클레바지 (엠브리오)
Cleavage (embryo)발달 생물학에서 갈라지는 것은 수정 후의 초기 배아에서 세포의 분열이다.[1] 많은 종의 지고는 전체적으로는 큰 성장 없이 빠른 세포 주기를 거치며, 원래의 지고테와 같은 크기의 세포 군집을 생성한다. 갈라진 틈에서 파생된 다른 세포들은 블라스토메레스라고 불리며 모룰라라고 불리는 콤팩트한 덩어리를 형성한다. 갈라지는 것은 발파선의 형성으로 끝난다.
알의 노른자 농도에 따라 갈라지는 부분이 홀로블라스틱(전체 또는 전체 갈라짐) 또는 메로블라스틱(부분 갈라짐)일 수 있다. 노른자 농도가 가장 높은 달걀의 장대를 식물성 장대라고 하고, 반대로 동물 장대라고 한다.
갈라지는 세포질량을 늘리지 않고 세포와 핵질량을 증가시킨다는 점에서 다른 형태의 세포분열과는 차이가 있다. 이것은 각각의 연속적인 분열이 있을 때, 각 딸 세포에는 그 분열이 일어나기 전보다 약 절반의 세포질이 존재하며, 따라서 핵 대 세포질 물질의 비율이 증가한다는 것을 의미한다.[2]
메커니즘
급속한 세포 주기는 사이클린과 관련 사이클 의존 키나제(cdk)와 같은 세포 주기 진행을 제어하는 단백질을 높은 수준으로 유지함으로써 촉진된다. 복합체 Cyclin B/CDK1 a.k. MPF(만성 촉진 인자)는 유사분열로의 진입을 촉진한다.
카리오키네시스(mitosis)와 사이토키네시스(cytokinesis)의 과정은 함께 작용하여 갈라지는 결과를 낳는다. 이 미토틱 기구는 미관이라고 불리는 튜불린 단백질의 중합체로 이루어진 중심축과 극성 아스터로 구성되어 있다. 아스터는 센트로솜에 의해 핵화되고 센트로솜은 정자가 난자에 기초체로서 가져온 센트리올에 의해 조직된다. 시토키네시스(Cytokinesis)는 마이크로필라멘트라고 불리는 액틴 단백질의 폴리머로 구성된 수축고리에 의해 매개된다. 카리오키네시스(Karyokinesis)와 사이토키네시스(cytokinesis)는 독립적이지만 공간적으로 그리고 일시적으로 조정된 과정이다. 사이토키네시스(cytokinesis)가 없을 때 유사분열이 발생할 수 있는 반면, 사이토키네시스(cytokinesis)는 유사분열체를 필요로 한다
갈라짐의 끝은 혈전사의 시작과 일치한다. 이 점을 중간블라스툴라 전이(midblastula transition)라고 하며 핵:사이토플라즘 비율(약 1/6)에 의해 제어되는 것으로 보인다.
갈라짐의 종류
결정하다
결정체 갈라짐(모자이크 갈라짐이라고도 함)은 대부분의 양성자에 있다. 그것은 배아발달 초기에 세포의 발달운명을 초래한다. 초기 배아 갈라짐으로 생성된 각각의 발파체는 완전한 배아로 발전할 수 있는 능력을 가지고 있지 않다.
미확정
세포는 방해받지 않는 동물/야생동물 사이토아키텍처적 특징의 완전한 세트를 가지고 있는 경우에만 불분명할 수 있다(규정이라고도 한다). 그것은 중수체 태아의 원래 세포가 분열할 때, 두 개의 결과 세포가 분리될 수 있고, 각각의 세포가 개별적으로 전체 유기체로 발전할 수 있다.
홀로블라스틱
홀로블라스틱 갈라짐에서 지고테와 블라토메르는 갈라짐 동안 완전히 나뉘기 때문에 블라토메르의 수는 갈라짐마다 두 배가 된다. 노른자 농도가 높지 않은 경우 4가지 주요 갈라짐 유형을 이졸시탈세포(노른자 분포가 작은 세포) 또는 중졸시탈세포 또는 미세시탈세포(경사로에서 노른자의 모데레이트 농도)에서 관찰할 수 있다 – 쌍방향 홀로블라스틱, 방사형 홀로블라스틱, 회전홀로블라스틱, 나선홀로블라스틱, 나선홀로블라스틱, 갈라진 [3]틈 이 홀로블라스틱 갈라진 평면은 사이토키네시스 과정에서 이솔레시탈 지고트를 통과한다. 코엘로블라술라는 이러한 방사상 갈라진 난자를 위한 다음 단계의 발전이다. 홀로블라스틱한 달걀에서 첫 번째 갈라짐은 항상 알의 식물-동물 축을 따라 발생하며, 두 번째 갈라짐은 첫 번째 갈라짐과 수직이다. 여기서 발파체의 공간 배치는 다양한 유기체에서 각기 다른 갈라짐 평면으로 인해 다양한 패턴을 따를 수 있다.
쌍방의
- 첫 번째 갈라짐은 zygote를 왼쪽과 오른쪽 반으로 분리하는 결과를 낳는다. 다음의 갈라진 평면은 이 축을 중심으로 하여 두 반쪽이 서로 거울에 비친 이미지가 된다. 양쪽의 홀로블라스틱 갈라짐에서, 블라스토메르의 분할은 완전하고 분리되어 있다; 블라스토메르가 부분적으로 연결되어 있는 양쪽의 메로블라스틱 갈라짐과 비교된다.
방사상
회전
- 회전 갈라짐은 경혈축을 따라 정상적인 1분할을 수반하며, 두 개의 딸세포가 생긴다. 이 갈라짐이 다른 방법은 딸세포 중 하나는 경맥으로, 다른 하나는 적도로 갈라지는 것이다.
나선형
- 나선 갈라짐은 스피랄리아라고 불리는 로포트로초안 세사의 많은 구성원들 사이에 보존되어 있다.[5] 일부 나선은 불평등한 갈라짐(아래 참조)[6]을 겪지만 대부분의 나선형 갈라짐은 동일하다. 이 소분류는 안네리스, 연체동물, 시펀쿨라를 포함한다. 나선형 갈라짐은 종마다 다를 수 있지만, 일반적으로 처음 두 개의 세포 분열은 각각 배아의 사분면을 나타내는 4개의 마크로미(A, B, C, D)라고도 한다. 이러한 처음 두 개의 갈라짐은 zygote의 동물-식물 축에 평행하게 직각으로 발생하는 평면에서 방향을 잡지 않는다.[5] 4세포 단계에서 A와 C마크롬은 동물극에서 만나 동물성 크로스퍼로를 만들고, B와 D마크롬은 식물성 폴에서 만나 식물성 크로스퍼로를 만든다.[7] 각각의 연속적인 갈라짐 사이클과 함께, 마크롬은 동물 기둥에 더 작은 마이크로미크롬 4중주단을 발생시킨다.[8][9] 이러한 사중주를 생성하는 분할은 동물성 채식축에 대해 90°의 배수가 아닌 각도인 비스듬한 각도에서 발생한다.[9] 각각의 4중주 마이크로롬은 그들의 모체 마크로미레에 비례하여 회전하며, 이 회전의 치례성은 홀수와 짝수 사중주단 사이에 서로 다른데, 이는 홀수와 짝수 사중주단 사이에 교대칭이 존재함을 의미한다.[5] 즉, 각 4중주를 생산하는 분절의 방향은 동물극에 대해 시계방향과 시계반대방향 사이에서 번갈아 나타난다.[9] 4중주단이 생성될 때 발생하는 교차 분할 패턴은 4개의 매크롬의 갈라진 고랑에 존재하는 마이크로롬 4중주를 생성한다.[7] 동물의 극에서 볼 때, 이 세포 배열은 나선형 패턴을 보여준다.

- D 마크로미어의 사양은 스파이럴 개발의 중요한 측면이다. 1차 축인 동물성 채소는 외생성 중에 결정되지만 2차 축인 등축은 D 사분면의 사양에 의해 결정된다.[9] D 마크로미어는 다른 3개의 마크로미엄이 생산하는 것과 다른 세포분열을 용이하게 한다. D 사분면의 세포는 나선형의 등쪽과 후측 구조를 일으킨다.[9] D 사분면을 지정하기 위해 알려진 두 가지 메커니즘이 있다. 이러한 메커니즘은 균등 분열과 불평등한 분열을 포함한다.
- 균등 분할에서 처음 두 개의 세포 분열은 서로 구별할 수 없는 4개의 매크롬을 생성한다. 각각의 마크로미어는 D 마크로미어가 될 가능성이 있다.[8] 제3 사중주단이 형성된 후, 마크로미어 중 한 마크로미어는 배아의 동물극에 있는 오버로잉 마이크로미럴과 최대한의 접촉을 시작한다.[8][9] 이 접촉은 하나의 마크로미어를 공식 D 사분면 폭발물로 구별하기 위해 필요하다. 똑같이 갈라지는 나선형 배아에서 D 사분면은 마이크로롬과의 접촉이 하나의 세포가 미래의 D 블라썸머가 될 것을 지시할 때 3번째 4중주단이 형성될 때까지 지정되지 않는다. 일단 지정되면, D 블라스토메르는 주변의 마이크로롬에 신호를 보내 세포 운명을 결정하게 된다.[9]
- 불평등한 갈라짐에서, 처음 두 개의 세포분열은 다른 세 개의 세포보다 하나의 세포가 더 큰 네 개의 세포를 생산하지 못한다. 이 더 큰 셀은 D 마크로미어로 지정된다.[8][9] 균등하게 갈라지는 나선형과 달리 D 마크로미어는 불평등한 갈라짐 동안 4세포 단계에서 지정된다. 불평등한 갈라짐은 두 가지 방법으로 발생할 수 있다. 한 가지 방법은 균열 스핀들의 비대칭 배치를 포함한다.[9] 이것은 한 극에 있는 별표가 세포막에 부착되어 다른 극에 있는 별자리보다 훨씬 작을 때 발생한다.[8] 이것은 두 마크로미어가 난자의 동물 영역의 일부를 물려받지만, 더 큰 마크로미어만이 식물성 영역을 물려받는 불평등한 사이토키네시스 현상을 초래한다.[8] 불평등한 갈라짐의 두 번째 메커니즘은 극성 로브라고 불리는 세포질, 막 결합, 세포질 돌출부의 생산을 포함한다.[8] 이 극성엽은 갈라지는 동안 식물성 극지방에서 형성되고, 그리고 나서 D 블라스토메르로 옮겨진다.[7][8] 극지엽에는 식물성 세포질(cytoplasm)이 들어 있는데, 이것은 미래의 D 마크로미아에 의해 유전된다.[9]

메로블라스틱
수정란 세포에 노른자가 많이 농축되어 있을 때, 세포는 부분적 또는 메로블라스틱한 갈라짐 현상을 겪을 수 있다. Meroblastic cleavage의 두 가지 주요한 형태는 원반과 피상적이다.[10][unreliable source?]
- 원반체의
- 원소형 갈라짐에서 갈라진 고랑들은 노른자를 관통하지 않는다. 배아는 노른자위에 발파성이라고 불리는 세포의 원반을 형성한다. 원소형 갈라짐은 말단소립 알세포(노른자가 한쪽 끝에 집중된 알세포)를 가진 단조류, 새, 파충류, 어류에서 흔히 발견된다. 불완전하게 분열되어 노른자와 접촉하는 세포층을 "합성층"이라고 한다.
- 피상적인
- 피상적인 갈라짐에서 유사분열은 발생하지만 사이토카인증은 발생하지 않아 다핵세포가 발생한다. 노른자가 난자 세포의 중앙에 위치하게 되면 핵은 난자의 주변부로 이동하게 되고, 플라즈마 막은 안쪽으로 자라 핵은 개별 세포로 분할된다. 피상적인 갈라짐은 중추성 난자 세포(노른자가 세포 중앙에 위치한 난자 세포)가 있는 절지동물에 발생한다. 이러한 종류의 갈라짐은 드로소필라와 같이 발달 시기에 동시성을 촉진하는 데 효과가 있을 수 있다.[11]
| I. 홀로블라스틱(완전) 갈라짐 | II. 메로블라스틱(완전하지 않은) 갈라짐 |
|---|---|
| A. 이솔레시탈(스파스, 균등하게 분포된 노른자)
B. 중졸레시탈(모더레이트 식물성 노른자 | A. 텔로레시탈(대부분의 셀에 걸쳐 있는 질소 노른자) B. 센트로레시탈(알의 중앙에 있는 얄크)
|
태반
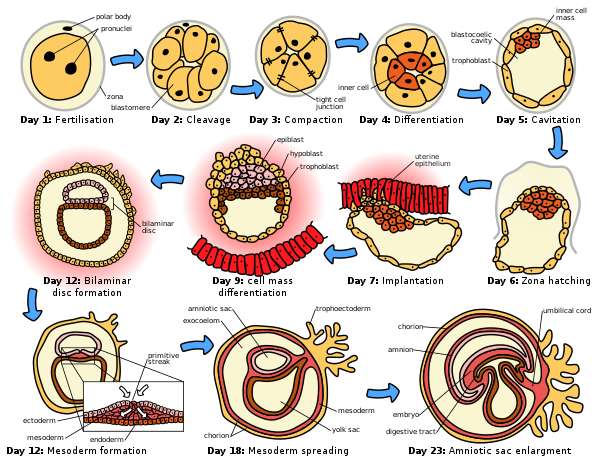
태반 포유류의 갈라짐과 다른 동물의 갈라짐 사이에는 차이가 존재한다.
포유류들은 12시간에서 24시간 사이의 느린 분열 속도를 가지고 있다. 이 세포 분열은 비동기적이다. 축약 전사는 2세포, 4세포 또는 8세포 단계에서 시작한다. 쇄골은 홀로블라스틱하고 회전적이다. 인간은 동등한 분열을 가진 홀로블라스틱 분열증을 가지고 있다.
8세포 단계에서, 배아는 세 번의 갈라짐 과정을 거친 후에 약간의 변화를 겪는다. 이 단계에서 세포들은 압축이라고 알려진 과정에서 단단하게 달라붙기 시작한다.[14][15] 최근, 태반 포유류에서 세포는 압축된 배아 내의 위치에 따라 내부 세포질량 또는 대류 세포질 중 하나에 더 많이 기여할 가능성이 있다고 제안되었다. 단일 세포는 사전 응축 8-세포 배아에서 제거되어 유전자 검사에 사용될 수 있으며 배아는 회복될 것이다.[16]
이 단계에서 대부분의 블라토메르들은 양극화되어 다른 블라토메르와 밀접한 결합을 형성한다. 이 과정은 세포의 다른 두 집단의 발달로 이어진다: 바깥쪽의 극세포와 안쪽에 있는 무극세포. 영양성분세포라 불리는 외세포는 외부에서 나트륨을 펌핑하는데, 이 나트륨과 함께 물을 자동으로 기저(내부) 표면에 유입시켜 공동화라고 하는 과정에서 블라스토콜 공동이 형성된다. 이 영양성분 세포는 결국 초리온이라고 불리는 태반에 배아적 기여를 하게 될 것이다. 내세포는 내세포질량(ICM)을 형성하기 위해 (배아가 더 커지지 않기 때문에) 충동의 한쪽으로 밀려나며, 배아와 일부 외피막들을 발생시킬 것이다. 이 단계에서 배아는 배반포성이라고 불린다.
참고 항목
참조
- ^ Gilbert, Scott F. (2000). "An Introduction to Early Developmental Processes". Developmental Biology (6th ed.). ISBN 978-0878932436.
- ^ Forgács, G.; Newman, Stuart A. (2005). "Cleavage and blastula formation". Biological physics of the developing embryo. Biological Physics of the Developing Embryo. Cambridge University Press. p. 27. Bibcode:2005bpde.book.....F. doi:10.2277/0521783372. ISBN 978-0-521-78337-8.
- ^ Gilbert, Scott F. (2000). "Early Development of the Nematode Caenorhabditis elegans". Developmental Biology (6th ed.). ISBN 978-0878932436. Retrieved 2007-09-17.
- ^ Gilbert SF (2016). Developmental biology (11th ed.). Sinauer. p. 268. ISBN 9781605354705.
- ^ a b c Shankland, M.; Seaver, E. C. (2000). "Evolution of the bilaterian body plan: What have we learned from annelids?". Proceedings of the National Academy of Sciences. 97 (9): 4434–7. Bibcode:2000PNAS...97.4434S. doi:10.1073/pnas.97.9.4434. JSTOR 122407. PMC 34316. PMID 10781038.
- ^ Henry, J. (2002). "Conserved Mechanism of Dorsoventral Axis Determination in Equal-Cleaving Spiralians". Developmental Biology. 248 (2): 343–355. doi:10.1006/dbio.2002.0741. PMID 12167409.
- ^ a b c Boyer, Barbara C.; Jonathan, Q. Henry (1998). "Evolutionary Modifications of the Spiralian Developmental Program". Integrative and Comparative Biology. 38 (4): 621–33. doi:10.1093/icb/38.4.621. JSTOR 4620189.
- ^ a b c d e f g h Freeman, Gary; Lundelius, Judith W. (1992). "Evolutionary implications of the mode of D quadrant specification in coelomates with spiral cleavage". Journal of Evolutionary Biology. 5 (2): 205–47. doi:10.1046/j.1420-9101.1992.5020205.x. S2CID 85304565.
- ^ a b c d e f g h i j Lambert, J.David; Nagy, Lisa M (2003). "The MAPK cascade in equally cleaving spiralian embryos". Developmental Biology. 263 (2): 231–41. doi:10.1016/j.ydbio.2003.07.006. PMID 14597198.
- ^ "Current Notes". Retrieved 2007-09-17.
- ^ 길버트 SF. 개발 생물학 11판. 선덜랜드 (MA): 시나워 어소시에이츠; 2014. 인쇄하다
- ^ Gilbert SF (2003). Developmental biology (7th ed.). Sinauer. p. 214. ISBN 978-0-87893-258-0.
- ^ Kardong, Kenneth V. (2006). Vertebrates: Comparative Anatomy, Function, Evolution (4th ed.). McGraw-Hill. pp. 158–64.
- ^ Nikas, G; Ao, A; Winston, RM; Handyside, AH (July 1996). "Compaction and surface polarity in the human embryo in vitro". Biology of Reproduction. 55 (1): 32–7. doi:10.1095/biolreprod55.1.32. PMID 8793055.
- ^ Nikas G, Ao A, Winston RM, Handyside AH (July 1996). "Compaction and surface polarity in the human embryo in vitro" (PDF). Biol. Reprod. 55 (1): 32–7. doi:10.1095/biolreprod55.1.32. PMID 8793055.
- ^ Wilton, L (2005). "Preimplantation genetic diagnosis and chromosome analysis of blastomeres using comparative genomic hybridization". Human Reproduction Update. 11 (1): 33–41. doi:10.1093/humupd/dmh050. PMID 15569702.
참고 문헌 목록
- Wilt, F.; Hake, S. (2004). Principles of Developmental Biology. ISBN 9780393974300.
- Scott F. Gilbert (2003). Developmental Biology.
- Scott F. Gilbert (2016). Developmental Biology.
추가 읽기
- Valentine, James W. (1997). "Cleavage Patterns and the Topology of the Metazoan Tree of Life". Proceedings of the National Academy of Sciences of the United States of America. 94 (15): 8001–5. Bibcode:1997PNAS...94.8001V. doi:10.1073/pnas.94.15.8001. PMC 21545. PMID 9223303.
- MadSci Network에서 중수체 배아 패턴을 개발할 때의 '장점'은 무엇인가?
- Lee, Seung-Cheol; Mietchen, Daniel; Cho, Jee-Hyun; Kim, Young-Sook; Kim, Cheolsu; Hong, Kwan Soo; Lee, Chulhyun; Kang, Dongmin; Lee, Wontae; Cheong, Chaejoon (2007). "In vivo magnetic resonance microscopy of differentiation in Xenopus laevis embryos from the first cleavage onwards". Differentiation. 75 (1): 84–92. doi:10.1111/j.1432-0436.2006.00114.x. PMID 17244024.