Fotosyntes
|
|
Den här artikeln behöver fler eller bättre källhänvisningar för att kunna verifieras. (2014-04) Åtgärda genom att lägga till pålitliga källor (gärna som fotnoter). Uppgifter utan källhänvisning kan ifrågasättas och tas bort utan att det behöver diskuteras på diskussionssidan. |
| Ämnesomsättningen | |
| Denna artikel är en del i serien Ämnesomsättningen med följande delar:
| |
| Metabolism · Katabolism · Anabolism | |
| Katabolism | |
|---|---|
| Matspjälkning · Glykolys · Beta-oxidation · Trans-/Deaminering · Citronsyracykeln · Elektrontransportkedjan · Oxidativ fosforylering · Ureacykeln | |
| Anabolism | |
| Glukoneogenes · Proteinsyntes · Fettsyrasyntes | |
| Se även | |
| Fotosyntes · Cellandning · Malat-aspartatskytteln |
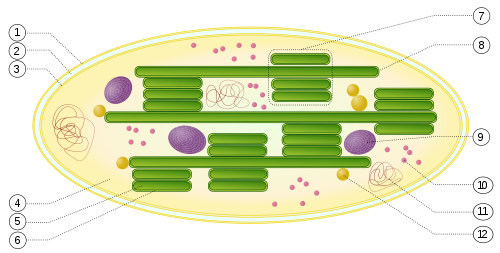

Fotosyntes är den process där växter och andra levande organismer tar hand om energi från solljus och lagrar energin i kemiska bindningar.
Den vanligaste formen är kolsyreassimilation hos växter och cyanobakterier, som innebär att de under dagen tar in koldioxid, vatten och solenergi som de med hjälp av klorofyll omvandlar till syre och druvsocker. Syret och druvsockret använder de vid cellandningen och under natten när de avger koldioxid.
Fotosyntesen i växter försiggår i bladen vars celler har kloroplaster, som anses vara symbiotiska cyanobakterier. Ljusreaktionerna sker i anslutning till tylakoidernas fosfolipidmembraner, medan mörkerreaktionen äger rum i det vätskefyllda stromat.
Fotosyntes utan syreproduktion anses vara en mer primitiv process och förekommer fortfarande hos bakterier[1].
Kemiska reaktioner
[redigera | redigera wikitext]
Kemisk formel för koldioxidfixerande fotosyntes[2]:
- 6 H2O + 6 CO2 + ljusenergi → C6H12O6 (druvsocker) + 6 O2
Vissa bakterier, cyanobakterier, utnyttjar vatten vid fotosyntes, på samma sätt som växter. Andra fotoautotrofa bakterier klarar inte av detta, utan använder istället H2S (svavelväte), som är en mer lättoxiderad förening[1].
Beskrivningen av den kemiska reaktionen är starkt förenklad jämfört med den faktiska processen som äger rum i kloroplasterna. Koldioxiden binds i själva verket genom en lång process med en rad katalysatorer och kemiska reaktioner i flera olika steg i mellanleden. Denna reaktionssekvens kallas Calvin-cykeln.[3]
Antennpigment
[redigera | redigera wikitext]Antennpigment är de pigmentmolekyler som fångar in solljuset och transporterar vidare energin till klorofyll i närliggande reaktionscentra där fotosyntesen sker. De deltar inte i själva syntesen. Exempel på molekyler som kan fungera som antennpigment hos växter är klorofyll a, klorofyll b och karotener. Rödalger och cyanobakterier har fykobiliproteiner, och brunalger en karotenoid vid namn fukoxantol.
Klorofyll
[redigera | redigera wikitext]
Fotosyntesen sker i särskilda strukturer, kloroplaster hos eukaryoter, som finns i växtcellerna. Kloroplasterna innesluts i ett dubbelt membran. Detta är ett av indicierna för att kloroplasterna en gång i tiden kan ha varit självständiga organismer. Prokaryoter har inte kloroplast.
Den kemiska reaktionen som binder solljuset sker genom molekyler av klorofyll eller karotenoider (som är gula till färgen) som sitter på ytan av små membransäckar, tylakoider, inuti kloroplasterna. Hos gröna växter finns klorofyll i två varianter, typ a och b, som absorberar något olika våglängder av ljus och har lite olika uppgifter i den kemiska processen. Hos alger och bakterier förekommer ytterligare varianter av klorofyll.
Klorofyll är det ämne som ger växter deras gröna färg. Ämnet spelar en avgörande roll i fotosyntesen, det vill säga växternas omvandling av koldioxid, vatten och energi till kolhydrater och syrgas.
Klorofyllmolekylerna deltar inte i själva reaktionen utan fungerar som en sorts ljussamlande antenn. De tar upp det inkommande ljuset och överför energin till reaktionscentret.
Klorofyll är en komplex molekyl som är uppbyggd kring en ring av organiska ämnen med en magnesiumatom i mitten och en sidokedja som består av en alkoholmolekyl (fytol hos klorofyll typ a och b). Grundstrukturen har vissa likheter med hemoglobin i blodet hos djur, men där finns en järnatom i mitten istället för magnesium.
Klorofyll finns i flera olika varianter. Typ a och b finns hos gröna växter. Typ c och d finns hos alger, medan bakterier som använder fotosyntes har många olika varianter av klorofyll.
Klorofyll typ a och b absorberar olika våglängder av solljus. Klorofyll typ c och d absorberar blått ljus (400-500 nm) och rött ljus (600-700 nm). De skiljer sig också något i sin funktion i den kemiska reaktionen som omvandlar vatten, koldioxid och solljus till socker och syre, 6H2O + 6CO2 + solljus → C6H12O6 + 6O2. Detta kan till exempel förklaras genom trädens löv som absorberar rött ljus på våren och sommaren, så att vi ser bladen som gröna. På hösten absorberar löven mest grönt ljus och reflekterar komplementfärgerna (i detta fall rött och gult); därför blir löven gula/orangea. I slutet av 1800-talet visade det sig att rött och blått ljus är effektiva vid fotosyntesen.
Ämnet är godkänt som färgämne i livsmedel; det har då E-nummer E 140.
Arkéer fotosyntiserar med proteinet bakteriorodopsin istället för klorofyll[4].
Fotosyntesens effektivitet beroende på våglängd.
[redigera | redigera wikitext]Vid 660 nm (rött) finns ett maximum som ges relativvärdet 100. Vid 550 nm (grön) finns ett minimum som får värdet 40. Vid 440 nm (blå) finns ett annat maximum som får relativvärdet 80. Vid 400 och 700 nm är relativvärdet ca 35.[5]
Energi
[redigera | redigera wikitext]Cyanobakterier med klorofyll f kan använda infraröd strålning[6], men växter utnyttjar enbart det synliga spektrumet, 400-700 nm, i huvudsak blått (400–500 nm) och rött (600–700 nm) ljus, vilket motsvarar 40-45 % av instrålad solenergi. På grund av andra omständigheter – tillgång på växttillgängligt vatten och växtnäring, växternas egen andning, växtperiod – lagras ungefär 1 % av infallande ljus kemiskt i växter. Under gynnsamma förhållanden kan det uppgå till 4 %. Sett över hela jorden är genomsnittet 0,1 %. Energiinnehållet i växter är vanligen 4,5–5 kWh eller 16–18 MJ per kg torr massa. I trä kan det bli upp till 5,6 kWh eller drygt 20 MJ per kg torr ved.
C3- och C4-växter
[redigera | redigera wikitext]Det finns två olika typer av fotosyntetiska reaktioner hos växter, där skillnaden är hur mellanstegen i koldioxidfixeringen sker i växtens celler. Växter med dessa olika reaktioner kan därför delas in i C3- och C4-växter. Dessutom förekommer en variant av C4 som kallas CAM.
C3 är en enklare process, som förekommer i träd. C4 sker i två steg och förekommer i gräs. De olika processerna binder olika mycket av kolets tunga isotop kol-13, vilket gör det möjligt att utifrån isotopsammansättningen hos arkeologiska fynd av tandemalj spåra huruvida ett djur har levt av trädens löv och frukter eller av gräs, samt om ett rovdjur har livnärt sig på lövätare (som giraff) eller gräsätare (som antilop). En liknande skillnad i sammansättningen av stabila isotoper gör det möjligt att skilja landbaserade växter och djur från havslevande.
Begreppets historia och avgränsning
[redigera | redigera wikitext]År 1771 upptäckte Joseph Priestley att möss dog om de var ensamma i lufttäta behållare, men överlevde om de var där tillsammans med mynta. Han drog först något felaktiga slutsatser av detta, men anses tillsammans med Jan Ingenhousz – som beskrev ljusets betydelse – ha upptäckt fotosyntesen.
Nordisk familjebok från början av 1900-talet definierar fotosyntes som ”kolsyreassimilationen under ljusets medverkan. Se Assimilation” Under uppslagsordet Assimilation kan man vidare läsa: ”I växtfysiologien förstås med assimilation sammanfattningen af alla de processer, hvarigenom de af växten upptagna näringsämnena ombildas till i organismen ingående beståndsdelar. Den ojämförligt mest framträdande assimilationsprocessen är kolsyrans; förr i tiden förstod man också med assimilation endast kolsyreassimilationen. Kolsyreassimilationen är en process, hvarigenom organisk substans bildas af kolsyra och vatten, och för densammas förlopp är energitillförsel nödvändig. Hos gröna växter är ljuset energikällan”
”Det närmare förloppet vid kolsyreassimilationen är ännu ej känt. Man antager, att kolsyran förekommer vid assimilationen som hydrat; och en del forskare anse, att den första produkten härvid skulle vara formaldehyd, som faktiskt kan påvisas i bladen. Så mycket kan sägas, att den första produkten af kolsyreassimilationen är en ökning af det organiskt bundna kolet i klorofyllkropparna. Häraf uppstå sedan alltefter växtens art olika organiska ämnen, och i vanliga fall är assimilationsprodukten stärkelse”
Tidigt stod alltså fotosyntes för all den uppbyggnad av organisk substans som pågick i (framför allt) växterna. Sedan forskarna kartlagt fotosyntesen mer i detalj har det blivit rimligt att sätta en gräns mellan det som görs direkt med hjälp av ljuset och processer som snarare bör ses som att organismen bygger om de molekyler som fotosyntesen skapat. Men det går inte att från objektiva kriterier säga exakt var gränsen går mellan fotosyntesen och andra kemiska reaktioner i den levande organismen.
Processens delsteg i huvuddrag
[redigera | redigera wikitext]Ljusreaktionerna
[redigera | redigera wikitext]Följande kedja av tio delsteg brukar tillsammans kallas ljusreaktionerna. Resultatet av dem är att energi lagrats i adenosintrifosfat (ATP) och reducerat nikotinamid-adenin-dinukleotidfosfat (NADPH).
- En foton kolliderar med en elektron i en klorofyllmolekyl och flyttar elektronen till en bana som har högre potentiell energi och i genomsnitt befinner sig längre bort från närmaste atomkärna. Man säger att elektronen blir exciterad.
- En exciterad elektron i klorofyll exciterar en elektron i ett närliggande klorofyll. Detta överför excitationsenergin till den andra elektronen, så att den första elektronen återgår till sitt normaltillstånd. Detta sker många gånger, så att excitationsenergin flyttar omkring mellan klorofyllmolekylerna.
- En exciterad elektron lämnar två klorofyllmolekyler och övergår till en feofytin-molekyl. Detta kan ske endast i ett så kallat fotosyntetiskt reaktionscentrum. Reaktionscentret ligger inbäddat i tylakoidens membran. Detta reaktionscentrum kallas också fotosystem II.
- Ett enzym spjälkar vatten, och oxiderar därigenom syre till syremolekyler. Samtidigt reduceras reaktionscentrat på fotosystem II. Syret som bildas vid spjälkningen diffunderar ut i atmosfären.
- Feofytin-molekylen lämnar elektronen vidare till ett cytokrom-komplex. Cytokrom-komplexet ligger också inbäddat i tylakoidens membran.
- I cytokrom-komplexet går elektronen vidare till en plastokinon-molekyl och förlorar genom detta potentiell energi (den binds starkare av plastokinon än av feofytin). Denna energi använder cytokrom-komplexet till att transportera protoner genom membranet, till tylakoidens inre. Härmed uppstår en hög koncentration av protoner i tylakoiden, det vill säga att det blir surt där.
- Plastokinon-molekylen transporterar elektronen till ytterligare ett fotosystem (fotosystem I) med klorofyll och exciterar där en elektron i klorofyll.
- Den exciterade elektronen i det andra klorofyllet exciteras ytterligare av en ytterligare foton.
- Den exciterade elektronen lämnar klorofyllmolekylen och övergår till en molekyl oxiderat nikotinamid-adenin-dinukleotidfosfat (NADP+). NADP+ reduceras därmed till NADPH. I fotosyntesen är NADPH i princip en bärare av en elektron med hög energi.
- Den skillnad i protonkoncentration som beskrivs i ett av de föregående stegen används av ett proteinkomplex som släpper tillbaka några protoner och använder den energiminskning och entropiökning som detta innebär till att producera ATP genom att kombinera adenosindifosfat (ADP) med fosfat.
Mörkerreaktionerna
[redigera | redigera wikitext]- Enzymet fosforibulos-kinas fäster en fosfatgrupp på en ribulos-5-fosfat-molekyl, så att det blir ribulos-1,5-bisfosfat.
- Enzymet rubisco kombinerar ribulos-1,5-bisfosfat med koldioxid. Den molekyl som detta resulterar i faller omedelbart isär till två stycken 3-fosfoglycerat-molekyler.
- En del av 3-fosfoglycerat-molekylerna byggs via ett antal transformationer om till fruktosfosfat, som vidare kan omvandlas till någon av ett mycket stort antal olika kolhydrater. De flesta 3-fosfoglycerat-molekylerna byggs om till ribulos-5-fosfat och används åter av rubisco för att binda koldioxid. På detta sätt bildas den så kallade Calvin-cykeln.
Se även
[redigera | redigera wikitext]Referenser
[redigera | redigera wikitext]- ^ [a b] Martin, William F; Bryant, Donald A; Beatty, J Thomas. ”A physiological perspective on the origin and evolution of photosynthesis”. FEMS Microbiology Reviews 42 (2). doi:. ISSN 0168-6445. PMID 29177446. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5972617/. Läst 3 april 2021.
- ^ ”The Cell, Respiration and Photosynthesis”. www.globalchange.umich.edu. Arkiverad från originalet den 2 oktober 2015. https://web.archive.org/web/20151002190140/http://www.globalchange.umich.edu/globalchange1/current/lectures/kling/energyflow/psn_primer.html. Läst 23 september 2015.
- ^ ”Kloroplastens inre”. www-vaxten.slu.se. Arkiverad från originalet den 15 december 2016. https://web.archive.org/web/20161215085631/http://www-vaxten.slu.se/vaxten/fotosyntes/mrkerrea.htm. Läst 23 september 2015.
- ^ Hayashi, Shigehiko; Tajkhorshid, Emad; Schulten, Klaus. ”Molecular Dynamics Simulation of Bacteriorhodopsin's Photoisomerization Using Ab Initio Forces for the Excited Chromophore”. Biophysical Journal 85 (3). doi:. PMID 12944261. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1303320/. Läst 3 april 2021.
- ^ J. Krochmann und R. Rattunde. Grössen und Einheiten der für die Photosynthese wirksamen Strahlungen, Gartenbauwissenschaft, 45, S. 214-219, 1980.
- ^ Nürnberg, Dennis J.; Morton, Jennifer; Santabarbara, Stefano; Telfer, Alison; Joliot, Pierre; Antonaru, Laura A.; Ruban, Alexander V.; Cardona, Tanai; et al. (2018-06-14). ”Photochemistry beyond the red limit in chlorophyll f–containing photosystems”. Science (American Association for the Advancement of Science (AAAS)) 360 (6394): sid. 1210–1213. doi:. ISSN 0036-8075.
Externa länkar
[redigera | redigera wikitext]Wikimedia Commons har media som rör Fotosyntes.
| ||||||||||||||||||||||||
|