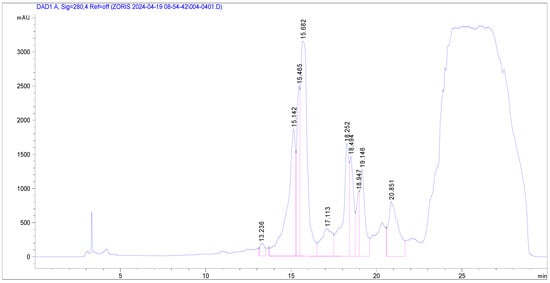
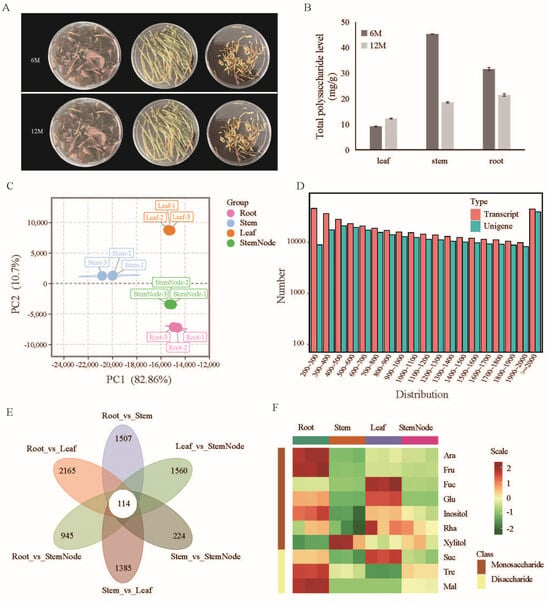
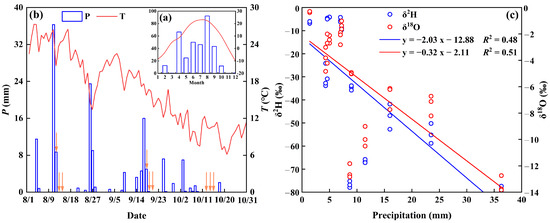
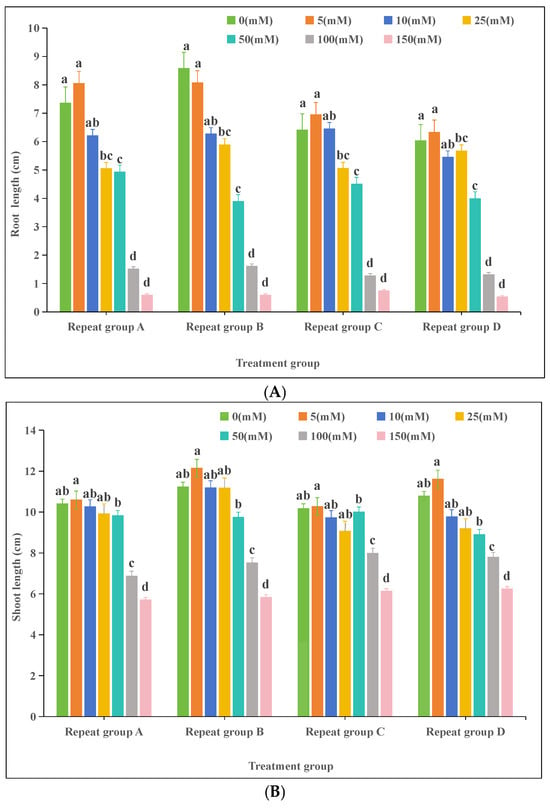
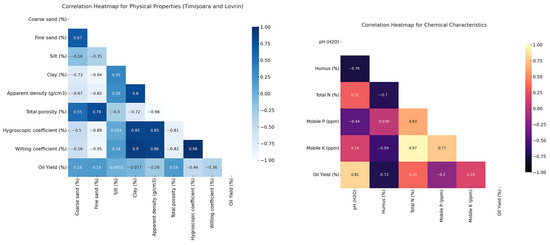
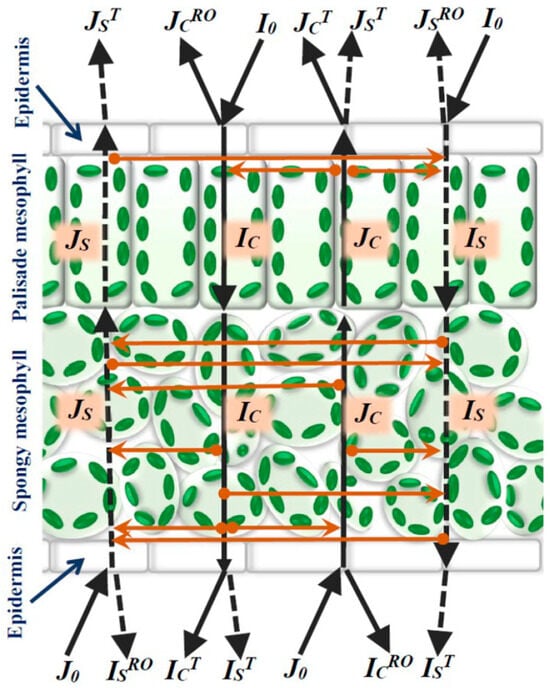
Plants 2024, 13(23), 3273; https://doi.org/10.3390/plants13233273 - 21 Nov 2024
Abstract
This long-term field study conducted in Yancheng, China, evaluated the effects of diverse crop rotation sequences on rice growth, yield, and soil properties. Six rotation treatments were implemented from 2016 to 2023 as follows: rice–wheat (control), rice––rape, rice–hairy vetch, rice–barley, rice–faba bean, and
[...] Read more.
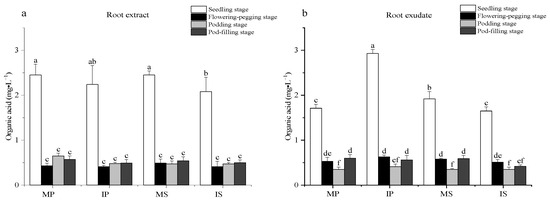
This long-term field study conducted in Yancheng, China, evaluated the effects of diverse crop rotation sequences on rice growth, yield, and soil properties. Six rotation treatments were implemented from 2016 to 2023 as follows: rice–wheat (control), rice––rape, rice–hairy vetch, rice–barley, rice–faba bean, and rice–winter fallow. Rice growth parameters, yield components, biomass accumulation, and soil properties were measured. Results showed that legume-based rotations, particularly rice–faba bean and rice–hairy vetch, significantly improved rice growth and yield compared to the rice–wheat control. The rice–faba bean rotation increased yield by 19.1% to 8.73 t/ha compared to 7.33 t/ha for the control, while rice–hairy vetch increased yield by 11.9% to 8.20 t/ha. These rotations also demonstrated higher biomass production efficiency, with increases of 33.33% and 25.00%, respectively, in spring crop biomass. Soil nutrients improvements were observed, particularly in available nitrogen, potassium, and electrical conductivity. Legume-based rotations increased the available nitrogen by up to 35.9% compared to the control. The study highlights the potential of diversified crop rotations, especially those incorporating legumes, to enhance rice productivity and soil health in subtropical regions. These findings have important implications for developing sustainable and resilient rice-based cropping systems to address challenges of food security and environmental sustainability in the face of climate change and resource constraints.
Full article
(This article belongs to the Special Issue Effects of Conservation Tillage on Crop Cultivation and Production)