リポ多糖
リポ多糖(リポたとう、英: lipopolysaccharide, LPS、中: 脂多糖)は、グラム陰性菌細胞壁外膜の構成成分であり、脂質及び多糖から構成される物質(糖脂質)である。LPSは内毒素(エンドトキシン、英: endotoxin)であり、ヒトや動物など他の生物の細胞に作用すると、多彩な生物活性を発現する。LPSの生理作用発現は、宿主細胞の細胞膜表面に存在するToll様受容体 (英: Toll-like receptor、TLR) 4 (TLR4) を介して行われる。
構造・性質
[編集]
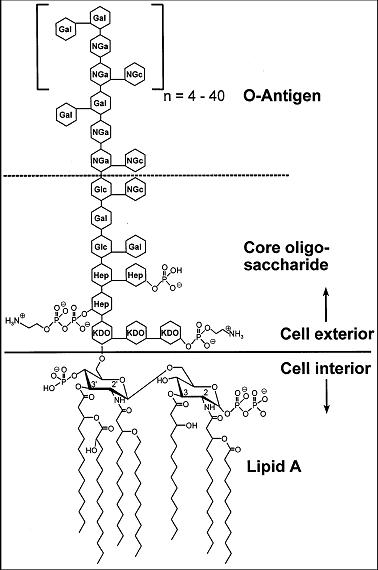
LPSはリピドAと呼ばれる脂質に、多分子の糖からなる糖鎖が結合した構造をとる(右図参照)。糖鎖部分は、コア多糖(またはコアオリゴ糖)と呼ばれる部分と、O側鎖多糖(O抗原)と呼ばれる部分から構成される。グラム陰性菌細胞壁のいちばん外側の部分には、外膜と呼ばれる脂質二重膜が存在しており、LPSは、リピドAの部分が、この脂質二重膜の外層を形成する分子として脂質層に入り込み、糖鎖の部分が細胞外に突き出す形で、グラム陰性菌の細胞表面に存在する。
O側鎖多糖は、3-5種類ほどの6炭糖(ヘキソース)や5炭糖(ペントース)からなる基本構造が、4-40回繰り返した構造を持つ。淋菌や髄膜炎菌など、菌種によっては特に短いO側鎖を有し、この場合はリポ多糖 (LPS) の代わりにリポオリゴサッカライド(リポオリゴ糖、LOS)と呼ぶことがある。O側鎖多糖の構造は、同種の細菌の中でも菌株ごとに異なり、菌体表面の親水性や抗原性に大きく関与する。O抗原と呼ばれる、細菌の表面抗原の本体であり、菌株の分類や鑑別に利用される。例えば大腸菌では約170種類の異なる型のO抗原が知られ、O抗原の違いからどのような病原性を持つ菌株か(O157など)を判別することができる(ただし、病原性自体はベロ毒素によるものであり、LPSの作用ではない)。また、O側鎖はファージに対する受容体にもなり、その菌に対してどのファージが感染しうるかを決定する因子として、ファージ型(phagovar)の決定にも関与する。
コア多糖は、5炭糖、6炭糖の他に、細菌に独特な7炭糖(ヘプトース)や8炭糖(特に2-ケトデオキシオクトン酸, KDO)などから構成される糖鎖である。O側鎖とは異なり、その構造は菌種によってほぼ一定である。コア多糖は疎水性の高い糖類から構成されており、親水性の高いO側鎖部分が短い場合には、細菌表面は疎水性になる。細菌表面の親水性が高い場合、しばしば寒天培地上のコロニーは表面が平滑で光沢を持つもの(S型、Smooth)に、疎水性が高い場合には表面が粗いもの(R型、Rough)なることが知られており、O側鎖およびコアからなる多糖の構成はコロニー性状にも影響する。
LPSの脂質部分に当たるリピドAはリン酸基が結合したグルコサミン2分子がグリコシド結合したものに対して脂肪酸鎖が複数結合した化学構造をとる。LPSの生理活性の発現において最も重要な働きをしていると考えられているのはリピドA部位であり、リピドA単独でも生理作用を示すことができる[1]。リピドAの構造中に含まれる脂肪酸鎖は細菌種により異なる。
LPSは細胞壁から容易には遊離せず、細菌が死滅したときなどに細胞が融解・破壊されることで遊離し、それが動物細胞などに作用することで毒性を発揮する。このような性質から、細菌が外に分泌する毒素(=外毒素)ではなく、分泌されない「菌体内に存在する毒素」、すなわち内毒素とも呼ばれる。LPSは熱的・化学的にも安定しており、通常の滅菌に用いられるオートクレーブや乾熱滅菌では不活化することができない。不活化には250℃で30分間の加熱を要する。
受容体
[編集]遊離したLPSは標的細胞の細胞膜に存在するTLR4を介してその作用を発現する。TLRファミリーは炎症性サイトカインの発現に関与し、自然免疫において重要な働きをしている。ヒトにおいては現在までにTLRファミリーに属する分子として10種類が知られている。TLRファミリーの細胞外ドメインはロイシンリッチリピート (LRR) と呼ばれる構造を持つ。LRRはアミノ酸の一つであるロイシンが規則的に配置されているロイシンリッチモチーフ(LRM)の繰り返し構造により構成される。また、細胞内ドメインはToll/IL-1R相同領域(TIRドメイン)と呼ばれ、インターロイキン-1 (IL-1) 受容体をはじめ、その他のIL-1受容体ファミリーに属する分子(IL-18受容体など)とも相同性を持つ領域である。また、近年ではTLR4以外にもLRRを有するNodという細胞内タンパク質がLPS受容体として働くことも報告されている[2]。
TLR4によるLPSの認識機構はまずLPS結合タンパク質 (LBP) によりLPSが捕捉され、CD14分子へと輸送されることにより開始される。さらにLPS-CD14複合体はTLR4に結合するわけであるが、その認識にはMD-2分子が必須である[3]。
シグナル伝達
[編集]
リガンドの結合による細胞内シグナル伝達経路はIL-1受容体と同様であり、以下の通りである。まず、TLR4にLPSが結合するとアダプタータンパク質であるミエロイド系分化因子88(英:Myeloid Differentiation Protein-88、MyD88)を介してセリン/スレオニンキナーゼであるIL-1受容体関連キナーゼ(英:IL-1 Receptor Associating Kinase、IRAK)を活性化する。さらにIRAKの下流にあるアダプタータンパク質TRAF-6 (TNF Receptor-associated Factor-6) を介して炎症反応に関与するNFκB (Nuclear Factor κB) やMAPキナーゼファミリー等の活性化を引き起こし、転写活性を示す[4]。
生理作用
[編集]LPSは上記に述べたシグナル伝達経路を介して種々の炎症性サイトカインの分泌を促進する作用を持つ。サイトカインの産生は細菌を除去するための生体防御反応として行われるが過剰になった場合に毒性が発現し、ショック状態に陥る(エンドトキシンショック)。また、LPSは抗原提示細胞である樹状細胞やマクロファージを活性化し、未分化なT細胞(ナイーブT細胞)を1型ヘルパーT細胞(Th1細胞)へと分化誘導する働きを持つ。このような作用を持つことからLPSは生物系の基礎研究においてin vivo及びin vitroの両方の系で炎症性刺激として多用されている。他にもLPSは発熱、多臓器不全、頻脈等の作用を有している。
自然界において、LPSは腸内細菌に由来するほか食用植物や漢方薬にも付着している。経口・経皮のLPSの自然摂取では毒性はなく、むしろ免疫系の成熟・調節に寄与していることを示す報告もある。たとえば、乳幼児期におけるLPSの自然摂取が、アレルギー体質になることを防いでいること[5]、生体内抗菌物質を誘導することで抗生物質耐性菌の繁殖を予防すること[6]、マウスにおける実験でLPSをインフルエンザの舌下ワクチンに添加することでIgGのほかIgA抗体の産生を高め予防効果が高いこと[7]、皮膚では、LPSのシグナル伝達が皮膚の創傷治癒やアレルギー抑制に必須であること[8][9]などが報告されている。さらに、炎症を抑制的に制御する制御性T細胞(Treg細胞)はLPSの刺激を受けることで、好中球の炎症誘導を抑制することが報告されている[10]。尚、外界との接点に存在する腸管マクロファージや皮膚のランゲルハンス細胞はLPSで刺激しても炎症性サイトカインを誘導しないことが報告されており[11][12]、LPSは生理的作用として生体恒常性維持に働く側面がある。
医薬への応用
[編集]LPS阻害薬であるポリミキシンBはファイザーから発売されているポリミキシンB錠やテラマイシン軟膏などの含有成分として医薬品に応用されている。
一方、LPSを医薬に用いている例として、サルモネラ菌由来のLPSのリピドAを脱リン酸化し、リン酸基を一つ残したモノフォスフォリルリピドA (MPL) [13]が、子宮頸がんワクチン(サーバリックス)のアジュバントとして使用されている[14]。
脚注
[編集]- ^ Galanos C, Lüderitz O, Rietschel ET, Westphal O, Brade H, Brade L, Freudenberg M, Schade U, Imoto M, Yoshimura H, Kusumoto S, Shiba T (1985). “Synthetic and natural Escherichia coli free lipid A express identical endotoxic activities”. Eur. J. Biochem. 148 (1): 1-5. doi:10.1111/j.1432-1033.1985.tb08798.x. PMID 2579812.
- ^ Inohara N, Ogura Y, Chen FF, Muto A, Nuñez G (2001). “Human Nod1 confers responsiveness to bacterial lipopolysaccharides”. J. Biol. Chem. 276 (4): 2551-2554. doi:10.1074/jbc.M009728200. PMID 11058605.
- ^ Shimazu R, Akashi S, Ogata H, Nagai Y, Fukudome K, Miyake K, Kimoto M (1999). “MD-2, a molecule that confers lipopolysaccharide responsiveness on Toll-like receptor 4”. J. Exp. Med. 189 (11): 1777-1782. doi:10.1084/jem.189.11.1777. PMC 2193086. PMID 10359581.
- ^ Li X, Qin J (2005). “Modulation of Toll-interleukin 1 receptor mediated signaling”. J. Mol. Med. 83 (4): 258-266. doi:10.1007/s00109-004-0622-4. PMID 15662540.
- ^ Braun-Fahrlander C, Riedler J, Herz U, Eder W, Waser M, Grize L, et al. (2002). “Environmental exposure to endotoxin and its relation to asthma in school-age children.”. The New England journal of medicine. 347 (12): 869-877. doi:10.1056/NEJMoa020057. PMID 12239255.
- ^ Brandl K, Plitas G, Mihu CN, Ubeda C, Jia T, Fleisher M, et al. (2008). “Vancomycin-resistant enterococci exploit antibiotic-induced innate immune deficits.”. Nature. 455 (7214): 804-807. doi:10.1038/nature07250.. PMC 2663337. PMID 18724361.
- ^ Fukasaka M, Asari D, Kiyotoh E, Okazaki A, Gomi Y, Tanimoto T, et al. (2015). “A Lipopolysaccharide from Pantoea Agglomerans Is a Promising Adjuvant for Sublingual Vaccines to Induce Systemic and Mucosal Immune Responses in Mice via TLR4 Pathway.”. PloS one. 10 (5): e0126849.. doi:10.1371/journal.pone.0126849.. PMC 4433252. PMID 25978818.
- ^ Brandt EB, Gibson AM, Bass S, Rydyznski C, Khurana Hershey GK. (2013). “Exacerbation of allergen-induced eczema in TLR4- and TRIF-deficient mice.”. Journal of immunology. 191 (7): 3519-3525. doi:10.4049/jimmunol.1300789.. PMC 3788607. PMID 23997219.
- ^ Chen L, Guo S, Ranzer MJ, DiPietro LA. (2013). “Toll-like receptor 4 has an essential role in early skin wound healing.”. The Journal of investigative dermatology. 133 (1): 258-267. doi:10.1038/jid.2012.267.. PMC 3519973. PMID 22951730.
- ^ Lewkowicz P, Lewkowicz N, Sasiak A, Tchorzewski H. (2006). “Lipopolysaccharide-activated CD4+CD25+ T regulatory cells inhibit neutrophil function and promote their apoptosis and death.”. Journal of immunology. 177 (10): 7155-7163. PMID 17082633.
- ^ Mitsui H, Watanabe T, Saeki H, Mori K, Fujita H, Tada Y, et al. (2004). “Differential expression and function of Toll-like receptors in Langerhans cells: comparison with splenic dendritic cells.”. The Journal of investigative dermatology. 122 (1): 95-102. doi:10.1046/j.0022-202X.2003.22116.x.. PMID 14962096.
- ^ Smythies LE, Sellers M, Clements RH, Mosteller-Barnum M, Meng G, Benjamin WH, et al. (2005). “Human intestinal macrophages display profound inflammatory anergy despite avid phagocytic and bacteriocidal activity.”. The Journal of clinical investigation. 115 (1): 66-75. doi:10.1172/JCI19229.. PMC 539188. PMID 15630445.
- ^ Baldridge JR, Crane RT. (1999). “Monophosphoryl Lipid A (MPL) Formulations for the Next Generation of Vaccines.”. Methods. 19 (1): 103–107. PMID 10525445.
- ^ Garçon N, Morel S, Didierlaurent A, Descamps D, Wettendorff M, Van Mechelen M. (2011). “Development of an AS04-adjuvanted HPV vaccine with the adjuvant system approach.”. BioDrugs. 25 (4): 217-226. doi:10.2165/11591760-000000000-00000.. PMID 21815697.
出典
[編集]- 杉原 久義、河野 恵、宿前 利郎 編集『第2版 新しい微生物学』廣川書店 1998年 ISBN 4567520548
- 吉田 眞一、柳 雄介、吉開 泰信 編集『改訂33版 戸田新細菌学』南山堂 2007年 ISBN 9784525160135