디스코이데움
Dictyostelium discoideum| 디스코이데움 | |
|---|---|
 | |
| D. discoideum 열매체 | |
| 경계가 곡률로 색칠된 이동 D. 디스코이디움, 스케일바: 5µm, 지속시간: 22초 | |
| 과학적 분류 | |
| 도메인: | 진핵생물 |
| 문: | 아메보조아목 |
| 클래스: | 딕티오스테리아속 |
| 주문: | 딕티오스텔리다 |
| 패밀리: | 딕티오스텔리아과 |
| 속: | 딕티오스텔리움 |
| 종류: | 디스코이디움 |
| 이항명 | |
| 디스코이데움 | |

디스코이데움 디스코이데움(Dictyostelium discoideum)은 아메보조아문(Mycetozoa)에 속하는 흙에 사는 아메바이다.흔히 슬라임 곰팡이로 불리는 디스코이디움은 단세포 아메바 집합에서 다세포 민달팽이로, 그리고 평생 동안 열매를 맺는 체로 변하는 진핵생물이다.독특한 무성의 라이프 사이클은 식물성, 집합성, 이주, 절정의 4단계로 구성되어 있습니다.D. discoideum의 라이프 사이클은 비교적 짧기 때문에 모든 스테이지를 시기적절하게 볼 수 있습니다.라이프 사이클에 관여하는 세포는 인간의 암 연구에 적용할 수 있는 움직임, 화학적 신호 전달 및 발달을 거친다.D. discoideum은 라이프사이클의 단순성으로 인해 다른 [2]유기체의 유전적, 세포적, 생화학적 과정을 연구할 수 있는 귀중한 모델 유기체가 됩니다.
자연 서식지와 식생활
야생에서, 디스코이디움은 흙과 촉촉한 나뭇잎 더미에서 발견됩니다.그것의 주요 식단은 토양에서 발견되는 대장균과 부패하는 유기물로 구성되어 있다.D. discoideum의 무핵 아메바는 낙엽성 숲 토양과 썩은 [3]잎을 포함한 자연 서식지에서 발견되는 박테리아를 소비한다.
라이프 사이클과 재생산
D. discoideum의 라이프 사이클은 성숙한 소로카르프(열매체)에서 포자가 배출되면서 시작된다.점사모배는 따뜻하고 습한 환경에서 포자에서 부화한다.그들의 생식 단계에서, myxamoebe는 박테리아를 먹이로 하기 때문에 유사분열로 분열합니다.그 박테리아는 엽산을 분비하여 myxamoebea를 끌어당긴다.균의 공급이 고갈되면 myxamoebe는 응집 단계로 들어간다.
응집하는 동안, 기아는 당단백질과 아데닐릴 사이클라아제와 [4]같은 단백질 화합물의 생산을 시작합니다.당단백질은 세포간 유착을 허용하고 아데닐환원효소는 고리형 AMP를 생성한다. 고리형 AMP는 아메배에서 분비되어 인접한 세포를 중앙 위치로 끌어들인다.신호를 향해 움직일 때, 그들은 당단백질 접착 분자의 사용에 의해 서로 부딪히고 함께 붙는다.
이동 단계는 아메바가 단단한 골재를 형성하고 길쭉한 세포 더미가 땅에 평평하게 누우면 시작됩니다.아메배는 민달팽이로도 알려진 운동성 유사플라스모듐으로 함께 작용합니다.민달팽이의 길이는 약 2-4mm이고, 최대 100,000개의 [5]세포로 구성되며 민달팽이가 이동하는 [6]앞 세포에서 셀룰로오스 피복을 만들어 이동시킬 수 있다.이 칼집의 일부는 빛, 열, 습도 등의 유인물을 향해 전진 전용 [6]방향으로 이동하기 때문에 끈적끈적한 자국으로 남겨집니다.고리형 AMP와 분화유도인자라고 불리는 물질은 다른 종류의 [6]세포를 형성하는데 도움을 준다.민달팽이는 앞부분과 뒷부분으로 각각 이동하는 전엽세포와 전엽세포로 분화된다.일단 적절한 환경을 발견하면, 민달팽이의 앞쪽 끝은 자실체의 줄기를 형성하고, 뒤쪽 끝은 자실체의 [6]포자를 형성한다.최근에야 발견된 앞부분과 같은 세포들도 민달팽이의 뒷부분 전체에 분산되어 있다.이 앞부분과 같은 세포들은 결실체의 맨 밑부분과 [6]포자의 캡을 형성합니다.민달팽이가 한 곳에 자리를 잡은 후, 앞부분이 공중으로 올라가면서 펼쳐져 소위 "멕시코 모자"라고 불리는 것을 형성하고, 절정기가 시작된다.
프리톡 세포와 프리포어 세포는 성숙한 결실체를 [6]형성하기 위해 절정 단계에서 위치를 바꾼다.멕시코 모자의 앞쪽 끝은 셀룰로오스 관을 형성하는데, 이것은 더 많은 뒤쪽 세포가 튜브의 바깥을 위로 이동하도록 하고, 그리고 프리토크 세포는 아래로 [6]이동한다.이 재배열은 슬러그의 전단에서 세포로 이루어진 자실체의 줄기를 형성하고, 슬러그의 후단에서 세포는 꼭대기에 있어 자실체의 포자를 형성한다.이 8~10시간의 과정이 끝나면 성숙한 결실체가 완전히 [6]형성된다.이 자실체는 높이가 1~2mm이고, 이제 myxamoebea가 되는 성숙한 포자를 방출함으로써 전체 주기를 다시 시작할 수 있습니다.
성적 생식
일반적으로 D. discoideum은 일반적으로 무성생식을 하지만, D. discoideum은 특정 조건이 충족되면 여전히 성생식을 할 수 있다.D. discoideum에는 세 가지 다른 짝짓기 유형이 있으며, 연구는 이 세 가지 짝짓기 유형을 규정하는 성흔적을 확인했습니다.1형 균주는 MatA라고 불리는 유전자에 의해 특정되고, 2형 균주는 MatB, Mat C, Mat D의 [7]세 가지 다른 유전자를 가지고 있고, 3형 균주는 Mat S와 Mat T의 유전자를 가지고 있다.이 성별들은 오직 두 개의 다른 성별과 짝짓기를 할 수 있고 그들 [7]자신의 성별과는 짝짓기를 할 수 없다.
그들의 박테리아 먹이 공급과 함께 배양될 때, 이질 또는 동질성 성 발달이 일어나 이배체 접합자가 [8][9]형성될 수 있다.헤테로탈 교배는 서로 다른 교배 유형의 두 개의 아메바가 어둡고 습한 환경에서 존재하며, 이 아메배는 집적 중에 융합하여 거대한 접합자 세포를 형성할 수 있습니다.그런 다음 거대 셀은 cAMP를 방출하여 다른 셀을 유인한 후 다른 셀을 집약체로 흡수합니다.소비된 세포는 골재를 보호하기 위해 두꺼운 셀룰로오스 벽으로 전체 골재를 감싸는 역할을 합니다.이것은 거시낭으로 알려져 있다.거대 세포는 먼저 감수분열 후 유사분열을 통해 분열되어 일반 아메배처럼 먹이를 주기 위해 방출되는 많은 반수상 아메배를 생산한다.호모탈릭 D. 디스코이디움 균주 AC4와 ZA3A도 매크로 [10]낭종을 생성할 수 있다.이 변종들은 이질적인 변종과 달리 결합형 대립 유전자(matA와 mata)를 발현할 가능성이 높다.성적인 번식은 가능하지만, 실험실 환경에서 D. discoideum macrocyst의 발아에 성공하는 것은 매우 드문 일이다.그럼에도 불구하고, 재조합은 D. discoideum 자연 집단 내에서 널리 퍼져 있으며, 이는 성관계가 그들의 [9]라이프사이클에서 중요한 측면일 가능성이 높다는 것을 보여준다.
모델 유기체로 사용
그것의 많은 유전자들이 인간의 유전자와 상동성이지만, 그것의 수명은 단순하기 때문에, 디스코이디움은 모델 유기체로 흔히 사용된다.그것은 주로 세포 유형 및 행동의 제한된 수와 빠른 [6]성장 때문에 유기체, 세포 및 분자 수준에서 관찰될 수 있습니다.그것은 모두 정상적인 세포 과정인 세포 분화, 화학축성, 그리고 세포자멸을 연구하는데 사용된다.그것은 또한 세포 분류, 패턴 형성, 식세포증, 운동성, 신호 [11]전달을 포함한 발달의 다른 측면을 연구하는 데 사용된다.이러한 과정과 발달의 양상은 다른 모델 유기체에서는 보이지 않거나 너무 어렵다.D. discoideum은 고등 메타조와 밀접한 관련이 있다.그것은 비슷한 유전자와 경로를 가지고 있어서 유전자 [12]녹아웃의 좋은 후보가 된다.
세포 분화 과정은 세포가 다세포 유기체로 발달하기 위해 더 전문화 될 때 일어난다.크기, 모양, 대사 활동 및 반응성의 변화는 유전자 발현 조절의 결과로 발생할 수 있습니다.이 종에서 세포의 다양성과 분화는 줄기 세포나 포자 [13]세포로 가는 경로에서의 세포-세포 상호작용에서 이루어지는 결정을 포함한다.이 세포들의 운명은 그들의 환경과 패턴 형성에 달려있다.그러므로, 그 유기체는 세포 분화를 연구하기 위한 훌륭한 모델이다.
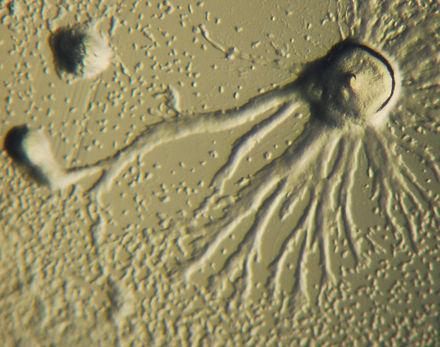
화학성(Chemotaxis)은 화학농도 구배를 따라 화학자극으로 향하거나 화학자극으로부터 멀어지는 유기체의 경로로 정의된다.어떤 생물들은 영양소의 공급으로 이동할 때 화학작용을 보인다.D. discoideum에서, 아메바는 세포 밖으로 신호 cAMP를 분비하여 다른 아메배들이 소스 쪽으로 이동하도록 유인합니다.모든 아메바는 가장 많은 양의 cAMP 분비물을 방출하는 중앙 아메바를 향해 이동합니다.cAMP의 분비는 모든 아메바에 의해 나타나며 집약을 시작해야 합니다.이러한 화학 물질 방출과 아메바 움직임은 6분마다 일어납니다.아메바는 농도 변화 방향으로 60초간 이동한 후 다음 분비물이 나올 때까지 멈춘다.개별 세포의 이러한 행동은 세포 그룹에서 진동을 일으키는 경향이 있으며 다양한 cAMP 농도의 화학파가 [14]: 174–175 나선형으로 그룹을 통해 전파됩니다.
수학 생물학자인 토마스 회퍼와 마틴 보얼리스트에 의해 D. 디스코이디움의 나선과 흐르는 패턴을 재현하는 우아한 수학 방정식 세트가 발견되었습니다.수학 생물학자인 코넬리스 J.Weijer는 유사한 방정식이 그것의 움직임을 모형화할 수 있다는 것을 증명했다.이러한 패턴의 방정식은 주로 아메바 집단의 밀도, 순환 AMP의 생산 속도 및 순환 AMP에 대한 개별 아메바의 감도에 의해 영향을 받는다.나선형 패턴은 군집의 중심에 있는 아메바에 의해 형성되며, 아메바는 순환형 [15][16]AMP의 파동을 보낼 때 회전합니다.
화학작용제로서의 cAMP의 사용은 다른 유기체에서는 확립되지 않았다.발달 생물학에서, 이것은 인간의 염증, 관절염, 천식, 림프구 밀매, 축삭 안내에 대한 이해에 중요한 화학작용의 이해할 수 있는 예 중 하나이다.식세포증은 면역 감시와 항원 제시에서 사용되는 반면, 세포 유형 결정, 세포 분류, 패턴 형성은 이러한 [6]유기체와 함께 연구될 수 있는 배아 형성의 기본 특징이다.
단, cAMP 발진은 다세포 단계에서의 집단 세포 이동에는 필요하지 않을 수 있습니다.연구에 따르면 d. discoideum의 [17]다세포 단계에서 cAMP 매개 신호가 전파에서 정상 상태로 변화합니다.
열축성은 온도 경사를 따라 이동하는 것입니다.슬러그는 0.05°C/cm의 극히 얕은 구배를 따라 이동하는 것으로 나타났지만, 선택한 방향이 복잡하다. 슬러그는 적응한 온도보다 약 2°C 낮은 온도에서 떨어져 있는 것으로 보인다.이 복잡한 행동은 매일의 공기 온도 변화로 인한 토양 온도 변화의 주기적 패턴과 행동을 컴퓨터 모델링으로 분석되었습니다.결론은 이 행동이 민달팽이를 지표면 아래로 몇 센티미터 이동시킨다는 것이다.이것은 [14]: 108–109 겉으로 보기에 중력이 없는 원시 유기체에 의한 놀랍도록 정교한 행동이다.
아포토시스(프로그램화된 세포사망)는 종 [4]발생의 정상적인 부분이다.아포토시스는 복잡한 장기의 적절한 간격과 조각에 필요합니다.D. discoideum 세포 중 약 20%는 성숙한 결실체를 형성하기 위해 이타적으로 자신을 희생한다.그것의 라이프사이클의 유사플라스모듐 단계 동안, 그 유기체는 세 가지 주요 유형의 세포를 형성했다: 프리토크, 프리포어, 그리고 앞부분과 같은 세포.절정 시 프리톡 셀은 셀룰로오스 피막을 분비하여 GREX를 [4]통해 튜브로 확장한다.그들은 분화하면서 포자전세포를 들어올리면서 액포를 형성하고 확대한다.줄기 세포는 아포토시스를 거쳐 포자세포가 기질 위로 높이 올라가면서 죽는다.포자전세포는 포자세포가 되고,[6] 각 세포는 흩어지면 새로운 균사모에바가 된다.이것은 성숙한 결실체인 생식기관의 형성에 아포토시스가 어떻게 사용되는지를 보여주는 예입니다.
딕티오스텔륨 연구의 최근 주요 공헌은 살아있는 [18]세포에서 개별 유전자의 활동을 시각화할 수 있는 새로운 기술로부터 나왔다.이것은 전사가 단순한 확률론적 또는 연속적인 행동을 따르는 것이 아니라 "폭발" 또는 "펄스"(전사 버스트)에서 발생한다는 것을 보여준다.터지는 전사는 이제 박테리아와 인간 사이에 보존된 것으로 보인다.이 유기체의 또 다른 주목할 만한 특징은 인간 세포에서 발견되는 DNA 복구 효소를 가지고 있다는 것이다. 이것은 많은 다른 인기 있는 메타조아 모델 [19]시스템에는 없는 것이다.DNA 수복의 결함은 파괴적인 인간 암으로 이어지기 때문에, 다루기 쉬운 간단한 모델에서 인간 수복 단백질을 연구하는 능력은 매우 귀중한 것으로 입증될 것이다.
실험실 재배
실험실에서[6] 쉽게 배양할 수 있는 이 유기체는 모델 유기체로서의 매력을 더한다.D. discoideum은 액체 배양으로 재배할 수 있지만, 그것은 보통 영양 한천이 함유된 페트리 접시에서 재배되며 표면은 촉촉하게 유지됩니다.배양물은 22~24°C(실온)에서 가장 잘 자란다.D. 디스코이디움은 주로 대장균을 먹이로 하며, 이는 라이프사이클의 모든 단계에 적합합니다.음식 공급이 감소하면, myxamoebea 집적체가 가성플라스모디아를 형성합니다.곧, 그 접시는 라이프 사이클의 다양한 단계들로 덮이게 된다.접시를 체크하는 것은 종종 발전의 상세한 관찰을 가능하게 한다.그 세포들은 어떤 발달 단계에서도 수확될 수 있고 빠르게 자랄 수 있다.
실험실에서 디스코이듐을 배양할 때, 그 행동 반응을 고려하는 것이 중요하다.예를 들어 빛, 높은 온도, 높은 습도, 낮은 이온 농도, pH 구배 산성에 대한 친화력이 있습니다.이러한 매개변수를 조작하여 개발을 어떻게 방해, 중지 또는 가속하는지 확인하기 위해 실험이 종종 수행됩니다.이러한 매개변수의 변화는 문화의 성장 속도와 생존 가능성을 변화시킬 수 있습니다.또한 자실체는 발달의 가장 높은 단계이기 때문에 기류와 물리적 자극에 매우 민감합니다.포자 방출과 관련된 자극이 있는지는 알려지지 않았다.
단백질 발현 연구
딕티오스텔륨의 단백질 발현에 대한 상세한 분석은 다른 발달 단계들 사이의 단백질 발현 프로필의 큰 변화와 딕티오스텔륨 [20]항원에 대한 상업적으로 이용 가능한 일반적인 항체의 부족으로 인해 방해되어 왔다.2013년 스코틀랜드 벳슨 웨스트 암 센터의 한 그룹은 스트렙타비딘 결합체를 [21]사용한 MCC1의 검출을 기반으로 면역 블로팅을 위한 항체 없는 단백질 시각화 표준을 보고했다.
레지오넬라병
레지오넬라속은 사람에게 레지오넬라병을 일으키는 종을 포함한다.디스코이디움은 레지오넬라의 숙주이기도 하며 감염 과정을 [22]연구하기에 적합한 모델이다.구체적으로 D. discoideum은 식세포증, 막 밀매증, 내구증, 소포 분류 및 화학작용을 포함한 레지오넬라 감염과 관련된 유사한 세포골격과 세포과정을 포유류의 숙주 세포와 공유한다.
"농사"
Nature지의 2011년 보고서는 D. discoideum [23][24]식민지의 "원시적인 농업 행동"을 보여주는 연구 결과를 발표했다.D. discoideum과 박테리아 먹이 사이의 "상징증"으로 묘사되는, 야생 채집 D. discoideum 군락의 약 3분의 1이 슬라임 곰팡이 [24]자실체에 박테리아가 포함되었을 때 박테리아의 "남편애"에 관여했다.이 박테리아가 과실체에 결합함으로써 포자가 분산된 위치에서 식량원을 "파종"할 수 있게 되고, 이것은 새로운 지역의 [24]식량 자원이 부족할 때 특히 가치가 있다."농장" 포자에서 생성된 군집 또한 일반적으로 포자를 형성할 때 같은 행동을 보입니다.이 통합에는 다음과 같은 비용이 수반됩니다.모든 먹이 박테리아를 소비하지 않는 군집은 더 작은 포자를 만들어내는데, 포자는 그렇게 넓게 흩어지지 않는다.게다가, 식량이 풍부한 지역에 상륙하는 박테리아를 포함한 포자는 훨씬 더 적은 이점이 있다.이러한 행동 비용과 이익의 균형은 D. discoideum 식민지의 소수가 이러한 관행에 [23][24]관여하는 사실에 기여할 수 있다.
디스코이디움은 그램 양성균뿐만 아니라 그램 음성균을 먹는 것으로 알려져 있지만,[25] 몇몇 인간 병원균을 포함한 식세포화된 박테리아 중 일부는 아메배에서 살 수 있고 세포를 죽이지 않고 빠져나갈 수 있다.그들이 언제 셀에 들어갔는지, 그들이 어디에 있는지, 그리고 언제 셀을 떠났는지 알 수 없습니다.이 연구는 아직 결정적이지는 않지만 농부 복제에 적합한 D. discoideum의 일반적인 라이프 사이클을 그려 이 공생 과정을 더 잘 이해할 수 있다.

사진에서, 사람들은 다른 단계들을 볼 수 있다.우선 기아단계에서 디스코이디움 [25]내에 세균을 봉입하고 아메배 진입 후 리소좀과의 융합을 차단하고 이들 미성형 파고솜을 미토콘드리아, 소포 및 암베에의 거친 내형질망막(RER)에서 유래한 다층막 등의 숙주세포소기관으로 둘러싸인다.세포내 감염에서 RER의 역할은 알려져 있지 않지만,[26] RER은 박테리아에 대한 단백질 공급원으로서 필요하지 않다.박테리아는 집적과 다세포 발달 단계에서 이러한 파고솜 안에 상주한다.아메바는 개성을 보존하고 각각의 아메바는 고유의 박테리아를 가지고 있다.포자가 만들어질 때, 박테리아는 숙주 세포 [27]파괴를 막는 세포 골격 구조의 도움으로 세포에서 소라스로 이동합니다.어떤 결과는 박테리아가 [27]세포를 죽이지 않고 세포 외이식을 이용하는 것을 암시한다.자유생활 아메바는 환경 내 일부 병원균의 지속과 분산을 위해 중요한 역할을 하는 것으로 보인다.레지오넬라 기모필라,[26] 많은 마이코박테리움 종, 프랜시셀라 툴라렌시스, 그리고 대장균을 포함한 많은 다른 박테리아에 대해 아메배와의 일시적인 연관성이 보고되었다.농사는 병원균의 생존에 중요한 역할을 하는 것으로 보인다. 왜냐하면 병원균은 D. 디스코이디움 안에서 살고 복제할 수 있고, 농사를 지을 수 있기 때문이다.네이처의 보고는 아메바 행동에 대한 지식에서 중요한 진전을 이루었고, "당신은 아메바보다 더 멍청하다"는 스페인어로 번역된 유명한 표현은 아메바가 종의 이익을 위해 놀라운 조정과 희생의식을 가진 사회적 행동의 훌륭한 예이기 때문에 감각을 잃어가고 있다.
센티넬 셀
Dictyostelium discoideum의 센티넬 세포는 사회 순환의 슬러그 단계에서 독성 물질을 제거하는 역할을 하는 식세포 세포입니다.일반적으로 둥근 모양이지만, 이 세포들은 자유롭게 순환하는 것으로 알려진 민달팽이 껍질 안에 있습니다.해독 과정은 이 세포들이 식세포증을 통해 민달팽이 속의 독소와 병원균을 삼킬 때 일어난다.그리고 나서, 그 세포들은 5개에서 10개의 세포들로 이루어진 그룹으로 뭉쳐지고, 그 세포들은 민달팽이의 안쪽 칼집에 부착된다.민달팽이가 음식 박테리아를 찾아 새로운 장소로 이동하면서 칼집이 벗겨진다.
센티넬 세포는 총 민달팽이 세포 수의 약 1%를 차지하며, 센티넬 세포 수는 방출되는 동안에도 일정하게 유지됩니다.이것은 독소나 병원균과 함께 제거되는 민달팽이 내 감시 세포가 지속적으로 재생된다는 것을 나타냅니다.제거할 독소나 병원균이 없는 경우에도 민달팽이에는 센티넬 세포가 존재한다.센티넬 세포는 다른 다섯 종의 딕티오스테리아에 위치하고 있는데, 이것은 센티넬 세포가 사회적 아메바에서 [28]선천적인 면역체계의 일반적인 특징으로 묘사될 수 있다는 것을 암시한다.
파밍 상태가 감시 세포에 미치는 영향
감시세포의 수는 야생 D. discoideum의 농업 상태에 따라 달라집니다.브롬화 에티듐의 사용으로 인한 독성 환경에 노출되었을 때, 농가가 비농가에 비해 밀리미터당 감시 세포 수가 낮은 것으로 나타났다.이것은 민달팽이들이 이동하면서 남겨진 흔적을 관찰하고 밀리미터에 존재하는 감시 세포의 수를 세는 것으로 결론을 내렸다.하지만, 감시 세포의 수는 농부들의 포자 생산과 생존력에 영향을 미치지 않습니다.독성이 있는 환경에 노출된 농민은 독성이 없는 환경에서 농부와 동일한 수의 포자를 생산하고, 포자의 생존력은 농가와 비농가 간에 동일했다.Clade 2 Burkholderia(농민 관련 박테리아)를 농가에서 제거했을 때 포자의 생산과 생존력은 비농민들과 비슷했다.따라서, 농가에 의해 운반되는 세균은 독소나 [29]병원균에 의한 잠재적 해악으로부터 농가를 보호하는 추가적인 역할을 할 것을 제안한다.
분류 및 계통발생
더 오래된 분류에서 딕티오스텔륨은 사라진 다계통 분류인 아크라시오균류에 속했다.이것은 세포성 슬라임 곰팡이의 한 종류로, 개별 아메바가 다세포 결실체로 집적되어 있는 것이 특징이며, 아크라시드를 딕티오스테로이드와 [30]연관시키는 중요한 요인이 되었다.
최근의 게놈 연구는 딕티오스텔륨이 식물과 동물보다 더 많은 조상들의 게놈 다양성을 유지해왔다는 것을 보여주었지만, 프로테옴에 기초한 계통 발생학은 아메보조아가 식물과 동물의 [31]분열 이후 동물과 풍뎅이의 혈통에서 갈라졌다는 것을 확인시켜주었다.Dictyosteliidae, order Dictyosteliales는 원생동물, dictyostelid 및 myxogastrid 슬라임 곰팡이를 포함하는 Mycetozoa 내의 단계통 집합체이다.신장인자-1α(EF-1α) 데이터 분석은 mycetozoa를 단통군으로서 지지하지만, rRNA 나무는 mycetozoa를 다계통군으로 배치한다.또, 이러한 데이터는, 딕토스테로이드와 myxogastrid가, 원시 스테로이드보다 보다 밀접하게 관련하고 있다는 생각을 뒷받침한다.EF-1α 분석에서는 또한 미케토조아를 동물-풍뎅이군의 [32]즉각적인 아웃그룹으로 지정했다.최신 계통학 데이터는 점액균과 함께 아메보조아 슈퍼그룹 내에 딕티오스테로이드(dictyostelids)를 확고히 배치한다.한편, 원생동물들은 다계통으로 판명되었고, 그들의 줄기가 있는 열매체는 관련이 없는 여러 [33]혈통의 수렴된 특징입니다.
게놈
D. discoideum 게놈 배열 프로젝트는 2005년 연구소의 국제적인 협업에 의해 완성되어 발표되었습니다.이것은 완전한 염기서열을 가진 최초의 자유생물의 게놈이었다.D. discoideum은 기본 구성이 77% [A+T]인 34-Mb 반배체 게놈으로 구성되며 약 12,500개의 [3]단백질을 코드하는 6개의 염색체를 포함합니다.D. discoideum 게놈의 배열은 세포와 발달 생물학에 대한 더 복잡한 연구를 제공합니다.
트리뉴클레오티드의 연속 반복은 이 게놈에 매우 풍부합니다; 게놈의 한 클래스가 군집되어 있어, 연구자들이 이것이 동원체 역할을 한다고 믿게 합니다.반복은 아미노산의 반복 배열에 대응하고 뉴클레오티드 [3]확장에 의해 확장되는 것으로 생각됩니다.트리뉴클레오티드 반복의 확대는 또한 인간에게서 일어나며, 일반적으로 많은 질병으로 이어진다.디스코이디움 세포들이 어떻게 이러한 아미노산 반복을 견뎌내는지를 배우는 것은 인간이 그것들을 견딜 수 있게 하는 통찰력을 줄 수 있다.
모든 게놈 배열은 시간이 지남에 따라 얻거나 손실된 유전자를 식별하는데 중요한 역할을 한다.비교 유전체 연구는 진핵생물 게놈의 비교를 가능하게 한다.프로테옴에 기초한 계통발생학에서는 아메보조아가 동식물의 [3]분열 후 동식물의 혈통에서 이탈한 것으로 나타났다.디스코이디움 게놈은 많은 암호화된 단백질이 곰팡이, 식물, [3]동물에서 흔히 발견되기 때문에 주목할 만하다.
데이터베이스
- DictyBase - Dictyostelium discoideum에 관한 일반 게놈 데이터베이스
- 막 데이터베이스는 Dictyostelium 및 여러 다른 유기체의 단일 패스 막 통과 단백질에 대한 정보를 제공합니다.
레퍼런스
- ^ Raper, K.B. (1935). "Dictyostelium discoideum, a new species of slime mold from decaying forest leaves". Journal of Agricultural Research. 50: 135–147. Archived from the original on 2017-12-08. Retrieved 2016-01-20.
- ^ Pears, Catherine J.; Gross, Julian D. (2021-03-01). "Microbe Profile: Dictyostelium discoideum: model system for development, chemotaxis and biomedical research". Microbiology. 167 (3). doi:10.1099/mic.0.001040. ISSN 1350-0872. PMID 33646931. S2CID 232092012.
- ^ a b c d e Eichinger L; Noegel, AA (2003). "Crawling in to a new era – the Dictyostelium genome project". The EMBO Journal. 22 (9): 1941–1946. doi:10.1093/emboj/cdg214. PMC 156086. PMID 12727861.
- ^ a b c 길버트 S.F. 2006발달생물학제8회선덜랜드(MA): 시나우어 페이지 36-39.
- ^ Cooper, Geoffrey M (2000). "Chapter 1. An Overview of Cells and Cell Research". The Cell (Work in NCBI Bookshelf). Part I. Introduction (2nd ed.). Sunderland, Massachusetts: Sinauer Associates. Cells As Experimental Models. ISBN 978-0-87893-106-4.
- ^ a b c d e f g h i j k l 타일러 M.S. 2000년발달생물학:실험적인 연구를 위한 가이드입니다.제2판선덜랜드(MA): 시나우어. 페이지 31-34.ISBN 0-87893-843-5
- ^ a b Bloomfield, Gareth; Skelton, Jason; Ivens, Alasdair; Tanaka, Yoshimasa; Kay, Robert R. (2010-12-10). "Sex Determination in the Social Amoeba Dictyostelium discoideum". Science. 330 (6010): 1533–1536. doi:10.1126/science.1197423. ISSN 0036-8075. PMC 3648785. PMID 21148389.
- ^ O'Day DH, Keszei A (May 2012). "Signalling and sex in the social amoebozoans". Biol Rev Camb Philos Soc. 87 (2): 313–29. doi:10.1111/j.1469-185X.2011.00200.x. PMID 21929567. S2CID 205599638.
- ^ a b Flowers JM, Li SI, Stathos A, Saxer G, Ostrowski EA, Queller DC, Strassmann JE, Purugganan MD (July 2010). "Variation, sex, and social cooperation: molecular population genetics of the social amoeba Dictyostelium discoideum". PLOS Genet. 6 (7): e1001013. doi:10.1371/journal.pgen.1001013. PMC 2895654. PMID 20617172.
- ^ Robson GE, Williams KL (April 1980). "The mating system of the cellular slime mould Dictyostelium discoideum". Curr. Genet. 1 (3): 229–32. doi:10.1007/BF00390948. PMID 24189663. S2CID 23172357.
- ^ 딕티베이스, 딕티오스텔리움에 대해서[온라인] (2009년 5월 1일)http://dictybase.org/
- ^ Dlip K.나그, 딕티오스텔륨의 네 가지 키네신 유전자의 파괴.[온라인] (2008년 4월 22일)http://ukpmc.ac.uk/articlerender.cgi?artid=1529371 아카이브에서 2012-07-29 오늘 아카이브 완료
- ^ Kay R.R.; Garrod D.; Tilly R. (1978). "Requirements for cell differentiation in Dictyostelium discoideum". Nature. 211 (5640): 58–60. doi:10.1038/271058a0. PMID 203854. S2CID 4160546.
- ^ a b Dusenbery, David B. (1996). Life at Small Scale. Scientific American Library. New York. ISBN 978-0-7167-5060-4.
- ^ Ian Stewart (November 2000). "Biomathematics Patterns: Spiral Slime. MATHEMATICAL RECREATIONS by Ian Stewart. Finding mathematics in creatures great and small". Scientific American.
- ^ Ian Stewart (2000). What Shape is a Snowflake? [Over sneeuwkristallen en zebrastrepen. De wereld volgens de wiskunde] (in Dutch). Uitgeverij Uniepers; Davidsfonds; Natuur & Techniek. pp. 96–97.
- ^ Ueda, Masahiro; Masato Yasui; Morimoto, Yusuke V.; Hashimura, Hidenori (2019-01-24). "Collective cell migration of Dictyostelium without cAMP oscillations at multicellular stages". Communications Biology. 2 (1): 34. doi:10.1038/s42003-018-0273-6. ISSN 2399-3642. PMC 6345914. PMID 30701199.
- ^ Chubb, JR; Trcek, T; Shenoy, SM; Singer, RH (2006). "Transcriptional pulsing of a developmental gene". Current Biology. 16 (10): 1018–25. doi:10.1016/j.cub.2006.03.092. PMC 4764056. PMID 16713960.
- ^ Hudson J. J.; Hsu D. W.; Guo K.; Zhukovskaya N.; Liu P. H.; Williams J. G.; Pears C. J.; Lakin N. D. (2005). "DNA-PKcs-dependent signaling of DNA damage in Dictyostelium discoideum". Curr Biol. 15 (20): 1880–5. doi:10.1016/j.cub.2005.09.039. PMID 16243037.
- ^ "Immunoblotting: Equality for slime molds!". BioTechniques (paper). 55 (1): 9. July 2013.
- ^ Davidson, Andrew J.; King, Jason S.; Insall, Robert H. (July 2013). "The use of strepavidin conjugates as immunoblot loading controls of mitrochondrial markers for use with Dictyostelium discoidium". Benchmarks. BioTechniques (paper). 55 (1): 39–41. doi:10.2144/000114054. PMID 23834384.
- ^ Bruhn H. 2008년레지오넬라균의 다루기 쉬운 모델 숙주 생물인 딕티오스텔륨.인: Heuner K, Swanson M, 편집자.레지오넬라: 분자 미생물학.Norwich(영국): Caister Academic Press.ISBN 978-1-904455-26-4
- ^ a b "아메바는 이동하면서 원시적인 농업 행동을 보여준다", BBC 뉴스, 2011년 1월 19일
- ^ a b c d Brock DA, Douglas TE, Queller DC, Strassmann JE (20 January 2011). "Primitive agriculture in a social amoeba". Nature. 469 (7330): 393–396. doi:10.1038/nature09668. PMID 21248849. S2CID 4333826.
- ^ a b Clarke, Margaret (2010). "Recent insights into host-pathogen interactions from Dyctiostelium". Cellular Microbiology. 12 (3): 283–291. doi:10.1111/j.1462-5822.2009.01413.x. PMID 19919566.
- ^ a b Molmeret M., Horn, M., Wagner, M., Abu Kwaik, Y (January 2005). "Primitive Amoebae as Training Grounds for Intracellular Bacterial Pathogens". Appl Environ Microbiol. 71 (1): 20–28. doi:10.1128/AEM.71.1.20-28.2005. PMC 544274. PMID 15640165.
{{cite journal}}: CS1 maint: 여러 이름: 작성자 목록(링크) - ^ a b Grant P.Ottom Mary Y.Wu; Margaret Clarke; Hao Lu; O.Roger Anderson; Hubert Hilbi; Howard A. Shuman; Richard H. Kessin (11 November 2003). "Macroautophagy is dispensable for intracellular replication of Legionella pneumophila in Dictyostelium discoideum". Molecular Microbiology. 51 (1): 63–72. doi:10.1046/j.1365-2958.2003.03826.x. PMID 14651611. S2CID 22801290.
- ^ Chen, Guokai; Zhuchenko, Olga; Kuspa, Adam (2007-08-03). "Immune-like Phagocyte Activity in the Social Amoeba". Science. New York, NY. 317 (5838): 678–681. doi:10.1126/science.1143991. ISSN 0036-8075. PMC 3291017. PMID 17673666.
- ^ Brock, Debra A.; Callison, W. Éamon; Strassmann, Joan E.; Queller, David C. (2016-04-27). "Sentinel cells, symbiotic bacteria and toxin resistance in the social amoeba Dictyostelium discoideum". Proc. R. Soc. B. 283 (1829): 20152727. doi:10.1098/rspb.2015.2727. ISSN 0962-8452. PMC 4855374. PMID 27097923.
- ^ Cavender J.C.; Spiegl F.; Swanson A. (2002). "Taxonomy, slime molds, and the questions we ask". The Mycological Society of America. 94 (6): 968–979. PMID 21156570.
- ^ Eichenger L.; et al. (2005). "The genome of the social amoeba Dictyostelium discoideum". Nature. 435 (7038): 34–57. doi:10.1038/nature03481. PMC 1352341. PMID 15875012.
- ^ Baldauf S.L.; Doolittle W.F. (1997). "Origin and evolution of the slime molds (Mycetozoa)". PNAS. 94 (22): 12007–12012. doi:10.1073/pnas.94.22.12007. PMC 23686. PMID 9342353.
- ^ Shadwick, LL; Spiegel, FW; Shadwick, JD; Brown, MW; Silberman, JD (2009). "Eumycetozoa = Amoebozoa?: SSUrDNA Phylogeny of Protosteloid Slime Molds and Its Significance for the Amoebozoan Supergroup". PLOS ONE. 4 (8): e6754. doi:10.1371/journal.pone.0006754. PMC 2727795. PMID 19707546.
추가 정보
- Mary S. Tyler (2000). Developmental Biology: A Guide for Experimental Study.2nd ed. Sinauer Associates. pp. 31–34. ISBN 978-0-87893-843-8.
- Scott F. Gilbert (2006). Developmental Biology. 8th ed. Sinauer. pp. 36–39. ISBN 978-0-87893-250-4.