Neuron
Neuron, komórka nerwowa – rodzaj elektrycznie pobudliwej komórki zdolnej do przetwarzania i przewodzenia informacji w postaci sygnału elektrycznego. Neurony są podstawowym elementem układu nerwowego zwierząt. Najwięcej neuronów znajduje się w ośrodkowym układzie nerwowym, w skład którego wchodzi mózgowie oraz rdzeń kręgowy.
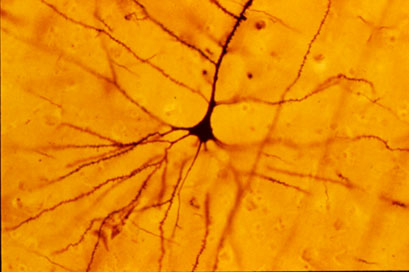


Typowy neuron jest zbudowany z ciała komórki (perikarion) oraz odchodzących od niego wypustek: aksonu i dendrytów[1]. Neurony kontaktują się między sobą poprzez synapsy, tworząc sieci neuronowe. Informacje od innych neuronów są odbierane przez synapsy położone na dendrytach, przewodzone wzdłuż neuronu i przekazywane dalej do synaps na zakończeniach aksonu.
Przewodzenie informacji w postaci sygnału elektrycznego jest możliwe dzięki temu, że wszystkie neurony są elektrycznie pobudliwe, czyli zdolne do generowania i przewodzenia potencjałów elektrycznych. Niepobudzony neuron utrzymuje potencjał spoczynkowy (będący różnicą między potencjałem elektrycznym wnętrza neuronu a zewnętrznej powierzchni błony) dzięki działaniu leżących w błonie pomp jonowych, które przenoszą określone jony przez błonę i generują różnicę w stężeniu tych jonów po obu stronach błony.
Pod wpływem dostatecznie silnego bodźca dochodzi do zmian w przepuszczalności określonych jonów przez błonę, co prowadzi do powstania potencjału czynnościowego – sygnału elektrycznego, który rozprzestrzenia się wzdłuż aksonu do synaps znajdujących się w zakończeniach aksonu.
Neurony powstają w procesie neurogenezy z neuronalnych komórek macierzystych i po zróżnicowaniu nie ulegają już dalszym podziałom komórkowym. Neurogeneza zachodzi głównie w okresie prenatalnym (przed narodzeniem), u dorosłych osobników proces ten zachodzi jedynie w określonych częściach mózgu, m.in. hipokampie i opuszce węchowej.
Budowa neuronu
edytujBudowa zewnętrzna
edytujNeuron składa się z ciała komórki (zwanego perykarionem lub somą) i odchodzących od niego wypustek nerwowych: dendrytów i aksonów. Cała komórka nerwowa, podobnie jak wszystkie inne komórki zwierzęce, pokryta jest błoną komórkową o grubości ok. 5 nm, która w przypadku komórek nerwowych bywa nazywana neurolemmą.
Dendryty
edytujDendryty są zazwyczaj silnie rozgałęzione i mogą stanowić nawet do 90% powierzchni wielu neuronów. Są pokryte tysiącami synaps, przez które odbierają informację pochodzącą z zakończeń aksonalnych innych neuronów i przekazują ją do ciała komórki. Neuron może mieć jeden lub wiele dendrytów (tworzących wspólnie tzw. drzewko dendrytyczne o wzorze typowym dla danego typu neuronów). Dendryty niektórych neuronów są pokryte kolcami dendrytycznymi, na których również znajdują się synapsy.
Aksony
edytujNeurony mają zazwyczaj tylko jeden akson wychodzący z ciała komórki w miejscu nazwanym wzgórkiem aksonu. Aksony zazwyczaj rozgałęziają się (szczególnie na końcu dystalnym, czyli tym położonym dalej od ciała komórki), tworząc kolaterale aksonu. Zakończenia aksonów, zwane kolbkami, są miejscem kontaktu z innymi neuronami (lub innym typem komórki), czyli tworzą synapsę.
Aksony mogą być otoczone osłonką mielinową – są wtedy nazywane aksonami zmielinizowanymi. Pokrycie osłonką zapewnia szybsze przewodzenie impulsu nerwowego – spełnia ona funkcję izolatora elektrycznego, a także zapewnia ochronę mechaniczną. W odstępach ok. 50 μm–1 mm w osłonce występują przerwy zwane przewężeniami Ranviera. Osłonka mielinowa jest wytwarzana przez komórki glejowe – w mózgowiu przez oligodendrocyty, a w obwodowym układzie nerwowym przez komórki Schwanna.
Rozmiary neuronów
edytujIstnieją ogromne różnice w rozmiarach różnych typów komórek nerwowych u różnych gatunków zwierząt. Ciało komórkowe typowego ludzkiego neuronu ma średnicę ok. 20 μm. Najmniejsze neurony mają średnicę ok. 5 μm, największe ok. 120 μm. Ludzkie aksony mają średnicę w granicach 0,2 do 20 μm (u bezkręgowców nawet do 1 mm) i długość od kilku μm do kilku metrów[2].
Budowa wewnętrzna
edytujCiało komórki nerwowej (perykarion) zawiera typowe organelle komórki zwierzęcej: jądro komórkowe, gładkie i szorstkie retikulum endoplazmatyczne, cytoszkielet (mikrotubule, mikrofilamenty, a także filamenty pośrednie, które w przypadku neuronów są często nazywane neurofilamentami), mitochondria, aparat Golgiego oraz rybosomy.
Cechą charakterystyczną komórek nerwowych jest występowanie ziarnistości Nissla – gęsto pokrytych rybosomami struktur tworzonych przez szorstkie retikulum endoplazmatyczne. Ich obecność odzwierciedla zdolność neuronów do utrzymywania szybkiego tempa syntezy białek, która zachodzi właśnie w rybosomach.
Skład organelli we wnętrzu dendrytów jest podobny do tego w perykarionie, natomiast we wnętrzu aksonu brak jest szorstkiego retikulum endoplazmatycznego i rybosomów, co oznacza że wszystkie znajdujące się w nim białka muszą być syntetyzowane w ciele komórki i przetransportowane wzdłuż aksonu. Transport białek z ciała komórki do zakończeń aksonów nazywany jest transportem aksonalnym. Zsyntetyzowane w ciele komórki białka są zamykane w specjalnych pęcherzykach i wędrują wzdłuż leżących w aksonie mikrotubuli dzięki białkom motorycznym zwanym kinezynami, które energię do generowania ruchu pozyskują z ATP. Ruch w przeciwną stronę – od zakończeń aksonu do ciała komórki – odbywa się przy udziale innych białek motorycznych zwanych dyneinami.
Synapsy
edytujSynapsy są miejscem komunikacji neuronu z innym neuronem lub komórką efektorową (komórką mięśniową lub gruczołową). Synapsa składa się z części presynaptycznej, którą stanowi błona komórkowa zakończenia aksonu oraz z części postsynaptycznej, którą stanowi błona komórkowa innego neuronu (zazwyczaj dendrytu, ale synapsy mogą też być tworzone na ciele komórki lub aksonie innego neuronu) lub komórki efektorowej. Między częścią presynaptyczną a postsynaptyczną znajduje się szczelina synaptyczna.
Większość synaps to synapsy chemiczne, które przekształcają sygnał elektryczny (impuls nerwowy) dochodzący do zakończenia aksonu w sygnał chemiczny poprzez wydzielenie neuroprzekaźnika z błony presynaptycznej do szczeliny synaptycznej. Neuroprzekaźnik łączy się z receptorami w błonie postsynaptycznej wywołując depolaryzację przekształcenie informacji z powrotem w sygnał elektryczny.
Znacznie rzadziej spotykane są synapsy elektryczne, w których szczelina synaptyczna jest bardzo wąska, a impuls nerwowy jest przekazywany bezpośrednio z jednego neuronu na drugi poprzez koneksony – specjalne kanały tworzące ścisłe połączenie między neuronami.
Funkcjonowanie neuronów
edytujPodstawową funkcją neuronów jest przenoszenie i przetwarzanie informacji w postaci impulsów nerwowych, będących krótkotrwałymi, gwałtownymi zmianami potencjału błony komórkowej neuronu. Impulsy nerwowe w warunkach naturalnych są przewodzone tylko w jednym kierunku (ortodromowo): od początkowego segmentu aksonu do synaps znajdujących się na jego zakończeniach.
Funkcjonalnie neuron można podzielić na cztery strefy:
- strefa wejścia – dendryty i ciało komórki, które odbierają impulsy od innych neuronów poprzez znajdujące się na nich synapsy
- strefa inicjacji – początkowy odcinek aksonu; tutaj powstaje potencjał czynnościowy neuronu
- strefa przewodzenia – akson
- strefa wyjścia – synapsy na zakończeniach aksonu
Potencjał spoczynkowy
edytujW stanie niepobudzonym potencjał elektryczny wewnątrz komórki nerwowej jest niższy niż na zewnątrz. Różnica ta jest nazywana potencjałem spoczynkowym neuronu i wynosi ok. -70 mV. Jest spowodowana nierównomiernym rozmieszczeniem naładowanych jonów po obu stronach błony. Za utrzymywanie potencjału spoczynkowego odpowiada działanie leżących w błonie komórkowej aktywnych pomp sodowo-potasowych, które wypompowują kationy sodu poza komórkę, a także odmienna przepuszczalność błony komórkowej dla różnych jonów.
Potencjał czynnościowy
edytujW wyniku pobudzenia neuronu przez odpowiednio silny bodziec (np. pobudzenie synaps na dendrytach neuronu przez inne neurony) dochodzi do powstania potencjału czynnościowego wskutek zmian w przepuszczalności błony komórkowej neuronu dla poszczególnych jonów. Następuje otwarcie się kanałów sodowych i gwałtowny napływ dodatnio naładowanych jonów sodu do wnętrza komórki. Prowadzi to do wyrównania potencjałów po obu stronach błony (0 mV), a następnie do odwrócenia się polaryzacji błony (do ok. +35 mV) – jest to tzw. faza depolaryzacji. Następnie kanały sodowe ulegają inaktywacji, otwierają się natomiast kanały potasowe, co powoduje wypływ dodatnio naładowanych jonów potasu na zewnątrz komórki i powrót do ujemnej polaryzacji błony komórkowej – jest to tzw. faza repolaryzacji. Faza depolaryzacji i repolaryzacji potencjału czynnościowego (iglica) trwa nie więcej niż 1 ms. Po nich następuje trwająca kilka milisekund hiperpolaryzacja następcza, czyli spadek potencjału błony poniżej potencjału spoczynkowego (do ok. -80 mV), a następnie powrót do wartości potencjału spoczynkowego.
Podczas trwania potencjału czynnościowego błona komórkowa neuronu jest całkowicie niepobudliwa, co oznacza że neuron nie może wytworzyć nowego potencjału czynnościowego. W czasie hiperpolaryzacji następczej pobudliwość neuronu jest silnie zmniejszona. Okresy niepobudliwości i zmniejszonej pobudliwości neuronu zwane są odpowiednio refrakcją bezwzględną i względną. Dzięki nim istnieje ograniczenie dla maksymalnej częstotliwości potencjałów czynnościowych jakie może wytwarzać neuron. Ponadto dzięki temu, że fragment błony neuronu w którym właśnie wystąpił potencjał czynnościowy jest niepobudliwy, potencjał czynnościowy może przenosić się wzdłuż aksonu tylko w jednym kierunku.
Zasada wszystko albo nic
edytujPrzewodzenie sygnałów przez neuron podlega zasadzie wszystko albo nic tzn. neuron wytwarza potencjał czynnościowy lub nie. Wszystkie powstające potencjały czynnościowe w danej komórce nerwowej mają tę samą wielkość, bez względu na wielkość bodźca (o ile tylko jest on wystarczająco intensywny, aby wywołać powstanie potencjału). Silniejsza stymulacja neuronów (bodźcami ponadprogowymi) nie prowadzi do wytwarzania silniejszych potencjałów. Może natomiast prowadzić do zwiększenia częstotliwości wytwarzania potencjałów przez neuron.
Historia badania funkcji neuronów
edytujZnaczna część pierwotnej wiedzy o aktywności elektrycznej neuronów pochodzi z eksperymentów na aksonach wielkich kałamarnic. W 1937 r. John Zachary Young zaproponował aksony kałamarnic jako model użyteczny do studiów nad elektrycznymi właściwościami neuronów. Są one dużo większe od ludzkich neuronów, więc lepiej nadawały się do eksperymentów w tamtych czasach.
Klasyfikacja neuronów
edytuj1 – jednobiegunowe
2 – dwubiegunowe
3 – wielobiegunowe
4 – pseudojednobiegunowe
Strukturalna
edytuj- Ze względu na liczbę wypustek (aksonów i dendrytów) odchodzących od ciała komórki neurony dzieli się na:
- jednobiegunowe posiadające tylko jedną wypustkę (np. w podwzgórzu)
- pseudojednobiegunowe posiadające dwie wypustki (1 akson i 1 dendryt), które uległy fuzji na początkowym odcinku (np. zwoje czuciowe nerwów czaszkowych i rdzeniowych)
- dwubiegunowe posiadające dwie wypustki – akson i dendryt (np. komórki siatkówki oka, błony węchowej)
- wielobiegunowe posiadające trzy lub więcej wypustek (1 akson, kilka dendrytów); do tego typu zalicza się zdecydowaną większość neuronów układu nerwowego kręgowców
- Ze względu na długość aksonów:
- neurony typu Golgi I (projekcyjne) o długich aksonach, przenoszące informacje na duże odległości
- neurony typu Golgi II (interneurony) o krótkich aksonach, przenoszące informacje na małe odległości
- Ze względu na morfologię w poszczególnych częściach mózgu można wyróżnić kilka charakterystycznych typów neuronów:
- neurony piramidowe leżące w korze mózgu, posiadają ciało komórkowe o trójkątnym kształcie oraz długie aksony (są to neurony typu Golgi I)
- neurony Purkiniego leżące w korze móżdżku, posiadają silnie rozgałęzione drzewko dendrytyczne, również należą do neuronów typu Golgi I
- komórki gwiaździste to niewielkie neurony leżące w korze mózgu, posiadają liczne dendryty układające się w kształt gwiazdy
- neurony wrzecionowate leżące w korze mózgu, charakteryzują się ciałem komórkowym o wrzecionowatym kształcie, są bipolarne
- neurony koszyczkowe leżące w korze mózgu i móżdżku posiadają rozwidlające się aksony, których zakończenia otaczają ciało docelowego neuronu na kształt koszyczka
Funkcjonalna
edytuj- Pod względem kierunku przekazywania sygnału neurony dzieli się na:
- czuciowe (aferentne, dośrodkowe) przenoszą informacje od receptorów w tkankach i narządach ciała do ośrodkowego układu nerwowego
- ruchowe (eferentne, odśrodkowe) przenoszą informacje od ośrodkowego układu nerwowego do komórek efektorowych – mięśniowych lub gruczołowych
- kojarzeniowe (interneurony, pośredniczące) przenoszą informacje między neuronami
- Neurony dzieli się również według głównego wydzielanego neuroprzekaźnika. Według tego kryterium wyróżnia się między innymi neurony:
- cholinergiczne – głównym neuroprzekaźnikiem jest acetylocholina
- dopaminergiczne – dopamina
- GABA-ergiczne – kwas gamma-aminomasłowy
- noradrenergiczne – noradrenalina
- serotoninergiczne – serotonina
- glutaminergiczne – glutaminian.
Liczba neuronów
edytujSzacuje się, że ludzki mózg zawiera ok. 1,5-1,6 x 1011 neuronów[3][4] i 1014 synaps[5]. Ogromna większość neuronów znajduje się w móżdżku, a najliczniejszą populacją komórek są małe neurony ziarniste móżdżku[6].
Nicień Caenorhabditis elegans, będący organizmem modelowym w biologii, posiada jedynie 302 neurony. Muszka owocowa Drosophila melanogaster posiada ok. 100 000 neuronów.
Przypisy
edytuj- ↑ podyplomie.pl – Podstawowe informacje o morfologii i fizjologii układu nerwowego [online], www.podyplomie.pl [dostęp 2018-04-26].
- ↑ Mark F. Bear, Barry W. Connors, Michael A. Paradiso: Neuroscience : exploring the brain. Philadelphia, PA: Lippincott Williams Wilkins, 2007, s. 28-46. ISBN 0-7817-6003-8.
- ↑ Alan Longstaff: Neurobiologia. Warszawa: Wydawnictwo Naukowe PWN, 2012, s. 1-26. ISBN 978-83-01-13805-9.
- ↑ Piotr Jaśkowski: Neuronauka poznawcza. Vizja Press & IT, 2009, s. 20-25. ISBN 978-83-61086-51-2.
- ↑ RW. Williams, K. Herrup. The control of neuron number.. „Annu Rev Neurosci”. 11, s. 423-53, 1988. DOI: 10.1146/annurev.ne.11.030188.002231. PMID: 3284447.
- ↑ FA. Azevedo, LR. Carvalho, LT. Grinberg, JM. Farfel i inni. Equal numbers of neuronal and nonneuronal cells make the human brain an isometrically scaled-up primate brain.. „J Comp Neurol”. 513 (5), s. 532-41, Apr 2009. DOI: 10.1002/cne.21974. PMID: 19226510.
Bibliografia
edytuj- Mark F. Bear, Barry W. Connors, Michael A. Paradiso: Neuroscience : exploring the brain. Philadelphia, PA: Lippincott Williams Wilkins, 2007. ISBN 0-7817-6003-8.
- Alan Longstaff: Neurobiologia. Warszawa: Wydawnictwo Naukowe PWN, 2012, s. 1-26. ISBN 978-83-01-13805-9.
- Piotr Jaśkowski: Neuronauka poznawcza. Vizja Press & IT, 2009, s. 20-25. ISBN 978-83-61086-51-2.
Linki zewnętrzne
edytujKarolina Głowacka i Jan Kamiński, Neurony i pamięć – nowe metody badań weryfikują dawne twierdzenia, kanał „Radio Naukowe” na YouTube, 12 września 2024 [dostęp 2024-09-16].
- Neuron – Scholarpedia (ang.)