 폴리페놀을 합성하고 저장하는 능력을 가진 비-glandular, Steatellite 및 Dendrite 트리콤을 포함한다.이들 트리콤은 또한 UV-B를 흡수할 수 있는 아세틸화 플라보노이드와 더 긴 파장의 UV-A를 흡수하는 비아세틸화 플라보노이드를 포함합니다.비글랜드 트리콤에서 플라보노이드의 유일한 역할은 식물을 보호하기 위해 가장 짧은 파장을 차단하는 것입니다; 이것은 선상 [4]트리콤에서의 그들의 역할과는 다릅니다.
폴리페놀류
Cistus속의 비-glandular trichomes는 엘라기타닌, 글리코사이드 및 켐페롤 유도체를 포함하는 것으로 밝혀졌다.엘라기타닌은 영양소를 제한하는 스트레스를 [4]받을 때 적응을 돕는 주요 목적을 가지고 있습니다.
트리콤과 뿌리모발육
많은 혈관 식물의 뿌리인 트리콤과 뿌리 털은 모두 표피층의 단일 세포에서 바깥쪽으로 자란다.뿌리털은 식물의 뿌리 표피에 있는 털을 형성하는 세포인 트리코플라스트에서 형성된다.뿌리털은 지름이 5~17마이크로미터, 길이가 80~1500마이크로미터로 다양하다(디트마, 에서, 1965년 인용).뿌리털은 2주에서 3주 정도 생존한 후 사라질 수 있습니다.동시에 뿌리의 꼭대기에는 새로운 뿌리털이 지속적으로 형성되고 있다.이렇게 하면 뿌리털 커버가 그대로 유지됩니다.따라서 뿌리털이 대부분 뽑히기 때문에 재배포는 주의해서 해야 한다는 것은 이해할 수 있다.이것이 식물을 심는 것이 식물을 시들게 할 수 있는 이유이다.
트리콤과 뿌리털의 패턴 형성에 대한 유전적 제어는 유사한 제어 메커니즘을 공유합니다.두 과정 모두 표피 발육의 시작과 발달을 제어하는 관련 전사 인자의 핵심을 포함한다.특정 단백질 전사인자(GLABRA1(GL1), GLABRA3(GL3) 및 Transparent TESTA GLABRA1(TTG1)을 코드하는 유전자의 활성화는 트리콤 또는 [9]뿌리털을 생성하는 세포운명의 주요 조절제이다.이 유전자들이 잎 표피 세포에서 활성화되면, 그 세포 내에서 트리크롬의 형성이 시작됩니다.GL1, GL3 및 TTG1은 또한 음의 조절제를 활성화하여 인접 세포에서 트리크롬 형성을 억제하는 역할을 한다.이 시스템은 잎 표면의 트리콤 간격을 조절합니다.일단 트리콤이 발달하면 그들은 분열하거나 [10]갈라질 수 있다.반면 뿌리털은 거의 가지를 치지 않는다.트리콤과 뿌리털이 형성되는 동안, 많은 효소들이 조절된다.예를 들어, 뿌리털이 생기기 직전에, 포스포릴라아제 [11]활성이 높아지는 지점이 있다.
과학자들이 트리콤 발달에 대해 알고 있는 많은 것들은 모델 유기체 Arabidopsis Thaliana에서 유래한다. 왜냐하면 그들의 트리콤은 단순하고 단세포이며, 비글랜드 형태이기 때문이다.발생 경로는 R2R3 MYB, 기본 나선-루프-나선 및 WD40 반복의 세 가지 전사 인자에 의해 조절됩니다.세 그룹의 TF는 트리머 복합체(MBW)를 형성하고 하류 제품의 발현을 활성화하여 트리콤 형성을 활성화한다.단, MYBs만 [12]네거티브 복합체를 형성하여 억제제 역할을 한다.
피토호르몬류
식물 피토호르몬은 식물의 성장과 환경 자극에 대한 반응에 영향을 미친다.이러한 피토호르몬 중 일부는 지베렐산, 사이토키닌, 그리고 자스몬산(JA)[12]을 포함하는 트리콤 형성에 관여합니다.
GA는 GLABRUS1(GL1)을 자극하여 트리콤의 성장을 촉진한다.
그러나, SPINDLY와 DELLA 단백질은 모두 GA의 영향을 억제하기 때문에, 이러한 단백질 중 더 적은 양이 더 많은 트리콤을 생성한다.
트리콤의 성장을 촉진하는 다른 피토호르몬은 브라시노스테로이드, 에틸렌, 살리실산을 포함한다.이것은 각각의 물질이 거의 또는 전혀 없는 돌연변이를 대상으로 실험을 함으로써 이해되었다.모든 경우에,[12] 존재하는 트리콤의 부정확한 형성뿐만 아니라 두 식물 표면 모두에서 트리콤 형성이 적었습니다.
분류법에 대한 중요성
트리콤의 종류, 존재 여부 및 위치는 식물 식별 및 식물 [13]분류에서 중요한 진단 특성이다.법의학적 검사에서, Cannabis sativa와 같은 식물은 트리콤의 [14][15]현미경 검사로 식별할 수 있다.화석에서 보존된 트리콤은 거의 발견되지 않지만, 트리콤 베이스는 정기적으로 발견되며, 어떤 경우에는 그들의 세포 구조가 식별에 중요하다.
아라비도시스탈리아나트리콤 분류
아라비도시스 탈리아나 트리콤은 공중,[16] 표피, 단세포, 관상구조로 분류된다.
식물 분자생물학의 의의
GLABRUS1 단백질에 의해 트리콤 형성이 개시되는 모델 식물 아라비도시스 탈리아나.대응하는 유전자의 녹아웃은 무광식물로 이어진다.이 표현형은 이미 게놈 편집 실험에서 사용되었으며 CRISPR/Cas9과 [17][18]같은 유전자 편집 방법을 개선하기 위한 식물 연구의 시각적 지표로서 관심을 가질 수 있다.트리콤은 [19]또한 식물의 패턴 형성과 세포 분화를 위한 모델 역할을 한다.
사용하다
콩잎은 역사적으로 동유럽의 집에 빈대를 가두는 데 사용되어 왔다.콩잎에 있는 트리콤은 발을 찔러서 곤충을 잡습니다.그러면 나뭇잎들은 [20]파괴될 것이다.
트리콤은 유럽 양털벌(Antidium manicatum)을 위한 둥지 건축의 필수적인 부분입니다.이 벌 종은 식물에서 트리콤을 떼어내 [21]둥지 공동을 위한 안감으로 사용함으로써 그들의 둥지에 트리콤을 통합합니다.
방어
식물은 물리적 및/또는 화학적 수단을 통해 초식동물의 공격을 억제하기 위해 트리콤을 사용할 수 있다. 예를 들어 히스타민과 같은 염증성 화학물질을 전달하는 우티카(네틀) 종의 특수하고 따끔따끔한 털이다.트리콤에 대한 연구는 초식동물을 억제한 결과인 농작물 보호에 초점을 맞추고 있다(Brookes et al. 2016).[22]하지만, 몇몇 생물들은 트리콤의 효과에 저항하는 메커니즘을 발전시켰다.예를 들어 헬리코니어스의 애벌레는 트리콤으로부터 물리적으로 해방되어 트리콤을 물어 뜯을 수 있으며 잎을 더 [23]잘 이동시키기 위해 실크 담요를 형성할 수 있습니다.
따끔따끔한 트리콤
침이 있는 트리콤은 형태와 종 간의 분포가 다양하지만, 큰 초식 동물에 대한 비슷한 영향은 그들이 비슷한 기능을 한다는 것을 암시합니다.초본에 취약한 지역에서는 더 높은 밀도의 날카로운 트리콤이 관찰되었다.우르티카에서, 찌르는 듯한 트리콤은 사람이 접촉했을 때 몇 시간 동안 고통스러운 감각을 유발합니다.이 감각은 큰 동물과 작은 무척추동물에 대한 방어 메커니즘으로 작용하며 대사물의 분비를 통한 방어 보충에 역할을 한다.연구에 따르면 이 감각은 접촉 및 해당 트리콤의 [24]구상 끝을 통한 침투 시 독소(히스타민 등)의 빠른 방출을 수반한다.
「 」를 참조해 주세요.
레퍼런스
- ^ "Identify That Alga". Retrieved September 20, 2013.
- ^ Payne, Willard W. (1978). "A Glossary of Plant Hair Terminology". Brittonia. 30 (2): 239–255. doi:10.2307/2806659. ISSN 0007-196X. JSTOR 2806659. S2CID 42417527.
- ^ Levin, Donald A. (1973). "The Role of Trichomes in Plant Defense". The Quarterly Review of Biology. 48 (1): 3–15. doi:10.1086/407484. ISSN 0033-5770. JSTOR 2822621. S2CID 83358711.
- ^ a b c d Tattini, M.; Matteini, P.; Saracini, E.; Traversi, M. L.; Giordano, C.; Agati, G. (2007). "Morphology and Biochemistry of Non-Glandular Trichomes in Cistus salvifolius L. Leaves Growing in Extreme Habitats of the Mediterranean Basin". Plant Biology. 9 (3): 411–419. doi:10.1055/s-2006-924662. ISSN 1438-8677. PMID 17143807.
- ^ Hülskamp, Martin (June 2004). "Plant Trichomes: A Model for Cell Differentiation". Molecular Cell Biology. 5: 471–480. PMID 15173826 – via EBSCO.
- ^ Tholl, Dorothea (2015), "Biosynthesis and Biological Functions of Terpenoids in Plants", in Schrader, Jens; Bohlmann, Jörg (eds.), Biotechnology of Isoprenoids, Advances in Biochemical Engineering/Biotechnology, vol. 148, Springer International Publishing, pp. 63–106, doi:10.1007/10_2014_295, ISBN 9783319201078, PMID 25583224
- ^ Jasiński, Michał; Biała, Wanda (2018). "The Phenylpropanoid Case – It Is Transport That Matters". Frontiers in Plant Science. 9: 1610. doi:10.3389/fpls.2018.01610. ISSN 1664-462X. PMC 6221964. PMID 30443262.
- ^ Huchelmann, Alexandre; Boutry, Marc; Hachez, Charles (2017). "Plant Glandular Trichomes: Natural Cell Factories of High Biotechnological Interest1[OPEN]". Plant Physiology. 175 (1): 6–22. doi:10.1104/pp.17.00727. ISSN 0032-0889. PMC 5580781. PMID 28724619.
- ^ Schellmann, S.; Schnittger, A.; Kirik, V.; Wada, T.; Okada, K.; Beermann, A.; Thumfahrt, J.; Jürgens, G.; Hülskamp, M. (2002-10-01). "TRIPTYCHON and CAPRICE mediate lateral inhibition during trichome and root hair patterning in Arabidopsis". The EMBO Journal. 21 (19): 5036–5046. doi:10.1093/emboj/cdf524. ISSN 0261-4189. PMC 129046. PMID 12356720.
- ^ Schellmann, Swen; Hulskamp, Martin (2004-09-01). "Epidermal differentiation: trichomes in Arabidopsis as a model system". The International Journal of Developmental Biology. 49 (5–6): 579–584. doi:10.1387/ijdb.051983ss. ISSN 0214-6282. PMID 16096966.
- ^ Dosier, Larry W.; Riopel, J. L. (1977). "Differential Enzyme Activity During Trichoblast Differentiation in Elodea canadensis". American Journal of Botany. 64 (9): 1049–1056. doi:10.2307/2442160. JSTOR 2442160.
- ^ a b c Yuan, Ling; Singh, Sanjay Kumar; Patra, Barunava; Pattanaik, Sitakanta (2014). "An overview of the gene regulatory network controlling trichome development in the model plant, Arabidopsis". Frontiers in Plant Science. 5: 259. doi:10.3389/fpls.2014.00259. ISSN 1664-462X. PMC 4071814. PMID 25018756.
- ^ Davis, P.H.; Heywood, V.H. (1963). Principles of angiosperm taxonomy. Princeton, New Jersey: Van Nostrandpage. p. 154.
- ^ Bhatia, R.Y.P.; Raghavan, S.; Rao, K.V.S.; Prasad, V.N. (1973). "Forensic examination of leaf and leaf fragments in fresh and dried conditions". Journal of the Forensic Science Society. 13 (3): 183–190. doi:10.1016/S0015-7368(73)70794-5. PMID 4774149.
- ^ United Nations Office on Drugs and Crime (2009). Recommended Methods for the Identification and Analysis of Cannabis and Cannabis Products (Revised and updated). New York: United Nations. pp. 30–32. ISBN 9789211482423.
- ^ Maithur, Jaideep; Chua, Nam-Hai (April 2000). "Microtubule Stabilization Leads to Growth Reorientation in Arabidopsis Trichomes" (PDF). Plant Cell. 12 (4): 465–477. doi:10.1105/tpc.12.4.465. PMC 139846. PMID 10760237.
- ^ Hahn, Florian; Mantegazza, Otho; Greiner, André; Hegemann, Peter; Eisenhut, Marion; Weber, Andreas P. M. (2017). "An Efficient Visual Screen for CRISPR/Cas9 Activity in Arabidopsis thaliana". Frontiers in Plant Science. 8: 39. doi:10.3389/fpls.2017.00039. ISSN 1664-462X. PMC 5258748. PMID 28174584.
- ^ Hahn, Florian; Eisenhut, Marion; Mantegazza, Otho; Weber, Andreas P. M. (5 April 2018). "Homology-Directed Repair of a Defective Glabrous Gene in Arabidopsis With Cas9-Based Gene Targeting". Frontiers in Plant Science. 9: 424. doi:10.3389/fpls.2018.00424. PMC 5895730. PMID 29675030.
- ^ Hülskamp, M.; Schnittger, A.; Folkers, U. (1999). "Pattern formation and cell differentiation: trichomes in Arabidopsis as a genetic model system". International Review of Cytology. 186: 147–178. doi:10.1016/S0074-7696(08)61053-0. ISBN 9780123645906. ISSN 0074-7696. PMID 9770299.
- ^ Szyndler, M.W.; Haynes, K.F.; Potter, M.F.; Corn, R.M.; Loudon, C. (2013). "Entrapment of bed bugs by leaf trichomes inspires microfabrication of biomimetic surfaces". Journal of the Royal Society Interface. 10 (83): 20130174. doi:10.1098/rsif.2013.0174. ISSN 1742-5662. PMC 3645427. PMID 23576783.
- ^ Eltz, Thomas; Küttner, Jennifer; Lunau, Klaus; Tollrian, Ralph (6 January 2015). "Plant secretions prevent wasp parasitism in nests of wool-carder bees, with implications for the diversification of nesting materials in Megachilidae". Frontiers in Ecology and Evolution. 2. doi:10.3389/fevo.2014.00086.
- ^ Karley, A (2016). "Exploiting physical defence traits for crop protection: leaf trichomes of Rubus idaeus have deterrent effects on spider mites but not aphids". Annals of Applied Biology. 168 (2): 159–172. doi:10.1111/aab.12252 – via Academic Search Premier.
- ^ Cardoso, Márcio Z (2008). "Ecology, Behavior and Binomics: Herbivore Handling of a Plant's Trichome: The Case of Heliconius Charithonia (L.) (Lepidoptera:Nymphalidae) and Passiflora Lobata (Kilip) Hutch. (Passifloraceae)". Neotropical Entomology. 37 (3): 247–52. doi:10.1590/s1519-566x2008000300002. PMID 18641894.
- ^ Fu, H. Y.; Chen, S. J.; Kuo-Huang, L. L. (2003). "Comparative study on the stinging trichomes and some related epidermal structures in the leaves of Dendrocnide meyeniana, Girardinia diversifolia, and Urtica thunbergiana". Taiwania. 48 (4): 213–223.
참고 문헌
- 에서, K. 1965년식물 해부학, 제2판John Wiley & Sons 767쪽