Kentum- en satemtalen
De omvangrijke Indo-Europese talenfamilie wordt traditioneel onderverdeeld in twee grote groepen, de westelijke kentumtalen en oostelijke satemtalen. De benamingen zijn afgeleid van de Latijnse en Avestische woorden voor 'honderd', resp. centum ([kɛntum]?) en satem [satəm]?.

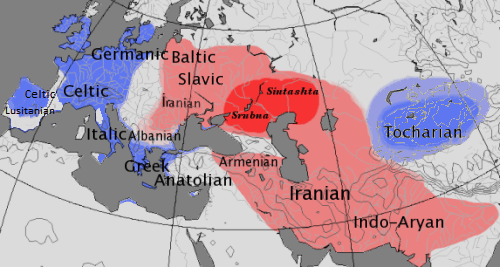
De onderscheiding tussen kentum- en satemtalen speelt geen rol meer in de historische taalwetenschap. Aan dit onderscheid werden vroeger belangrijke kenmerken verbonden, maar de vondst van oude of geïsoleerde talen zoals het Luwisch, Hettitisch en Tochaars wijst op een complexere evolutie langs meerdere ontwikkelingslijnen. Tochaars en Hettitisch werden namelijk gesproken in het oosten van het vermoede Indo-Europese taalgebied, hoewel ze kentumtalen waren. Aan de waarde van een specifieke beschrijving van veranderingen die zouden hebben geleid tot de Kentumgroep wordt getwijfeld. Daarentegen wordt algemeen geaccepteerd dat er in de vroege ontwikkeling van het Indo-Europees een klankwet werkzaam was waardoor in de talen waaruit zich het Sanskriet, het Iraans, het Slavisch en andere zogenaamde satemtalen vormden, de oorspronkelijke Proto-Indo-Europese palatovelaren veranderden in sibilanten (sisklanken). Deze ontwikkeling staat bekend als satemisering en kan worden teruggevoerd tot een kerngebied ten noorden van de Zwarte Zee, dat overeenkomt met de verspreiding van de jamnacultuur en dat geïdentificeerd is als de bakermat van de Indo-Iraanse volkeren.
Kentumtalen en satemtalen
bewerkenDe kentum-satemscheidslijn is de belangrijkste isoglosse van de Indo-Europese taalfamilie. Hierdoor worden twee hoofdgroepen afgebakend waarvan de namen zijn afgeleid van woorden voor het getal honderd, dat zich vanuit het Proto-Indo-Europese (PIE) *[ḱm̥tóm]? ontwikkelde tot respectievelijk het Latijnse centum [kɛntum]? en het Avestische [satəm]?. Het onderscheid concentreert zich rondom de evolutie uit het Proto-Indo-Europees van ten minste twee verschillende series velaren (veelal gutturale consonanten): de palatiovelaren (*ḱ, *ǵ en *ǵʰ) en de labiovelaren (*kʷ,*gʷ,*gʷʰ). Voor het proto-Albanees en in het Luwisch[1] zijn aanwijzingen gevonden dat er een onafhankelijke derde groep ‘gewone’ velaren (*k, *g en *gʰ) kan hebben bestaan, die dan vervolgens bij de kentumtalen zouden zijn samengesmolten met de PIE-palatovelaren en bij de satemtalen met de PIE-labiovelaren. Echter, behalve dat er te weinig gegevens bestaan om een dergelijke fundamenteel gescheiden ontwikkeling van de twee taalgroepen te kunnen afleiden, kan de ontwikkeling van de satemgroep uitstekend buiten de voorafgaande aanwezigheid van zo'n derde groep velaren. Algemeen wordt aangenomen dat de satemgroep zich heeft losgemaakt van de overige groepen, aangeduid als kentumtalen, door de velaren te palataliseren en vervolgens te veranderen in s-klanken, terwijl de labiale componenten van de PIE-labiovelaren verloren gingen en deze zich daarmee tot velaren ontwikkelden.
De kentumtalen omvatten de Italische talen / Romaanse talen, de Keltische talen, de Germaanse talen, het Grieks en mogelijk een aantal uitgestorven groepen waarvan maar weinig bekend is, zoals de Venetische talen, het vroegere Macedonisch en misschien de Illyrische talen. Het Tochaars had alle velaren-series samengevoegd en bewaard als velaren, en wordt op deze grond geclassificeerd bij de kentumgroep, zonder dat duidelijk is in welk stadium de PIE-labiovelaren zich tot velaren ontwikkeld hebben. De Anatolische talen hebben duidelijk kentumeigenschappen, maar worden vanwege een groot aantal goed gedocumenteerde archaïsche eigenschappen ook wel beschouwd als een aparte subgroep.
De satemtalen omvatten de Indo-Iraanse talen en het Balto-Slavisch (Baltische talen en Slavische talen). Verder worden gewoonlijk een aantal slecht gedocumenteerde uitgestorven talen tot de satemgroep gerekend, zoals de talen van de Thraciërs, de Daciërs en de Frygiërs. De classificatie van de Armeense taal als satemtaal is omstreden, omdat er sprake kan zijn van een secundaire assibilatie zoals die bv. ook in het Frans is opgetreden, maar het bewijs dat de sibilanten uit PIE-palatovelaren in plaats van uit een samengesmolten groep PIE-velaren/labiovelaren ontstaan kunnen zijn, berust op gegevens van te weinig woorden. Het Albanees gold traditioneel als een satemtaal en staat het dichtst bij de Balto-Slavische groep, maar wordt vanwege een aantal zeer archaïsche eigenschappen tegenwoordig liever in een aparte subgroep geplaatst.
Ongeacht of het verschil in kentumtalen en satemtalen voortkwam uit taal- of omgevingsfactoren, werd het onderscheid lang gezien als een vroege opsplitsing van het PIE in een westelijk en een oostelijk gebied. De ontdekking van de Tochaarse taal heeft geleid tot de concurrerende opvatting dat de satemisering een innovatie was die vanuit een centraal gebied uitwaaierde over de omringende IE-volkeren, maar daarbij de perifere gebieden in het westen (bv. de Keltische en Italische talen) en oosten (de Tochaarse talen) nooit heeft bereikt.
Indo-Europese expansie versus de expansie van de Satemgroep
bewerkenVolgens de oorspronkelijke koerganhypothese van Gimbutas, geïllustreerd door de kaart hiernaast, waren er een aantal fasen waarin de volkeren ten noorden van de Zwarte Zee, in de Pontisch-Kaspische Steppe van Oekraïne en Zuid-Rusland, zich over het Euraziatische continent verspreidden. Echter, ten westen hiervan, in het gebied dat zich uitstrekt van Nederland tot de bovenloop van de Wolga en de middenloop van de Dnjepr en van Scandinavië tot aan Zwitserland, bevond zich al de neolithische touwbekercultuur, die eveneens geldt als een drager van minstens een deel van de Indo-Europese cultuur. Gemengde grafvondsten van rond 2900 v.Chr. op de grens van de touwbekercultuur en de trechterbekercultuur in Nederland en Jutland suggereren een lokale continuïteit van Indo-Europese volkeren in noordwestelijk Europa die is terug te voeren tot aan het begin van de trechterbekercultuur, variërend van 3500 v.Chr. in Nederland tot ruim 4300 v.Chr. elders. Deze lokale continuïteit doet niet onder voor die van de Indo-Europese volkeren in de steppen, waardoor in archeologische kringen de kurganhypothese deels als achterhaald wordt beschouwd.[2]
Ook is er continuïteit van de touwbekercultuur met de kogelamforacultuur, die eveneens nog belangrijke agrarische elementen bezat en voor het eerst in 3500 v.Chr. in het noorden van Midden-Europa opkwam, vrijwel tegelijk met de koerganvolkeren van de Pontisch-Kaspische steppen.
Vooral Alexander Häusler heeft zich ingezet tegen elke mogelijke culturele band tussen deze twee cultuurgebieden. Evzen Neustupný heeft zich stelselmatig verzet tegen de opvatting dat de touwbekercultuur een uitheems element in Noord-Europa vertegenwoordigde en suggereerde een lange lokale voorgeschiedenis die kon worden teruggevoerd tot het vroege neolithicum en de kopertijd. Over de kwestie bestaat nog geen enigheid. Buchvaldek merkte verwantschap op tussen de touwbekercultuur en de jamnacultuur en suggereert een gezamenlijk herkomstgebied vanuit een onbekend middelpunt tussen Wisła en Dnjepr. Anderen suggereerden dat de Baalbergse groep (4300-4000 v.Chr.) in centraal Duitsland en Bohemen een inheemse overgangscultuur was tussen de Noord-Europese trechterbekercultuur en de kogelamforacultuur.
Overigens was er volgens de koerganhypothese van Gimbutas niets wat tegen een lokale ontwikkeling van de touwbekercultuur van Noord-Europa pleit, zij het dat volgens haar een heersende klasse vanuit de Pontisch-Kaspische steppen al deze culturen heeft ver-Indo-Europeaniseerd in een proces dat al moet zijn ingezet tijdens de jamnacultuur. Lichardus adopteerde het oude idee dat de trechterbekercultuur uit het oosten kwam en wees als oorsprong zowel de Dnjepr-Donetscultuur als de Sredny Stogcultuur aan, hetgeen weer even hard werd ontkend door sovjetarcheologen.
Uiteindelijk sloot Gimbutas niet uit dat de "eerste migratiegolf" van steppenvolkeren die tussen 4500-4000 v.Chr. Zuidoost-Europa hybridiseerde een domino-effect had op de ontwikkeling van meer westelijk gelegen volkeren. Mallory gaf toe dat deze theorie niet verkeerd was, maar dat er onvoldoende bewijs was om zulke processen te kunnen evalueren. Deze zogenoemde koerganisering is taalkundig nog altijd een zeer schaars beschreven proces en archeologisch nog altijd onvoldoende ondersteund door vondsten die zouden wijzen op grootschalige vermenging met volkeren afkomstig uit de steppen.
De massale overgang van de vroege Indo-Europeanen naar herdersvolken verliep over een enorm gebied en is snel verlopen. In Nederland spreekt men over hoogstens twee generaties, onder invloed van (religieuze, sociale of economische) impulsen van trechterbekervolkeren uit Oost-Duitsland (de Baalbergecultuur). Van deze herdersvolkeren zijn geen duidelijke nederzettingen bekend, maar de archeologische vondsten wijzen op een sterk gewijzigde religie, een patrilineaire samenleving en kenmerken die typisch zijn voor een strijdbaar volk. In het oosten werd deze cultuur vertegenwoordigd door de jamnacultuur, die veel kenmerken gemeen heeft met de westelijke touwbekerculturen zonder dat deze twee culturen gelden als wederzijds van elkaar afleidbaar.
Het door Gimbutas geschetste beeld van op paarden binnenvallende nomadische herdervolkeren uit de Pontisch-Kaspische steppen die in scherp contrast stonden met de meer sedentaire volkeren in het westen, werd genuanceerd door Renfrew,[3] die stelde dat zulke ruiters pas voor het eerst rond 1000 v.Chr. op het Europese toneel verschenen. Katrin Kell[4] wees erop dat het PIE getuigt van een aanzienlijke ontwikkeling op het gebied van de landbouw, alsook van een meer sedentaire leefwijze. Ook Mallory stelt dat de ontwikkeling naar het nomadische pastorale bestaan secundair moet zijn geweest. Of deze argumenten de jamnacultuur heeft gediskwalificeerd als unieke PIE-Urheimat of dat zulke diskwalificatie zou berusten op een verkeerde beeldvorming, feit is dat er een neiging bestaat om de werkelijke Urheimat verder terug in de tijd te zoeken, naar een cultuur die minder exclusief van de steppen was.
Voor de bepaling van het oorspronkelijke PIE-gebied moet de precieze relatie tussen de touwbekercultuur en de Pontisch-Kaspische steppencultuur worden vastgesteld, temeer omdat de verspreidingsgebieden iets van de Kentum-Satemtegenstelling lijkt te weerspiegelen. Vermeldenswaard is dat de tegenstanders van een koerganisering van Noord-Europa, vertegenwoordigd door Lothar Kilian en Alexander Häusler, een Proto-Indo-Europese "Urheimat" voorstelden die zowel Noord-Europa als de Pontisch-Kaspische steppen omvatte, en vele sovjetarcheologen de Pontisch-Kaspische steppen als bakermat van de Indo-Iraanse volkeren beschouwen. De bakermat van de overige Indo-Europese volkeren, in het bijzonder de Balto-Slavische volkeren, wordt volgens deze zienswijze daar noordwestelijk van gelokaliseerd. Dit, ondanks het commentaar van Mallory dat alle theorieën die uitgaan van een te groot verspreidingsgebied of van een taaldiffusie over een te lange tijdspanne, voorbijgaan aan de veranderlijkheid van een taal.
Linguïsten blijven nog in grote meerderheid achter de koergantheorie staan en tevens blijft het verspreidingsmodel van de koerganhypothese archeologisch voor de meeste satemtalen gewoon van toepassing. Toch hebben ook linguïsten aanmerkingen op de koerganhypothese. Frits Kortlandt stelde een herziening voor.[5] Hij noemt als belangrijkste bezwaar dat tegen het invasieschema van Gimbutas[6] kan worden ingebracht, dat het uitgaat van archeologisch bewijs en zoekt naar een linguïstische interpretatie. Daarentegen uitgaande van het linguïstische bewijs voor het samenvoegen van de puzzelstukjes, kwam hij tot het volgende beeld: Het gebied van de Sredny Stogcultuur in Oostelijk Oekraïne, waar landbouw nog samen met veeteelt voorkwam, noemt hij de overtuigendste kandidaat voor het oorspronkelijke Indo-Europese stamland. De Indo-Europeanen die na de migraties naar het westen, oosten en zuiden (zoals beschreven door Mallory[7]) overbleven werden de sprekers van Balto-Slavisch, terwijl de sprekers van de andere Satemtalen behoorden bij de jamnacultuurhorizon, en de westelijke Indo-Europeanen tot de touwbekercultuurhorizon. Terugkerend naar de Balten en Slaven, zouden hun voorouders gecorreleerd mogen worden met de Midden-Dnjeprcultuur. De gedachtegang van Mallory[8] volgend en aangenomen dat de oorsprong van deze cultuur te vinden is in de Sredny Stogcultuur, de jamnacultuur en de late Cucutenicultuur, stelt hij voor dat de loop van deze gebeurtenissen overeenkomt met de ontwikkeling van een Satemtaal die in de westelijke invloedssfeer werd getrokken.
De onvolledige satemisering in het Baltisch en, in minder mate, het Slavisch, omvat overblijfselen van labiale elementen uit de PIE-labiovelaren en wordt als aanwijzing gezien van een diffusie van de satem klankverschuivingen van buitenaf, naast een vroeg contact tussen Proto-Baltoslaven en Proto-Germanen.
Overigens gaat ook Kortlandt ervan uit dat er een algemene neiging bestaat om prototalen verder terug in de tijd te dateren dan wordt gegarandeerd door het linguïstische bewijs en er geen alternatief bestaat voor de zienswijze van Gimbutas over een secundair stamland in het westen: op deze grond sluit hij het Indo-Europese karakter van de Chvalynskcultuur aan de Midden-Wolga en de Majkopcultuur in de noordelijke Kaukasus uit. Hij stelt dat elke verwantschap die verder teruggaat dan de Sredny Stogcultuur te maken krijgt met de affiniteiten ten opzichte van andere prototalen, in welk verband hij de typologische overeenkomst tussen het Proto-Indo-Europees en de Noordwest-Kaukasische talen noemt en veronderstelt dat die kan worden toegeschreven aan regionale factoren, zodat het Indo-Europeaans een aftakking van het Oeraals zou kunnen zijn geweest dat zich onder invloed van een Kaukasisch substratum heeft ontwikkeld.
Genetische herkomst
bewerkenAntropologische gegevens wijzen erop dat de allereerste sprekers van de Indo-Europese talen voornamelijk behoorden tot het kaukasoïde (blanke) ras, cq. robuuste "cro-magnoïde" types vergelijkbaar met het langschedelige (dolichocefale) "noordelijke" subras en variaties daarvan in meer oostelijke richting.[9]
Vanaf de steentijd lijkt het kortschedelige (brachycefale) type in Europa een opmars te hebben gemaakt, waardoor dit type zich uiteindelijk als een wig tussen de noordelijke en zuidelijke langschedelige typen heeft gevestigd en er ook overal elders een sterke vermenging is opgetreden. Over de precieze herkomst van dit brachycefale element is weinig bekend. Het type lijkt een nieuwe ontwikkeling te zijn dat ook gerelateerd werd aan de klokbekerculturen. Dit is tegenwoordig echter omstreden, en ook voor omgevingsfactoren wordt een belangrijke rol ingeruimd. Door deze verschuivingen is het niet eenvoudig, of onmogelijk, om de PIE-"Urheimat" nu nog aan de hand van raskenmerken alleen te bepalen. Evenmin valt het onderscheid naar herkomst op grond van ras te maken tussen de eerste sprekers van Kentumtalen en de eerste sprekers van Satemtalen.
Onderzoek van genetische markers op het Y-chromosoom heeft meer opgeleverd. Haplogroep R1a1 van het Y-chromosoom is het gevolg van een mutatie die ongeveer 10.000 jaar geleden plaatsvond, maar wordt in het bijzonder geassocieerd met de expansie van de jamnacultuur. Dit DNA wordt van vader op zoon overgedragen en is daarmee extra behulpzaam voor het bepalen van afstamming in een patriarchale samenleving, zelfs in gebieden waar het oorspronkelijke fenotype door vergaande vermenging vrijwel geheel verloren is gegaan. R1a1 wordt het meest aangetroffen in Polen, Rusland, Oekraïne en Hongarije, maar ook in Centraal- en West-Azië en India. In mindere mate komt deze marker voor in West-Europa, bijvoorbeeld Frankrijk en Engeland.[10] Een belangrijke concentratie bevindt zich in Scandinavië, waar 23.6% van de Noren, 18.4% van de Zweden, 16.5% van de Denen en 11% van de Saami deze marker bezit.[11]
De ouderdom van R1a1, de hoge concentraties ervan onder de niet-Indo-Europese Saami en het antropologische bewijs voor een pre-noords subras in Noorwegen[12] laten de mogelijkheid open van een oorspronkelijk verspreiding van het DNA die niet samenviel met de PIE Urheimat. Evenmin wordt het gebied waar de hoogste concentraties van dit DNA zijn aangetroffen, Polen, beschouwd als beste optie voor de PIE Urheimat. Waarschijnlijk zijn hogere concentraties van dit DNA tijdens de IE expansie ergens opgepikt en daarna verder verspreid. Het romantische beeld dat door de media[13] geschetst is over een een-op-eenrelatie tussen R1a1 en PIE, is niet meer dan dat. De verdeling van R1a1 volgt grotendeels de Kentum-Satem-isoglosse en valt beter te correleren met de satemisering dan met de eerdere "verindo-europeanisering" van westelijk en zuidoostelijk Europa.
Voetnoten
bewerken- ↑ Craig Melchert, 'Proto-Indo-European velars in Luvian', in: Calvert Watkins (red.), Studies in Memory of Warren Cowgill (1929-1985): Papers from the Fourth East Coast Indo-European Conference, Cornell University, June 6-9, 1985, Berlijn/New York: de Gruyter, 1987, p. 182–204.
- ↑ Pre- & protohistorie van de lage landen, onder redactie van J.H.F. Bloemers & T. van Dorp 1991. De Haan/Open Universiteit. ISBN 90 269 4448 9, NUGI 644
- ↑ Renfrew, C. 1999. Time Depth, Convergence Theory and Innovation in Proto-Indo-European: "Old Europe" as a PIE Linguistic Area, Journal of Indo-European Studies, 27 (3-4): p258-293
- ↑ Krell, Kathrin S. 1998. "Gimbutas' Kurgan-PIE Homeland Hypothesis: A Linguistic Critique". In Roger Blench and Mathew Spriggs (eds.) Archaeology and Language, II:267-289. London: Routledge.
- ↑ Frederik Kortlandt, The spread of the Indo-Europeans, 2002. Gearchiveerd op 6 augustus 2023.
- ↑ (bv. 1985: blz. 198)
- ↑ J.P. Mallory, In search of the Indo-Europeans: Language, archaeology and myth. London: Thames and Hudson, 1989.
- ↑ (pp197f)
- ↑ Het oorspronkelijke antropologische type wordt geassocieerd met dat van de touwbekercultuur ("Corded" in de glossary van snpa), waaronder mediterrane mengvormen daarvan in de omgeving van de Pontische steppen van Oekraïne ("North-Pontid" in de glossary van snpa). Volgens antropologische opvattingen is het Noordse ras in zijn geheel een vrij recente mengvorm van robuuste, breedschedelige Cro-Magnoïde oudere vormen en slanke, hoog- en smalschedelige Mediterrane immigranten. Gearchiveerd op 15 augustus 2023.
- ↑ http://www.roperld.com/YBiallelicHaplogroups.htm, https://web.archive.org/web/20081201075030/http://mbe.oupjournals.org/cgi/content/full/19/7/1008
- ↑ http://www.oxfordancestors.com/papers/mtDNA04%20Saami.pdf
- ↑ "Valle Type" in de glossery van snpa (Society for Nordish Physical Anthropology)
- ↑ Atlas of the Human Journey - The Genographic Project
Referenties
bewerken- The New Encyclopædia Britannica, 15th edition.
- J.P. Mallory, In Search of the Indo-Europeans, Thames & Hudson, 1989, ISBN 0-500-27616-1.
- Frederik Kortlandt, The spread of the Indo-Europeans, 2002, [1].