골수형성
Myelinogenesis
골수형성은 신경계에서 미엘린 피복이 형성되고 발달하는 것으로, 일반적으로 산전 신경 발달에서 늦게 시작되고 산후 [1]발달 내내 지속된다.골수형성은 신경 회로 가소성을 통한 학습과 기억을 지원하기 위해 평생 지속되며,[2] 부상 후 재골수형성 또한 지속된다.축삭의 성공적인 미엘리네이션은 공간적으로 분리된 뇌 영역 간의 적시 신호 전도에 필수적인 염분 전도를 가능하게 하여 활동 전위 속도를 증가시키고 [3]뉴런에 대사 지원을 제공합니다.
스테이지
미엘린은 중추신경계의 올리고덴드로사이트와 말초신경계의 슈반세포에 의해 형성된다.따라서 골수형성의 첫 단계는 종종 올리고덴드로사이트 전구세포 또는 슈반세포 전구세포가 성숙한 [4]세포로 분화된 후 축삭 [5]주위에 미엘린이 형성되는 것으로 정의된다.
올리고덴드로사이트 계통은 골수화 [6]시작과의 관계에 따라 4단계로 더 분류할 수 있다.
- 분화: OPC는 증식, 자가 재생 상태를 벗어나 올리고덴드로사이트의 운명 약속과 관련된 유전자와 단백질을 발현하기 시작합니다.
- 프리올리고덴드로사이트:이 세포들은 O4 항원을 발현하고 특별한 [7]조직 없이 방사상으로 확장되는 여러 과정을 발달시킨다.
- 미성숙한 올리고덴드로사이트:때때로 골수성 올리고덴드로사이트라고 불리는 이 세포들은 축삭과 접촉하고 축삭 신호에 반응하여 골수형성을 시작할 준비가 되도록 신경세포에 고정시키는 "피오니어 과정"을 확장한다.이러한 선구적 과정은 목표 축선을 [7]따라 종방향으로 성장합니다.
- 성숙한 올리고덴드로사이트:미엘린 생성 후 성숙한 올리고덴드로사이트는 미엘린 염기성 단백질(MBP)과 미엘린 단백질(PLP)을 포함한 조직화된 다층 미엘린 피복으로 축삭을 둘러싸고 있다.
따라서 골수형성은 3단계와 [6]4단계 사이의 전환 과정을 포함한다.미엘린 형성이 시작되면 각 개척자 과정은 대상 축삭을 중심으로 확장되고 정교해지는 층상 확장을 형성합니다.이것이 미엘린 [7]칼집의 첫 번째 턴을 형성한다.칼집은 표적 축삭의 길이를 따라 계속 확장되며, 새로운 막은 나선형 단면 구조를 띠기 시작하는 미엘린 칼집의 내설의 앞쪽 가장자리에서 합성됩니다.

막층의 적절한 조립을 촉진하기 위해 PLP를 막에 삽입하여 미엘린막의 외부 리플릿 간의 상호작용을 안정시키고 MBP를 국소적으로 번역하여 세포질막 리플릿에 삽입하여 [8]미엘린막을 내부적으로 강화한다.랑비에르의 축삭절 형성과 함께 미엘린 칼집의 가장자리가 초자연적인 [9]고리를 형성합니다.
메커니즘
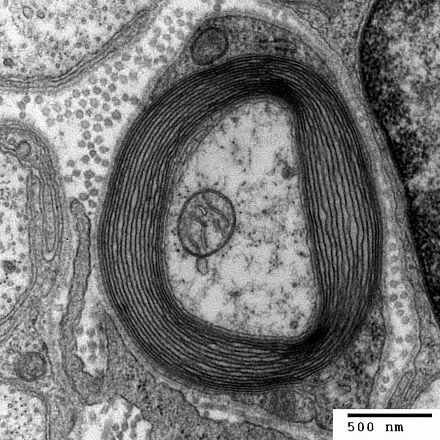

기본 나선-루프-나선 전사인자 OLIG1은 미엘린 관련 유전자의 발현을 조절하여 올리고덴드로사이트 미엘린 형성의 과정에서 필수적인 역할을 한다.OLIG1은 뇌 내 올리고덴드로사이트에 의한 골수화를 개시하기 위해 필요하지만 [10]척수에서는 다소 불필요하다.
축삭에서 파생된 신호는 미엘린 형성의 시작을 조절한다.연구자들은 말초 축삭이 골수화를 시작하도록 올리고덴드로사이트를 자극하는지 여부를 조사하기 위해 28주 동안 PNS 축삭 재생을 연구했다.말초 축삭 재생에 의한 미엘리네이션의 실험적 유도는 슈반 세포와 올리고덴드로사이트가 미엘리네이션 [11]자극에 대한 공유 메커니즘을 가지고 있음을 보여주었다.미엘린 형성의 신경 조절에 대한 증거를 제공하는 유사한 연구는 미엘린 형성이 연관된 [11]축삭의 정의되지 않은 특성에 의해 통제된 슈반 세포에 기인한다고 시사했다.
최근 쥐를 대상으로 한 연구는 아포트랜스페린이 갑상선 호르몬 수용체 [12]알파의 발현을 촉진하기 때문에 아포트랜스페린과 갑상선 호르몬이 분리되고 상승적으로 미엘린 형성을 촉진한다는 것을 시사했다.
말초 미엘린 생성

말초 미엘린 형성은 단백질 P1, P2, P0의 [13]합성에 의해 제어된다.연구진은 SDS-PAGE를 이용해 2만7000개의 달톤(P1), 1만9000개의 달톤(P2), 1만4000개의 달톤(P0)의 밴드 크기를 가진 뚜렷한 밴드를 밝혀냈다.연구들은 또한 P1과 P2가 P0보다 먼저 활성화된다는 것을 보여주었습니다. 왜냐하면 이 단백질은 말초 신경계에서 [13]나오기 때문입니다.재생 과정에서, Schwann 세포는 축삭의 존재가 재정립되면 미엘린 특이 단백질과 관련된 단백질을 재합성한다.미엘린 특이 단백질의 합성은 [13]축삭과 관련된 슈반 세포에서만 일어난다.또한 축삭간 막막 상호작용은 P1, P2, P0의 합성을 촉진하기 위해 필요할 수 있다.
시신경의 골수형성
미엘린 형성의 과정과 역학적 기능은 전통적으로 쥐 시신경의 초미세 구조와 생화학적 기술을 사용하여 연구되어 왔다.이 연구 방법의 실행은 전적으로 미수축으로 구성된 모델 유기체 신경에서 미엘린 형성의 실험적인 관찰을 오랫동안 허용해 왔다.게다가, 쥐 시신경의 사용은 초기 미엘린 발생 연구자들에게 미엘린 [14]발생의 부적절하고 비정형적인 과정에 대한 통찰력을 제공하는데 도움을 주었다.
한 초기 연구는 발달하는 래트 시신경에서 올리고덴드로사이트의 형성과 그에 따른 골수화가 산후 발생한다는 것을 보여주었다.시신경에서 올리고덴드로사이트 세포는 5일 후에 마지막으로 분열되어 미엘린 형성이 6일 또는 7일 전후로 시작되었다.그러나, 올리고덴드로사이트가 미엘린을 생성하기 위해 자극된 정확한 과정은 아직 완전히 이해되지 않았지만, 시신경의 초기 미엘리네이션은 다양한 지질(콜레스테롤, 뇌사면, 술파타이드)[14]의 생산 증가와 관련이 있다.
연구원들이 산후 연구를 시작하면서 쥐 시신경의 골수 형성은 남아 있는 작은 축삭으로 진행되기 전에 가장 큰 축삭에서 시작된다는 것을 발견했습니다.산후 2주차에, 올리고덴드로사이트의 형성은 느려졌다.이 시점에서는 축삭의 15%가 미엘린화되었지만, 미엘린 형성은 계속 급속히 증가했다.생후 4주 동안 쥐의 시신경에 있는 축삭의 거의 85%가 [14]미엘린화되었습니다.5주째와 16주째에 골수화가 느려지고 나머지 골수화되지 않은 축삭은 [15]골수화면에 감열되었다.쥐 시신경을 통해 초기 연구는 골수형성 분야의 지식에 중요한 기여를 했다.
술파티드의 역할
발달 중인 시신경에 대한 연구 결과 갈락토크레브로사이드(술파타이드를 형성하는)가 산후 9일에 나타났고 산후 [14]15일에 최고점에 도달한 것으로 밝혀졌다.이 표현은 시신경이 축삭의 최대 골수화 기간을 보인 시기와 유사했다.축삭골수화의 활성이 감소함에 따라 효소의 활성이 황산염([35S])의 생체내 혼입과 병행한다는 결론을 내릴 수 있었다.
쥐 시신경에 대한 연구는 생후 15일이 골수 감소가 관찰되는 시기라는 것을 밝혀냈다.이 시기 이전에는 축삭의 약 70%가 수액화되지 않았습니다.이때 [35S]황산염이 황산염에 함유되어 뇌브로사이드인 술포전달효소의 활성은 효소 활성에 최고점에 달했다.이 시간대는 또한 생화학 데이터를 [14]기반으로 최대 골수화 기간을 보여주었다.
CNS에서 술파타이드, 황산당단백질 및 황산무코다당류는 미엘린보다는 뉴런과 관련이 있는 것으로 보인다.[35S]에서 생성된 술파타이드와 술포전달효소 활성도를 그래프로 나타내면 [14]피크가 구분된다.피크는 생후 15일에 발생합니다.이러한 피크는 실험 [14]내내 관찰된 시신경의 최대 골수화 기간과 일치합니다.
결론적으로, 골수화의 초기 단계는 지질, 콜레스테롤, 뇌사면 및 술파타이드 [14]합성의 증가와 관련이 있었다.이러한 화합물은 올리고덴드로글리아의 [14]골지 기구에 합성되어 패키징될 가능성이 있다.이러한 지질들의 수송은 알려져 있지 않지만, 그들의 합성 없이 골수화가 지연되는 것으로 보인다.
임상적 의의
미엘린은 일부 신경 세포의 축삭을 둘러싼 전기 절연층을 형성하기 때문에, 탈수 질환은 신경계의 기능에 영향을 미칠 수 있습니다.그러한 질병 중 하나는 중추신경계에서 탈수 현상이 일어나는 다발성 경화증이다.[16]MS에서 [17]올리고덴드로사이트를 보호하고 재분해를 촉진하는 연구가 진행되고 있지만, 현재 치료법은 [18]탈수에서 면역체계의 역할을 주로 다루고 있다.
조사 이력

또 다른 연구원인 폴 플렉시그는 그의 경력의 대부분을 인간의 대뇌피질에서 그 과정의 세부사항을 연구하고 출판하는 데 보냈다.이것은 주로 생후 두 달 사이에 일어납니다.그는 45개의 분리된 피질 부위를 확인했고, 실제로 골수 패턴으로 대뇌피질을 지도화했다.골수화되는 첫 번째 피질 영역은 운동 피질(Brodmann 영역 4의 일부)이고, 두 번째 피질은 후각 피질이고 세 번째 피질은 체질 감각 피질(BA 3,1,2)의 일부입니다.
골수화 마지막 영역은 전측 대상 피질(F#43)과 하측 측두엽 피질(F#44) 및 배측 전전두엽 피질(F#45)이다.
중추신경계의 다른 모든 부분과 마찬가지로 뇌신경계에서도 신경섬유는 모든 곳에서 동시에 발달하는 것이 아니라 일정한 연속성을 가지며, 이 일련의 사건들은 특히 수질물질의 출현과 관련하여 유지된다.대뇌의 회전에서는 태아의 성숙 3개월 전에 이미 수질 물질에 대한 투자(미엘린화)가 시작된 반면, 다른 곳에서는 많은 섬유들이 출생 3개월 후에도 수질 물질이 결여되어 있다.회합의 승계 순서는 척수, 수두 및 중두엽에 유효한 법칙과 동일한 법칙에 의해 관리되며, 이 법칙은 어느 정도 설명될 수 있습니다.즉, 대략적으로 말하면, 동등하게 중요한 신경섬유가 동시에 개발되지만, 유사하지 않습니다.rtance는 명령법칙(골수형성기본법칙)에 의해 정의된 연속적으로 개발된다.수질의 형성은 어떤 경우에는 시작도 안 하고 어떤 경우에는 약간만 [19]진행된 상태에서 거의 완료된다.
레퍼런스
- ^ Eilam, R.; Bar-Lev, D.D.; Levin-Zaidman, S.; Tsoory, M.; LoPresti, P.; Sela, M.; Arnon, R.; Aharoni, R. (2014). "Oligodendrogenesis and myelinogenesis during postnatal development effect of glatiramer acetate". Glia. 62 (4): 649–665. doi:10.1002/glia.22632. PMID 24481644. S2CID 25559134.
- ^ Williamson JM, Lyons DA (2018). "Myelin Dynamics Throughout Life: An Ever-Changing Landscape?". Front Cell Neurosci. 12: 424. doi:10.3389/fncel.2018.00424. PMC 6252314. PMID 30510502.
- ^ Philips T, Rothstein JD (2017). "Oligodendroglia: metabolic supporters of neurons". J Clin Invest. 127 (9): 3271–3280. doi:10.1172/JCI90610. PMC 5669561. PMID 28862639.
- ^ Serrano-Regal MP, Luengas-Escuza I, Bayón-Cordero L, Ibarra-Aizpurua N, Alberdi E, Pérez-Samartín A; et al. (2020). "Oligodendrocyte Differentiation and Myelination Is Potentiated via GABAB Receptor Activation". Neuroscience. 439: 163–180. doi:10.1016/j.neuroscience.2019.07.014. PMID 31349008. S2CID 198934117.
{{cite journal}}: CS1 maint: 여러 이름: 작성자 목록(링크) - ^ Watkins, T., Mulinyawe, S., Emery, B., Barres, B. (2008)골수화의 다른 단계 급속 골수성 CNS 공배양 시스템에서 Y-시크리테아제 및 성세포에 의해 조절된다.555-569
- ^ a b Kinney, H. C., & Volpe, J. J. (2018).골수 이벤트Volpe의 신생아 신경학, 176–188. doi:10.1016/b978-0-323-42876-7.00008-9
- ^ a b c Friedrich, V., Hardy, RJ, (1996년)골수형성 동안 올리고덴드로사이트 프로세스 아버의 점진적 리모델링 243-54
- ^ Baron W, Hoekstra D (2010). "On the biogenesis of myelin membranes: sorting, trafficking and cell polarity". FEBS Lett. 584 (9): 1760–70. doi:10.1016/j.febslet.2009.10.085. PMID 19896485.
- ^ Snaidero N, Möbius W, Czopka T, Hekking LH, Mathisen C, Verkleij D; et al. (2014). "Myelin membrane wrapping of CNS axons by PI(3,4,5)P3-dependent polarized growth at the inner tongue". Cell. 156 (1–2): 277–90. doi:10.1016/j.cell.2013.11.044. PMC 4862569. PMID 24439382.
{{cite journal}}: CS1 maint: 여러 이름: 작성자 목록(링크) - ^ Xin, M. (2005년뇌에서 골수형성과 올리고덴드로사이트에 의한 축삭인식은 올리고1-늘마우스에서 분리된다.신경과학 저널, 1354-1365.doi:10.1523/jneurosci.3034-04.2005
- ^ a b 와인버그, E. & 스펜서, P. (1979년)골수형성의 제어에 관한 연구. 3. 말초 축삭을 재생함으로써 올리고덴드로사이트 골수화의 신호.뇌연구, 162(2), 273-279.도이:10.1016/0006-8993(79)90289-0
- ^ Marziali, L.N., Garcia, C.I., Pasquini, J.M. (2015년)트랜스페린과 갑상선 호르몬은 미엘린 형성의 조절에 수렴한다.실험 신경학.권 265. 129~141.
- ^ a b c Politis, MJ, N. Sternberger, Kathy Ederle, Peter S. Spencer.골수형성의 제어에 관한 연구.신경과학 저널 2.9(1982) : 1252-266.
- ^ a b c d e f g h i 테네쿤, 주, 코헨, SR, 프라이스, DL, 맥칸, GM(1977년).시신경의 골수형성.형태학적, 자동 방사선 촬영 및 생화학적 분석.세포생물학 저널, 72(3), 604-616.
- ^ 단가타, Y, 카우프만, M. (1997년)(C57BL x CBA) F1 잡종 생쥐의 시신경 내 골수형성: 형태계 분석.유럽 형태학 저널, 35(1), 3-18.
- ^ "NINDS Multiple Sclerosis Information Page". National Institute of Neurological Disorders and Stroke. 19 November 2015. Archived from the original on 13 February 2016. Retrieved 6 March 2016.
- ^ Rodgers, Jane M.; Robinson, Andrew P.; Miller, Stephen D. (2013). "Strategies for protecting oligodendrocytes and enhancing remyelination in multiple sclerosis". Discovery Medicine. 86 (86): 53–63. PMC 3970909. PMID 23911232.
- ^ Cohen JA (July 2009). "Emerging therapies for relapsing multiple sclerosis". Arch. Neurol. 66 (7): 821–8. doi:10.1001/archneurol.2009.104. PMID 19597083.
- ^ Flechsig, Paul (1901-10-19). "Developmental (myelogenetic) localisation of the cerebral cortex in the human subject". The Lancet. 158 (4077): 1028. doi:10.1016/s0140-6736(01)01429-5.