Mesoderm


Mesoderm[2] (van Oudgrieks μέσος [mésos] “midden” en δέρμα [derma] “huid”) of mesoblast is het middelste kiemblad van een embryo. Bij mensen ontstaan de mesodermcellen in de derde week (vanaf de bevruchting) van de embryonale ontwikkeling door immigratie tussen de epiblast en de hypoblast (enterocoelia).
Het mesoderm differentieert in vier gebieden:
- een axiaal gebied, het axiaal mesoderm genoemd,
- een dorsaal gebied, het paraxiaal mesoderm of epimeer genoemd, dat aan elke kant van de neurale buis ligt;
- een middengebied, het tussenliggend mesoderm, mesomeer of nefrotoom genoemd, gelegen lateraal en ventraal van het paraxiaal mesoderm; een groot ventraal gebied aan elke kant van het archenteron,
- het mesoderma laminae lateralis of hypomeer genoemd.
Het mesoderm onderscheidt zich van de rest van het embryo door intercellulaire signalering, waarna het mesoderm gepolariseerd wordt door een organiserend centrum.[3] De positie van het organiserende centrum wordt op zijn beurt bepaald door de regio's waarin bèta-catenine wordt beschermd tegen afbraak door GSK-3. Bèta-catenine fungeert als een cofactor die de activiteit van de transcriptiefactor tcf-3 verandert van onderdrukken naar activeren, wat de synthese initieert van genproducten die cruciaal zijn voor mesodermdifferentiatie en gastrulatie. Bovendien heeft mesoderm het vermogen om de groei van andere structuren te induceren, zoals de neurale plaat, de voorloper van het zenuwstelsel.
Het mesoderm groeit uit tot:
- skelet
- bloedcellen
- bindweefsel van de darm en de huid
- mesenchym
- mesotheel
- spieren (hartspiercellen, gestreepte spieren en gladde spieren)
- peritoneum (buikvlies)
- voortplantingssysteem
- urinewegen en nieren
- chorda dorsalis
- somieten
Geschiedenis
[bewerken | brontekst bewerken]Heinz Christian Pander, een Baltische Duitser-Russische bioloog, wordt gecrediteerd voor de ontdekking van de drie kiembladen die zich vormen tijdens gastrulatie. Pander behaalde zijn doctoraat in zoölogie aan de Universiteit van Würzburg in 1817. Hij begon zijn studie in de embryologie met behulp van kippeneieren, waarbij hij het ectoderm, mesoderm en endoderm ontdekte. Vanwege zijn bevindingen wordt Pander soms de "grondlegger van de embryologie" genoemd.
Pander's werk op het gebied van het vroege embryo werd voortgezet door een Baltische-Duitse bioloog uit Pruisen-Estland genaamd Karl Ernst von Baer. Baer nam Pander's concept van de kiembladen over en door uitgebreid onderzoek naar veel verschillende soorten kon hij dit principe uitbreiden naar alle gewervelde dieren. Baer kreeg ook krediet voor de ontdekking van de blastula. Baer publiceerde zijn bevindingen, inclusief zijn kiembladentheorie, in een leerboek dat zich vertaalt naar Over de ontwikkeling van dieren, dat hij in 1828 uitbracht.[4]

Definitie
[bewerken | brontekst bewerken]Het mesoderm is een van de drie kiembladen die verschijnt bij de mens in de derde week van de embryonale ontwikkeling. Het wordt gevormd via een proces dat gastrulatie wordt genoemd. Er zijn vier belangrijke componenten, namelijk het axiaal, paraxiaal, tussenliggend en lateraal plaatmesoderm. Uit het axiale mesoderm ontstaat de chorda dorsalis. Het paraxiale mesoderm vormt de somitomeren, die zich organiseren in somieten in hoofd- en staartsegmenten, en aanleiding geven tot sclerotomen (kraakbeen en botten) en dermatomen (onderhuids weefsel).[5][6] Signalen voor somietdifferentiatie worden afgeleid van omringende structuren, inclusief de chorda dorsalis, neurale buis, en epidermis. Het tussenliggend mesoderm verbindt het paraxiale mesoderm met het mesoderma laminae lateralis. Uiteindelijk differentieert het in urogenitale structuren die bestaan uit de nieren, geslachtsklieren, de bijbehorende kanalen en de bijnierschors. Het mesoderma laminae lateralis geeft aanleiding tot het hart, de bloedvaten en de bloedcellen van de bloedsomloop, evenals tot de mesodermale componenten van de ledematen.[7]
Sommige van het mesoderm afgeleiden omvatten de spieren (gladde, hart- en skeletspieren), de spieren van de tong (hoofdsomieten), de keelholtespier (kauwspieren, spieren voor gezichtsuitdrukkingen), bindweefsel, de dermis en de onderhuidse laag van de huid, bot en kraakbeen, hard hersenvlies, het endotheel van de bloedvaten, rode bloedcellen, witte bloedcellen, microglia, de dentine van tanden, de nieren en de bijnierschors.[8]
Ontwikkeling bij de mens
[bewerken | brontekst bewerken]

Tijdens de derde week gerekend vanaf de bevruchting vormt een proces genaamd gastrulatie een mesodermale laag tussen het endoderm en het ectoderm. Dit proces begint met de vorming van een primitieve streep op het oppervlak van de epiblast.[9] De cellen van de lagen bewegen tussen de epiblast en de hypoblast, en beginnen zich lateraal en naar het hoofd (craniaal) te verspreiden. De cellen van de epiblast bewegen zich naar de primitieve streep en glijden eronder, in een proces dat "invaginatie" wordt genoemd. Sommige van de migrerende cellen verplaatsen de hypoblast en creëren het endoderm, en andere cellen migreren tussen het endoderm en de epiblast om het mesoderm te vormen. De overige cellen vormen het ectoderm. Daarna maken de epiblast en de hypoblast contact met het extra-embryonale mesoderm totdat ze de dooierzak en het amnion bedekken. Ze bewegen zich naar weerszijden van de lamina praechordalis. De prechordale cellen migreren naar de middellijn om de lamina notochorsalis te vormen. Het axiaal mesoderm is het centrale gebied van het rompmesoderm.[7] Dit vormt de chorda dorsalis, dat de vorming van de neurale buis induceert en de anterieur-posterieure lichaamsas tot stand brengt. De chorda dorsalis strekt zich uit onder de neurale buis van de kop tot de staart. Het mesoderm beweegt naar de middellijn totdat het de chorda dorsalis bedekt. Wanneer de mesodermcellen zich vermenigvuldigen, vormen ze het paraxiaal mesoderm. Aan elke kant blijft het mesoderm dun en staat bekend als het mesoderma laminae lateralis. Het tussenliggende mesoderm ligt tussen het paraxiaal mesoderm en het mesoderma laminae lateralis. Tussen dag 13 en 15 gerekend vanaf de bevruchting vindt de proliferatie van extra-embryonaal mesoderm, primitieve streep en embryonaal mesoderm plaats. Het chorda-dorsalisproces vindt plaats tussen dag 15 en 17 gerekend vanaf de bevruchting. Uiteindelijk vindt de ontwikkeling van het notochordkanaal en het axiale kanaal plaats tussen dag 17 en 19 gerekend vanaf de bevruchting, wanneer de eerste drie somieten worden gevormd.[10]
Axiaal mesoderm
[bewerken | brontekst bewerken]
Axiaal mesoderm of chordamesoderm is het mesoderm in het embryo dat langs de centrale as onder de neurale buis ligt.
Axiaal mesoderm:
- zal aanleiding geven tot de chorda dorsalis
- begint als een proces van de chorda dorsalis, waarvan de formatie bij mensen op dag 20 eindigt.
- is niet alleen belangrijk bij het vormen van de chorda dorsalis zelf, maar ook bij het induceren van de ontwikkeling van het bovenliggende ectoderm in de neurale buis
- zal uiteindelijk de wervels vormen.
- vormt aanvankelijk een cilindervorm. Deze cilinder fuseert (bij mensen rond de 20e dag) met het eronder liggende endoderm en vormt dan een structuur die de chordale plaat genoemd wordt.
- De chorda dorsalis zal de nucleus pulposus van de tussenwervelschijven vormen. Er is enige discussie over de vraag of deze cellen afkomstig uit de chorda dorsalis worden vervangen door andere uit het aangrenzende mesoderm.
Naast het paraxiaal mesoderm ligt het axiaal of chordamesoderm. De cellen van het chordamesoderm zijn dichter opeengepakt dan die van het paraxiaal mesoderm. Het chordamesoderm geeft aanleiding tot de lamina praechordalis, de chorda dorsalis en het dorsale mesoderm (axiaal mesoderm dat aanleiding geeft tot de somieten). Chordamesoderm is de bron van follistatine, cordine, noggin, Frzb en dickkopf, die respectievelijk de botmorfogenetische proteïne (BMP)- en Wnt-gradiënten van het ventrale mesoderm en endoderm tegenwerken. De belangrijkste afgeleide van het chordamesoderm dat belangrijk is voor het vormen van patronen in de dorso-ventrale as van de neurale buis is de chorda dorsalis.[11] Net als bij het paraxiaal mesoderm worden bij het chordamesoderm planaire celpolariteit (PCP)-signaleringscomponenten asymmetrisch gelokaliseerd op de voorste of achterste randen van intercalerende mediaal presomitische mesoderm-cellen bij het begin van convergentie en uitbreiding[12][13] en reguleren ze zowel gepolariseerd planair (horizontale vlak) als radiaal intercalatiegedrag.
Paraxiaal mesoderm
[bewerken | brontekst bewerken]
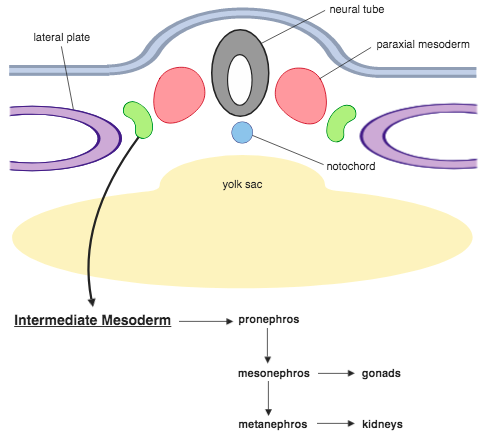

Tijdens de derde week gerekend vanaf de bevruchting wordt het paraxiaal mesoderm in segmenten georganiseerd. Als ze in het hoofdgebied verschijnen en in kopstaart richting groeien, worden ze somitomeren genoemd. Als ze in het hoofdgebied verschijnen maar contact maken met de neurale plaat, staan ze bekend als neuromeren, die later het mesenchym in het hoofd zullen vormen. De somitomeren organiseren zich in somieten die in paren groeien. In de vierde week verliezen de somieten hun organisatie en bedekken ze de chorda dorsalis en het ruggenmerg om de ruggengraat te vormen. In de vijfde week zijn er 4 occipitale somieten, 8 cervicale, 12 thoracale, 5 lumbale, 5 sacrale en 8 tot 10 coccygeale (stuitbeen) die het axiale skelet zullen vormen. Somitische afleidingen worden bepaald door lokale signalering tussen aangrenzende embryonale weefsels, in het bijzonder de neurale buis, chorda dorsalis, oppervlakte-ectoderm en de somitische compartimenten zelf.[14] De juiste specificatie van de afgeleide weefsels, het skelet, het kraakbeen, het endotheel en het bindweefsel, worden bereikt door een opeenvolging van morfogene veranderingen van het paraxiaal mesoderm, leidend tot de drie tijdelijke somitische lagen. Deze structuren worden gespecificeerd van dorsaal naar ventraal en van mediaal naar lateraal.[14] Elke somiet heeft de drie lagen: sclerotoom (dat wervels vormt), dermatoom (dat huid vormt) en myotoom (dat spieren vormt). Het myotoom is verdeeld in twee secties, het epimeer en het hypomeer, die respectievelijk de epaxiale en hypaxiale spieren vormen. De enige epaxiale spieren bij mensen zijn de musculus erector spinae (rugstrekker) en de kleine tussenwervelspieren, en worden geïnnerveerd door de dorsale takken van de spinale zenuwen. Alle andere spieren, inclusief die van de ledematen, zijn hypaxiaal en worden geïnnerveerd door de ventrale takken van de spinale zenuwen.[15]
Tussenliggend mesoderm
[bewerken | brontekst bewerken]Het tussenliggend mesoderm verbindt het paraxiaal mesoderm met het mesoderma laminae lateralis en differentieert zich in urogenitale structuren.[16] De voorlopernier (pronefros), de oernier (mesonefros) en de definitieve nier (metanefros) ontwikkelen zich bij gewervelde dieren uit het tussenliggend mesoderm. In de bovenste borstkas- en nekregio's vormt het tussenliggend mesoderm het nefrisch mesenchym als enkele langwerpige primordia. In de staartgebieden vormt het tussenliggend mesoderm de nefrogene streng, die een onderdeel is van de gonadale richel. De gonadale richel is de locatie van de embryonale geslachtsklieren.
Vroege vorming
[bewerken | brontekst bewerken]Factoren die de vorming van het tussenliggend mesoderm reguleren, worden niet volledig begrepen. Er wordt aangenomen dat botmorfogene eiwitten, of BMP's, groeigebieden langs de dorsaal-ventrale (achterkant-voorkant) as van het mesoderm specificeren en een centrale rol spelen bij de vorming van het tussenliggend mesoderm.[17] Vg1/Nodal-signalering is een geïdentificeerde regulator van tussenliggend mesodermvorming die werkt via BMP-signalering.[18] Overmatige Vg1/Nodal signalering tijdens vroege gastrulatie-stadia resulteert in uitbreiding van het tussenliggend mesoderm ten koste van het aangrenzende paraxiaal mesoderm, terwijl remming van Vg1/Nodal signalering de vorming van tussenliggend mesoderm onderdrukt.[19] Er is een relatie gevonden tussen Vg1/Nodal-signalering en BMP-signalering, waarbij Vg1/Nodal-signalering de tussenliggend mesodermvorming reguleert door de groei-inducerende effecten van BMP-signalering te moduleren.[19]
Andere noodzakelijke markers voor tussenliggend mesoderm-inductie zijn onder meer het protein odd-skipped-related 1 gen (Osr1) en het paired-box-2-gen (Pax2), waarvoor tussenliggende niveaus van BMP-signalering voor het activeren nodig zijn.[18] Markers van vroege tussenliggend mesodermvorming zijn vaak niet exclusief voor het tussenliggend mesoderm. Dit is te zien in de vroege stadia van tussenliggend mesoderm differentiatie, waarbij hogere niveaus van BMP de groei van lateraal plaatmesoderm stimuleren, terwijl lagere concentraties leiden tot paraxiaal mesoderm- en somiet-vorming.[20] Osr1, dat codeert voor een zinkvinger DNA-bindend eiwit, en LIM-type homeobox-gen (Lhx1) expressie overlapt zowel het tussenliggend mesoderm als het mesoderma laminae lateralis. Osr1 heeft expressiedomeinen die de gehele lengte van de anterior-posterieur-as (voorkant-achterkant-as) vanaf de eerste somieten omvatten. Pas in het 4e tot 8e somietstadium worden markers met een grotere specificiteit voor het tussenliggend mesoderm geïdentificeerd, waaronder Pax2/8-genen geactiveerd vanuit de 6e somiet (Bouchard, 2002). Lhx1 expressie wordt ook meer beperkt tot het tussenliggend mesoderm.[21] Genetische analyses in dierstudies laten zien dat Lhx1, Osr1 en Pax2/8 signalering allemaal cruciaal zijn bij de specificatie van het tussenliggend mesoderm tot zijn vroege afgeleiden.[20]
Mesoderma laminae lateralis
[bewerken | brontekst bewerken]
Het mesoderma laminae lateralis, zijplaatmesoderm of laterale plaat is het mesoderm dat zich aan de periferie van het embryo bevindt. Het bevindt zich aan de zijkant van het paraxiaal mesoderm en verder naar het axiale mesoderm. Het mesoderma laminae lateralis wordt gescheiden van het paraxiaal mesoderm door een smal gebied van tussenliggend mesoderm.
Het mesoderma laminae lateralis splitst zich in de pariëtale (somatisch mesoderm) en viscerale (splachnisch mesoderm) lagen. De pariëtale laag staat bekend als de somatopleura (mesenchyma somatopleurale) en de viscerale laag als de splanchnopleura (mesenchyma splanchnopleurale). De vorming van deze lagen begint met het verschijnen van intercellulaire holtes.[16] De twee lagen bedekken de intra-embryonale holte. De pariëtale laag vormt samen met het bovenliggende ectoderm de laterale lichaamswandplooien. De viscerale laag vormt de wanden van de darmbuis. Mesodermcellen van de pariëtale laag vormen de mesothelium-membranen of serosa-membranen, die de peritoneale, pleurale, pericardiale holtes en de binnenkant van de balzak bekleden.[5][6]. (Visceraal betekent dat de betreffende structuur gehecht is aan, of geassocieerd met de ingewanden en pariëtaal dat het gehecht is aan, of geassocieerd met de lichaamswand van borst of buik.) (De tunica serosa (ook wel serosa) is de gladde bekleding van de borstholte (cavitas pleuralis), de buikholte (cavitas peritonealis), het hartzakje (cavitas pericardialis) en het scrotum (cavitas serosa scroti).)

Cellen van het mesoderma laminae lateralis en het myotoom, de somiet die voor de spiervorming zorgt, migreren naar het ledemaatveld en prolifereren voor het vormen van de ledemaatknop. De lateraleplaatcellen produceren de kraakbeenachtige en skeletachtige delen van de ledematen, terwijl de myotoomcellen de spiercomponenten produceren. De mesodermcellen op de laterale plaat scheiden een fibroblastgroeifactor af (vermoedelijk FGF7 en FGF10) om het bovenliggende ectoderm ertoe aan te zetten een belangrijke organiserende structuur te vormen, de apicale ectodermale rand (AER) genoemd. De AER scheidt over en weer FGF8 en FGF4 af, waardoor het FGF10-signaal in stand wordt gehouden en proliferatie in het mesoderm wordt geïnduceerd. De positie van FGF10-expressie wordt gereguleerd door Wnt8c in de achterste en Wnt2b in de voorste ledematen. De voorste en de achterste ledematen worden gespecificeerd door hun positie langs de voorste/achterste as en mogelijk door twee T-boxen die transcriptiefactoren bevatten: respectievelijk Tbx5 en Tbx4.
Elke somiet heeft de drie lagen: sclerotoom (dat wervels vormt), dermatoom (dat huid vormt) en myotoom (dat spieren vormt).
Zie ook
[bewerken | brontekst bewerken]Externe link
[bewerken | brontekst bewerken]Literatuurverwijzingen
[bewerken | brontekst bewerken]- ↑ Vasiev, Bakhtier, Balter, Ariel, Chaplain, Mark, Glazier, James A., Weijer, Cornelis J. (2010). Modeling Gastrulation in the Chick Embryo: Formation of the Primitive Streak. PLOS ONE 5 (5): e10571. PMID 20485500. PMC 2868022. DOI: 10.1371/journal.pone.0010571.
- ↑ Everdingen, J.J.E. van, Eerenbeemt, A.M.M. van den (2012). Pinkhof Geneeskundig woordenboek (12de druk). Houten: Bohn Stafleu Van Loghum.
- ↑ Gastrulation: from cells to embryo. CSHL Press (2004), "Vertebrate Mesoderm Induction: From Frogs to Mice", p. 363. ISBN 978-0-87969-707-5.
- ↑ Baer KE von (1986) In: Oppenheimer J (ed.) and Schneider H (transl.), Autobiography of Dr. Karl Ernst von Baer. Canton, MA: Science History Publications.
- ↑ a b Ruppert, E.E., Fox, R.S., Barnes, R.D. (2004). Invertebrate Zoology, 7th. Brooks/Cole, "Introduction to Bilateria", 217–218. ISBN 978-0-03-025982-1.
- ↑ a b Langman's Medical Embryology, 11th edition. 2010.
- ↑ a b Scott, Gilbert (2010). Developmental biology, ninth. Sinauer Associates, US.
- ↑ Dudek, Ronald W. (2009). High-yield. Embryology, 4th. Lippincott Williams & Wilkins.
- ↑ Paraxial Mesoderm: The Somites and Their Derivatives. National Center for Biotechnology Information. Gearchiveerd op 4 mei 2022. Geraadpleegd op 15 april 2013.
- ↑ Drew, Ulrich (1993). Color atlas of embryology. Thieme, German.
- ↑ DEVELOPMENTAL BIOLOGY 3230 Neurogenesis. Gearchiveerd op 23 september 2024.
- ↑ Roszko I, Sawada A, Solnica-Krezel L. Regulation of convergence and extension movements during vertebrate gastrulation by the Wnt/PCP pathway. Semin Cell Dev Biol. 2009 Oct;20(8):986-97. doi: 10.1016/j.semcdb.2009.09.004. Epub 2009 Sep 15. PMID 19761865; PMC 2796982. Gearchiveerd op 24 april 2024.
- ↑ Chunyue Yin, Maria Kiskowski, Philippe-Alexandre Pouille, Emmanuel Farge, Lilianna Solnica-Krezel; Cooperation of polarized cell intercalations drives convergence and extension of presomitic mesoderm during zebrafish gastrulation . J Cell Biol 14 January 2008; 180 (1): 221–232. doi: https://doi.org/10.1083/jcb.200704150
- ↑ a b Yusuf, Faisal (2006). The eventful somite: Patterning, fate determination and cell division in the somite. Anatomy and Embryology 211 Suppl 1: 21–30. PMID 17024302. DOI: 10.1007/s00429-006-0119-8. ProQuest 212010706
- ↑ Sweeney, Lauren (1997). Basic Concepts in Embryology: A Student's Survival Guide, 1st Paperback. McGraw-Hill Professional. ISBN 9780070633087.
- ↑ a b Kumar, Rani (2008). Textbook of human embryology. I.K. International.
- ↑ Barak, Hila, Rosenfelder, Lea, Scultheiss, Thomas.M, Reshef, Ram (9 March 2005). Cell fate specification along the anterior–posterior axis of the intermediate mesoderm. Developmental Dynamics 232 (4): 901–904. PMID 15759277. DOI: 10.1002/dvdy.20263.
- ↑ a b Katsu, K, Tatsumi, N, Niki, D, Yamamura, K, Yokouchi, Y (1 februari 2013). Multi-modal effects of BMP signaling on Nodal expression in the lateral plate mesoderm during left-right axis formation in the chick embryo.. Developmental Biology 374 (1): 71–84. PMID 23206893. DOI: 10.1016/j.ydbio.2012.11.027.
- ↑ a b Fleming, BM, Yelin, R, James, RG, Scultheiss, TM (April 2013). A role for Vg1/Nodal signaling in specification of the intermediate mesoderm.. Development 140 (8): 1819–1829. PMID 23533180. PMC 3621495. DOI: 10.1242/dev.093740.
- ↑ a b Patel, SR, Dressler, GR (July 2013). The genetics and epigenetics of kidney development. Seminars in Nephrology 33 (4): 314–326. PMID 24011574. PMC 3767926. DOI: 10.1016/j.semnephrol.2013.05.004.
- ↑ Dressler, GR (Dec 2009). Advances in early kidney specification, development and patterning.. Development 136 (23): 3863–3874. PMID 19906853. PMC 2778737. DOI: 10.1242/dev.034876.