5α-감소효소
5α-Reductase| 3-옥소-5α-스테로이드 4-수소효소 | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| 식별자 | |||||||||
| EC 번호 | 1.3.1.22 | ||||||||
| CAS 번호. | 9036-43-5 | ||||||||
| 데이터베이스 | |||||||||
| 인텐츠 | IntEnz 뷰 | ||||||||
| 브렌다 | 브렌다 입력 | ||||||||
| 엑스퍼시 | 나이스자이메 뷰 | ||||||||
| 케그 | KEG 입력 | ||||||||
| 메타사이크 | 대사통로 | ||||||||
| 프리암 | 프로필 | ||||||||
| PDB 구조 | RCSB PDB PDBe PDBsum | ||||||||
| 진 온톨로지 | 아미고 / 퀵고 | ||||||||
| |||||||||
| 스테로이드-5α-저감효소, 알파 폴리펩타이드 1 | |||||||
|---|---|---|---|---|---|---|---|
| 식별자 | |||||||
| 기호 | SRD5A1 | ||||||
| 엔씨비유전자 | 6715 | ||||||
| HGNC | 11284 | ||||||
| 오밈 | 184753 | ||||||
| RefSeq | NM_001047 | ||||||
| 유니프로트 | P18405 | ||||||
| 기타자료 | |||||||
| EC 번호 | 1.3.1.22 | ||||||
| 로커스 | 5번 씨 p15 | ||||||
| |||||||
| 스테로이드-5α-저감효소, 알파 폴리펩타이드 2 | |||||||
|---|---|---|---|---|---|---|---|
| 식별자 | |||||||
| 기호 | SRD5A2 | ||||||
| 엔씨비유전자 | 6716 | ||||||
| HGNC | 11285 | ||||||
| 오밈 | 607306 | ||||||
| RefSeq | NM_000348 | ||||||
| 유니프로트 | P31213 | ||||||
| 기타자료 | |||||||
| EC 번호 | 1.3.1.22 | ||||||
| 로커스 | 2번 씨 p23 | ||||||
| |||||||

3-oxo-5α-스테로이드 4-수소효소라고도 알려진 5α-감소효소는 스테로이드 신진대사에 관여하는 효소다. 그들은 세 가지 대사 경로에 참여한다: 담즙산 생합성, 안드로겐과 에스트로겐 대사. SRD5A1, SRD5A2, SRD5A3 유전자로 인코딩된 5α-감소효소의 3개의 이소성분이 있다.
5α-감소화합물은 다음과 같은 일반화된 화학반응을 촉진한다.
- 3-oxo-5α-스테로이드 + 수용체 ⇌ 3-oxo-Δ-스테로이드4 + 감소 수용체
3-oxo-5α-스테로이드와 수용자가 기판인 경우, 해당 3-oxo-Δ4-스테로이드와 감소된 수용자가 제품이다. 5α-감소효소 타입 2 촉매로 일반화된 반응의 예는 다음과 같다.
- 디히드로테스토스테론 + NADP+ ⇌ 테스토스테론 + NADPH+ + H
여기서 디히드로테스토스테론은 3-oxo-5α-스테로이드, NADP는+ 수용체, 테스토스테론은 3-oxo-Δ4-스테로이드, NADPH는 감소된 수용체다.
생산 및 활동
이 효소는 남성과 여성 모두의 많은 조직, 생식기관, 고환과 난소,[1] 피부, 정맥, 전립선, 전염병 그리고 신경계를 [2]포함한 많은 기관에서 생성된다.[3][4] 5α-저감효소에는 스테로이드 5α-저감효소 1, 2, 3(SRD5A1, SRD5A2, SRD5A3)의 세 가지 등소성분이 있다.[2][5][6][7]
5α-감소효과는 3-oxo(3-케토)에 작용하며, Δ C19/C21 스테로이드제를 기판으로 삼으며4,5, "3-케토"는 세 번째 탄소와 산소의 이중 결합을 말한다. 카본스 4와 5도 'Δ4,5'로 대표되는 이중 결합을 가지고 있다. 이 반응은 공동 인자로서 NADPH의 도움을 받아 Δ의4,5 입체적이고 영구적인 분리를 포함한다. 5번째 탄소의 α 면에는 수화 음이온(H-)이, 탄소 4의 β 면에는 양성자가 배치된다.[8]
연령별 분포
5α-R1은 태아 두피와 등허리의 비엔에니탈피부에 표현되며, 성인과 유사한 태아 보형물로 표현된다. 5α-R1은 주로 상피와 5α-R2 태아 전립선의 스트롬에 표현된다. 과학자들은 태아 간, 부신, 고환, 난소, 뇌, 두피, 가슴, 생식기 피부에서 면역 결핍증을 이용해 5α-R2 발현을 찾았고 생식기 피부에서만 발견할 수 있었다.[8]
출생 후, 5α-R1은 간, 피부, 두피, 전립선을 포함한 더 많은 장소에서 표현된다. 5α-R2는 전립선, 정맥주사, 표피외, 간, 그리고 두피와 피부로 표현된다. 5α-R1과 2의 간 표현은 모두 즉각적이지만 18개월이 되면 피부와 두피에서 사라진다. 그리고 나서 사춘기에 피부와 두피에 5α-R1만 다시 발현된다.
남성 태아에서는 전립선에 5α-R1과 5α-R2가 표현되는 것으로 나타나며, 산후 수명은 물론 간, 생식기, 전립선, 비뇨기피부, 정낭, 고환, 난소, 자궁, 신장, 외분비 췌장, 뇌에도 5α-R1과 5α-R1과 5α-R2도 표현된다.[3][8]
성인기에는 5α-R1-3이[clarification needed] 보편적으로 표현된다.
기판
특정 기질로는 테스토스테론, 프로게스테론, 안드로스테리온,[9] 에피스토스테론, 코르티솔, 알도스테론, 디옥시코르티코스테론 등이 있다. 디하이드로테스토스테론 외에서는 5α 감소 스테로이드의 생리학적 역할의 상당 부분이 알려져 있지 않다.[8] 테스토스테론을 디하이드로테스토스테론으로 줄이는 것을 넘어 5알파-감소효소 이소포름 I와 II는 프로게스테론을 디히드로게스테론(DHP)으로, 디옥시코르티코스테론은 디하이드로데옥시코르티코스테론(DHDOC)으로 감소시킨다. 시험관내 및 동물 모델은 DHT, DHP 및 DHDOC의 후속 3알파 감소를 제안하며, GABAergic 억제를 강화하여 뇌 기능에 미치는 영향을 스테로이드 대사물로 유도한다. 이러한 신경 활성 스테로이드 유도체는 GABA(A) 수용체에서 알로스테릭 변조를 통해 GABA를 향상시키고 항경련제, 항우울제, 항불안제 효과를 가지며 성 및 알코올 관련 행동을 변화시킨다.[10] 5α-디하이드로코르티솔은 눈의 수성 유머에 존재하며 렌즈에 합성되며 수성 유머 자체를 만드는 데 도움이 될 수 있다.[11] 알롭 메네탄올론과 THDOC는 신경성 스테로이드로, 후자는 발작에 대한 동물의 민감성에 영향을 미친다. 사회적으로 고립된 생쥐에서 5α-R1은 피질 및 해마 영역에서 편도체에 수렴하는 글루타마테라믹 피라미드 뉴런에서 특별히 하향 조절된다. 이러한 하향 조정은 불안, 공격성, 인지장애와 같은 행동장애의 출현을 설명할 수 있다.[3][4] 5α-디하이드로알도스테론은 알도스테론과는 다르지만 강력한 항정신병 약물이다. 신장 내 그것의 형성은 식이소금의 제한에 의해 강화되며, 다음과 같이 나트륨을 유지하는 데 도움이 될 수 있음을 암시한다.[12]
- 이것은 테스토스테론에 5α-저감효소를 DHT(디히드로테스토스테론)로 변환하는 메커니즘이다. 5α-감소효소는 NADPH의 감소력을 이용하여 링의 이중 탄소 결합에 하이드라이드 시프트를 수행함으로써 에놀레이트 형성과 이후 DHT를 형성하는 튜타머화를 유발한다.[13]
- {5\alpha -substrate}+NADP+}}}">

5α-DHP는 정상적인 사이클링과 임산부가 순환하는 주요 호르몬이다.[14]
테스토스테론
5α-감소효소는 남성 성호르몬인 테스토스테론을 보다 강력한 디하이드로테스토스테론으로 바꾸는 것으로 가장 잘 알려져 있다.
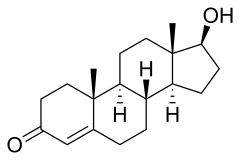

주요 차이점은 A(맨 왼쪽) 링 위의 Δ4,5 더블본드다. 도표 간의 다른 차이점은 구조와 무관하다.
전환 목록
다음과 같은 반응은 5α-저감효소에 의해 촉매되는 것으로 알려져 있다.[9]
- 콜레스테논 → 5α-콜레스타논
- 프로게스테론 → 5α-디히드로게스테론
- 3α-디히드롭로게스테론 → 알로프 메탄올론
- 3β-디히드롭로게스테론 → 이솝 메탄올론
- 디옥시코르티코스테론 → 5α-디하이드로데옥시코르티코스테론
- 코르티코스테론 → 5α-디하이드로코르티코스테론
- 코티솔 → 5α-디하이드로코르티솔
- 알도스테론 → 5α-디하이드로알도스테론
- 안드로스테네디온 → 5α-안드로스타네디온
- 테스토스테론 → 5α-디하이드로테스토스테론
- 난드롤론 → 5α-디히드로난드로논
구조

5α-감소효소는 스테로이드 기판 내 이중 결합의 NADPH 의존적 감소를 촉진해 효소를 증가시키는 막 결합 효소다.[15] 5α-감소효소 1과 2의 호몰로그램의 결정구조는 프로테오박테리아(단백질 5α-감소효소)에서 발견되었다. 이것은 공동 인자 NADPH와 스테로이드 기질 결합 주머니를 점유하는 모노레인을 수용하는 소수성 포켓을 수용하는 7개의 알파 헥스 투과 브레인 구조를 가진 모노머로 존재한다.[16] 곤충 세포에서 모놀레인은 발견되지 않고 다른 안드로겐과 억제제를 위해 하위 처리된다.[17] 일체형 7개 투과상 위상은 종에 걸쳐 보존될 가능성이 높으며, 종말의 N 종단부는 소포체 망막 루멘과 C 종단부가 시토솔을 향한다. 세포질 영역의 높은 순응 역학 관계는 NADPH/NADP+ 교환을 규제할 가능성이 있다.[17] 알려진 결정 구조에 걸친 염기서열 보존은 효소 구조에서 높은 보존을 입증했다.[16]
억제
5α 환원효소 억제 메커니즘은 복잡하지만 NADPH를 효소에 결합한 후 기질을 수반한다. 5α-감소효소 억제제는 양성 전립선 비대증, 전립선암, 패턴 탈모(안드로젠틱 탈모증), 트랜스젠더 여성을 위한 호르몬 대체요법 등에 쓰인다.
효소의 억제는 되돌릴 수 없는 스테로이드와 비스테로이드의 두 가지 범주로 분류할 수 있다. 스테로이드 억제제가 더 많으며, 예를 들어 피나스테라이드(MK-906), 두타스테라이드(GG745), 4-MA, 투로스테라이드, MK-386, MK-434, MK-963 등이 있다. 연구진은 스테로이드의 원치 않는 부작용 때문에 5α-감소효소를 억제하기 위해 비스테로이드 합성을 추진해왔다. 5α-R1의 가장 강력하고 선택적인 억제제는 이 등급에서 발견되며, 벤조퀴놀론, 비스테로이드 아릴산, 부타노산 유도체, 그리고 보다 인지적으로 다불포화 지방산(특히 리놀렌산), 아연, 녹차 등이 포함된다.[8] 리보플라빈은 5α-감소효소 억제제로도 확인되었다.[18]
또한 알파트라디올은 링지 버섯의 가노데르산, 쏘팔메토뿐만 아니라 이러한 활동 메커니즘(5α-감소효소)을 통해 작용한다고 주장되어 왔다.
5α-감소효소가 억제되면 테스토스테론의 DHT로의 전환이 감소하여 테스토스테론과 에스트라디올이 증가하게 된다. 다른 효소들은 특히 환원성 17β-히드록시스테로이드 탈수소효소, 산화성 3α-히드록시스테로이드 탈수소효소, 3β-히드록시스테로이드 탈수소효소 피부의 국소적 표현으로 전환에 대한 정도를 보상한다.[19]
자네마스티아, 발기부전, 인지기능 저하, 피로, 저혈당, 간기능 저하, 변비, 우울증 등은 5α-감소효소 억제라는 가능한 부작용 중 몇 가지에 불과하다. 약물 중단 이후에도 지속된 장기 부작용이 보고됐다.[20]
피나스테라이드
피나스테라이드는 2개의 5α-감소효소 이소엔자임(II, III)을 억제하고, 두타스테라이드는 3개 모두를 억제한다.[2] 피나스테라이드는 69nM의 평균 억제 농도 IC에서50 5α-R2를 강력하게 억제하지만, 360nM의 IC까지50 5α-R1로 효과가 낮다.[21] 피나스테라이드는 6개월 후 DHT의 평균 혈청 수치를 71% 감소시키고,[22] 시험관내에서는 감염된 세포 라인의 5α-R2와 유사한 효력에서 5α-R3을 억제하는 것으로 나타났다.[2]
두타스테라이드
두타스테라이드는 피나스테라이드보다 5α-감소효소 이소엔자임 1형과 2형을 더 잘 억제해 24주(94.7% 대 70.8%)에 DHT가 더 완전하게 감소한다.[23] 또한 전립선암에 걸린 남성의 정전기적 DHT를 3개월에 걸쳐 하루 5mg/day로 97% 감소시킨다.[24] 4개월 동안 하루 3.5mg을 투여한 두 번째 연구는 정전기 내 DHT를 99%[25] 더 감소시켰다. 생체내 DHT 억제와 두타스테라이드가[26] 5α-R3을 억제한다는 보고서는 두타스테라이드가 3중 5α 환원효소 억제제일 수 있음을 시사한다.[8]

선천성결핍증
5α-감소효소 1
5α-감소효소 1형 불활성화 수컷 생쥐는 뼈의 질량과 전림근육 그립 강도를 감소시켰는데, 이는 뼈와 근육의 5α-감소효소 1형 발현이 부족하기 때문이라고 제안되어 왔다.[29] 알파 환원효소 2형 결핍 남성 5명에서는 1형 아이소엔자임이 사춘기에 그들의 처녀성을 책임지는 것으로 생각된다.[6]
5α-감소효소 2
5α-감소효소 2 활성도가 손상된 경우 SRD5A2 유전자의 돌연변이로 인해 발생할 수 있다. 5α-감소효소 2 결핍증이라고 알려진 이 질환은 남성의 외부 성기의 비정상적인 외관으로서 나타나는 범위가 있다.
5α-감소효소 3
작은 간섭 RNA를 사용하여 세포 라인의 5α-R3 이소자임 발현을 쓰러뜨리면 세포 성장, 생존성, DHT/T 비율의 감소가 있다.[30] 테스토스테론, 안드로스테르네디온을 줄이는 능력, 안드로겐 내 프로게스테론이 아데노바이러스 벡터에 의해 전립선 세포라인을 자극하는 능력도 보였다.[8]
유전자 SRD53A에서 선천성 5α-R3 결핍은 중증 지적장애와 소뇌 및 안구 결함을 가지고 태어난 희귀한 자가 열성 질환과 연관되어 있다. 추정결손은 폴리프레놀의 단자 결합을 돌리콜에 감소시키는 것으로, 단백질의 N글리코실화에서 중요한 단계로서, 내소성 망막의 초기 단백질에 아스파라긴 잔류물을 적절히 접는 데 중요하다.[31]
신경계
정서 장애
격리 사육은 랫드 모델의 피질 및 아구질 뇌 영역에서 5α-감소효소 등소성분 1과 2의 단백질 발현을 낮추는 것으로 나타났다. 그러나 5α 감소된 대사물의 양은 영향을 받지 않았다. 이는 격리양육이 뇌에서 5α-감소효소의 발현과 활성의 변화로 이어져 도파민 신경전달의 조절이 잘못되어 초기 만성 스트레스[32] 5α-감소효소 억제제인 피나스테라이드를 이용한 치료는 SSRI가 성기능장애를 일으키는 효과를 모방한 것으로 나타났음을 의미한다.[33] 5α-감소효소는 신경스테로이드 합성에서 속도제한 효소로 특히 프로게스테론에서 allopefulanoone으로 변환하는 과정에서 [34]낮은 수준의 allope fegenaloone이 우울증, 불안, 정신분열증에 묶여 있다는 연구결과가 나왔다. 수면부족은 전전두피질에서 5α-감소효소 발현과 활성을 강화시켜 쥐에게 조증 관련 증상으로 이어질 수 있다.[34] 또한 5α-감소효소 억제제를 사용하는 것이 양성 전립선 과대증용으로 사용하는 환자 모집단의 자살이념 및 우울증과 관련이 있는지 여부도 논쟁의 대상이 된다.[35][36] 이러한 증상은 억제제의 적극적인 사용과 즉각적인 후속 조치에서 발견되었다.[35] 그러나 이러한 증상이 양성 전립선 비대증에서 자연적으로 발생하는지는 알 수 없다.[36]
시상하부-피티하수체-아드레날린 축 기능 장애
코티솔 조절의 대안적 메커니즘은 코티솔의 A-링 감소를 촉진하여 화합물을 대사시키는 5α-감소효소를 통해 조절된다.[37] 5α-저감효소 1형과 2형은 간을 통한 코티솔 간극에 관여하는 주요 효소다.[38] 과도한 코티솔은 비알코올성 지방간질환(NAFLD)에 묶여 있었지만, 체내 연구 결과 5α-감소효소 타입 2의 과다 발현이 지방생식을 억제할 수 있는 것으로 나타났다.[39] 코티솔 파괴와 지방증강에서 5α-감소효소의 주요 역할은 5α-감소효소 억제제의 부작용을 일부 해명하였다. 인간 자원 봉사자에 대한 무작위 연구에서 두타스테라이드와 피나스테라이드를 사용함으로써 5α-감소효소 억제가 남성에게 간지질 축적을 유발할 수 있다는 것이 밝혀졌다.[40] 중병에서 스트레스 반응의 일부로서 코티솔을 과하게 자극하면 간을 통한 코티솔 간 간극이 감소하고 11β-히드록시스테로이드 탈수소효소 타입 2를 통해 신장 간을 통한 코티솔의 간극이 감소할 [41]수 있으며, 코티솔의 장기 상승은 쿠싱 증후군으로 이어질 수 있다.
명명법
이 효소는 다른 수용체와 함께 CH-CH 기증자 그룹에 작용하는 산화효소군에 속한다. 이 효소 등급의 체계적 명칭은 3-oxo-5α-스테로이드:수용체 Δ-oxidoreductase이다4. 일반적으로 사용되는 다른 이름에는 다음이 포함된다.
- 5α-감소효소
- 3-옥소스테로이드 Δ-탈수소효소4
- 3-옥소-5α-스테로이드 Δ-제수소효소4
- 스테로이드 Δ-5α-감소효소4
- Δ-케토4-3 스테로이드 5α-감소효소
- Δ-옥소4-3 스테로이드 환원효소
- Δ-케토스테로이드-5α-oxidoreductase4-3
- Δ-옥소스테로이드-5α-감소효소4-3
- 3-케토-Δ-스테로이드-5α-저감효소4
- 테스토스테론 5α-감소효소
- 4-Ene-3-케토스테로이드-5α-oxidoreductase
- Δ-5α-탈수소효소4
- 3-옥소-5α-스테로이드:(수용자) Δ-oxidoreductase4
참고 항목
참조
- ^ Pinna G, Agis-Balboa RC, Pibiri F, Nelson M, Guidotti A, Costa E (October 2008). "Neurosteroid biosynthesis regulates sexually dimorphic fear and aggressive behavior in mice". Neurochemical Research. 33 (10): 1990–2007. doi:10.1007/s11064-008-9718-5. PMID 18473173. S2CID 19338424.
- ^ a b c d Yamana K, Labrie F, Luu-The V (August 2010). "Human type 3 5α-reductase is expressed in peripheral tissues at higher levels than types 1 and 2 and its activity is potently inhibited by finasteride and dutasteride". Hormone Molecular Biology and Clinical Investigation. 2 (3): 293–9. doi:10.1515/hmbci.2010.035. PMID 25961201. S2CID 28841145.
- ^ a b c Agís-Balboa RC, Pinna G, Zhubi A, Maloku E, Veldic M, Costa E, Guidotti A (September 2006). "Characterization of brain neurons that express enzymes mediating neurosteroid biosynthesis". Proceedings of the National Academy of Sciences of the United States of America. 103 (39): 14602–7. Bibcode:2006PNAS..10314602A. doi:10.1073/pnas.0606544103. PMC 1600006. PMID 16984997.
- ^ a b Agís-Balboa RC, Pinna G, Pibiri F, Kadriu B, Costa E, Guidotti A (November 2007). "Down-regulation of neurosteroid biosynthesis in corticolimbic circuits mediates social isolation-induced behavior in mice". Proceedings of the National Academy of Sciences of the United States of America. 104 (47): 18736–41. Bibcode:2007PNAS..10418736A. doi:10.1073/pnas.0709419104. PMC 2141846. PMID 18003893.
- ^ Killian J, Pratis K, Clifton RJ, Stanton PG, Robertson DM, O'Donnell L (May 2003). "5alpha-reductase isoenzymes 1 and 2 in the rat testis during postnatal development". Biology of Reproduction. 68 (5): 1711–8. doi:10.1095/biolreprod.102.009142. PMID 12606426.
- ^ a b Thiele S, Hoppe U, Holterhus PM, Hiort O (June 2005). "Isoenzyme type 1 of 5alpha-reductase is abundantly transcribed in normal human genital skin fibroblasts and may play an important role in masculinization of 5alpha-reductase type 2 deficient males". European Journal of Endocrinology. 152 (6): 875–80. doi:10.1530/eje.1.01927. PMID 15941927.
- ^ Godoy A, Kawinski E, Li Y, Oka D, Alexiev B, Azzouni F, et al. (July 2011). "5α-reductase type 3 expression in human benign and malignant tissues: a comparative analysis during prostate cancer progression". The Prostate. 71 (10): 1033–46. doi:10.1002/pros.21318. PMC 4295561. PMID 21557268.
- ^ a b c d e f g Azzouni F, Godoy A, Li Y, Mohler J (2012). "The 5 alpha-reductase isozyme family: a review of basic biology and their role in human diseases". Advances in Urology. 2012: 530121. doi:10.1155/2012/530121. PMC 3253436. PMID 22235201.
- ^ a b Paba S, Frau R, Godar SC, Devoto P, Marrosu F, Bortolato M (2011). "Steroid 5α-reductase as a novel therapeutic target for schizophrenia and other neuropsychiatric disorders". Current Pharmaceutical Design. 17 (2): 151–67. doi:10.2174/138161211795049589. PMID 21361868.
- ^ Finn DA, Beadles-Bohling AS, Beckley EH, Ford MM, Gililland KR, Gorin-Meyer RE, Wiren KM (2006). "A new look at the 5alpha-reductase inhibitor finasteride". CNS Drug Reviews. 12 (1): 53–76. doi:10.1111/j.1527-3458.2006.00053.x. PMC 6741762. PMID 16834758.
- ^ Weinstein BI, Kandalaft N, Ritch R, Camras CB, Morris DJ, Latif SA, et al. (June 1991). "5 alpha-dihydrocortisol in human aqueous humor and metabolism of cortisol by human lenses in vitro". Investigative Ophthalmology & Visual Science. 32 (7): 2130–5. PMID 2055703.
- ^ Kenyon CJ, Brem AS, McDermott MJ, Deconti GA, Latif SA, Morris DJ (May 1983). "Antinatriuretic and kaliuretic activities of the reduced derivatives of aldosterone". Endocrinology. 112 (5): 1852–6. doi:10.1210/endo-112-5-1852. PMID 6403339.
- ^ Ahmed S, Denison S (September 1998). "Mechanism based representation of the active site of 5 alpha-reductase (5AR)". Bioorganic & Medicinal Chemistry Letters. 8 (18): 2615–70. doi:10.1016/S0960-894X(98)00463-6. PMID 9873591.
- ^ Milewich L, Gomez-Sanchez C, Crowley G, Porter JC, Madden JD, MacDonald PC (October 1977). "Progesterone and 5alpha-pregnane-3,20-dione in peripheral blood of normal young women: Daily measurements throughout the menstrual cycle". The Journal of Clinical Endocrinology and Metabolism. 45 (4): 617–22. doi:10.1210/jcem-45-4-617. PMID 914969.
- ^ Wilson JD (8 February 2002). "The role of 5alpha-reduction in steroid hormone physiology". Reproduction, Fertility, and Development. 13 (7–8): 673–8. doi:10.1071/rd01074. PMID 11999320.
- ^ a b Han Y, Zhuang Q, Sun B, Lv W, Wang S, Xiao Q, et al. (January 2021). "Crystal structure of steroid reductase SRD5A reveals conserved steroid reduction mechanism". Nature Communications. 12 (1): 449. Bibcode:2021NatCo..12..449H. doi:10.1038/s41467-020-20675-2. PMC 7815742. PMID 33469028.
- ^ a b Xiao Q, Wang L, Supekar S, Shen T, Liu H, Ye F, et al. (October 2020). "Structure of human steroid 5α-reductase 2 with the anti-androgen drug finasteride". Nature Communications. 11 (1): 5430. Bibcode:2020NatCo..11.5430X. doi:10.1038/s41467-020-19249-z. PMC 7591894. PMID 33110062.
- ^ Nakayama O, Yagi M, Kiyoto S, Okuhara M, Kohsaka M (December 1990).