Corpo cetónico
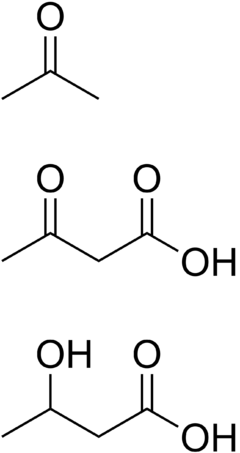
Os corpos cetónicos son compostos hidrosolubles, que se producen como subprodutos do catabolismo dos ácidos graxos, cando estes se utilizan para a produción de enerxía nas mitocondrias dos hepatocitos do fígado e nos riles. Quimicamente son cetonas, cetoácidos ou derivados de cetoácidos. Aínda que se denominen co termo "corpo", non son partículas, senón substancias disoltas. Utilízanse como fonte de enerxía no corazón e cerebro ou son produtos residuais do metabolismo, que son excretados. No cerebro son unha fonte vital de enerxía durante o xaxún.[1] Na diabetes mellitus tipo 1, pode acumularse no sangue unha excesiva cantidade de corpos cetónicos, producindo cetoacidose diabética.
Corpos cetónicos importantes
[editar | editar a fonte]Os máis importantes son os tres corpos cetónicos endóxenos orixinados no noso metabolismo, que son:[2]
- Acetona. É unha cetona. Orixínase porque unha parte (moi pequena en condicións normais) do acetoacetato sofre descarboxilación non encimática a acetona[3].
- Ácido acetoacético. A pH celular está ionizado como acetoacetato. É un ácido carboxílico cun grupo cetona, polo que é un cetoácido.
- Ácido beta-hidroxibutírico. A pH celular está ionizado como beta-hidroxibutirato. Tecnicamente non é unha cetona, senón un ácido carboxílico (beta-hidroxiácido), pero deriva de cetonas.
Outros corpos cetónicos como o beta-cetopentanoato e o beta-hidroxipentanoato poden orixinarse como o resultado do metabolismo de triacilglicéridos sintéticos como a triheptanoína, utilizada en certos tratamentos médicos [4].
Como se desprende das descricións anteriores, os corpos cetónicos agrúpanse nunha mesma categoría de moléculas porque derivan uns doutros e teñen características metabólicas e fisiolóxicas que así o aconsellan, pero, realmente, desde un punto de vista estritamente químico, non pertencen a un mesmo grupo de substancias químicas (hai unha cetona, un cetoácido e un hidroxiácido, o cal nin sequera ten un grupo cetona).
Utilización no corazón e cerebro
[editar | editar a fonte]Os corpos cetónicos poden utilizarse para a produción de enerxía. Os corpos cetónicos transpórtanse desde o fígado a outros tecidos, onde o acetoacetato e o beta-hidroxibutirato poden reconverterse en acetil-CoA para producir enerxía, por medio do ciclo do ácido cítrico. Os corpos cetónicos son solubles no plasma sanguíneo, polo que viaxan facilmente, pero, polo contrario, non se poderían transportar así os ácidos graxos, que son insolubles.
O corazón obtén unha parte pequena da súa enerxía dos corpos cetónicos, agás en circunstancias especiais; xeralmente utiliza principalmente os ácidos graxos.[5]
O cerebro obtén tamén parte da súa enerxía dos corpos cetónicos cando a glicosa está menos dispoñible (por exemplo, durante o xaxún, exercicios extenuantes, con niveis baixos de carbohidratos, en dietas cetoxénicas e nos neonatos). Cando hai unha baixada do nivel de azucre (glicosa) no sangue, moitos outros tecidos poden obter enerxía adicional a partir dos corpos cetónicos orixinados a partir dos ácidos graxos do propio tecido, pero o cerebro depende dos corpos cetónicos que chegan por vía sanguínea. Despois de que os niveis de glicosa baixen no sangue durante 3 días seguidos, o cerebro obtén o 25% da súa enerxía dos corpos cetónicos.[6] Despois de 4 días, a cifra sobe ao 70%. Durante as fases iniciais deste proceso o cerebro non consome os corpos cetónicos, xa que son importantes substratos para a síntese dos lípidos do cerebro.
Produción
[editar | editar a fonte]

A gran maioría dos corpos cetónicos orixínanse como resultado do metabolismo dos ácidos graxos (vía acetil-CoA), pero tamén se poden orixinar menores cantidades como resultado do metabolismo dos aminoácidos cetoxénicos.
No metabolismo de ácidos graxos vía acetil-CoA
[editar | editar a fonte]Os corpos cetónicos orixínanse a partir do acetil-CoA principalmente na matriz mitocondrial dos hepatocitos do fígado cando os carbohidratos son escasos e a enerxía debe obterse a partir da degradación dos ácidos graxos por beta-oxidación. Neste estado o nivel de acetil-CoA na célula é alto, o que fai que o complexo encimático da piruvato deshidroxenase se inhiba, e a piruvato carboxilase se active. O oxalacetato producido entra na gliconeoxénese en lugar de no ciclo do ácido cítrico, e, ademais, este está tamén inhibido polo elevado nivel de NADH, que se orixina como resultado da ß-oxidación dos ácidos graxos. Como nesas condicións o acetil-CoA non pode entrar no ciclo do ácido cítrico, o exceso de acetil-CoA utilízase para a produción de corpos cetónicos (cetoxénese). Isto ocorre en estado de xaxún.
A biosíntese mitocondrial de corpos cetónicos ocorre seguindo os seguintes pasos:
- A primeira reacción é a condensación de dúas moléculas de acetil-CoA para formar acetoacetil-CoA, pola acción do encima β-cetotiolase.
- A molécula de acetoacetil-CoA condensa con outra molécula de acetil-CoA para formar ß-hidroxi-ß-metilglutaril-CoA (HMG-CoA), por acción do encima HMG-CoA sintase.
- Finalmente, a HMG-CoA liase rompe o HMG-CoA nunha molécula de acetoacetato e outra de acetil-CoA.
A partir do acetoacetato vanse orixinar os outros corpos cetónicos. Unha parte do acetoacetato redúcese a β-hidroxibutirato na propia mitocondria, o que consome unha molécula de NADH; esta reacción é catalizada pola β-hidroxibutirato deshidroxenase que se encontra estreitamente asociada á membrana mitocondrial interna.
En condicións normais, unha mínima proporción de acetoacetato sofre unha lenta descarboxilación espontánea, non encimática, a acetona (o que significa que este corpo cetónico será degradado nunhas 5 horas se non se precisa para producir enerxía e o produto será eliminado como un residuo). A acetona non pode converterse de novo en acetil-CoA, polo que se excreta na urina, ou é exhalada ao respirar (como consecuencia da súa alta presión de vapor). A acetona é responsable do característico cheiro "a froita" do alento das persoas en estado de cetoacidose.[7]
No metabolismo de aminoácidos
[editar | editar a fonte]A degradación dos aminoácidos cetoxénicos orixina corpos cetónicos. Concretamente, os aminoácidos fenilalanina, tirosina, leucina, lisina e triptófano degrádanse orixinando acetoacetil-CoA no fígado, o cal pode converterse nos corpos cetónicos acetoacetato e beta-hidroxibutirato.
Algúns aminoácidos normalmente glicoxénicos que orixinan piruvato, especialmente alanina, cisteína e serina, poden formar acetoacetato a partir de acetil-CoA, particularmente en casos de diabete.
Cetose e cetoacidose
[editar | editar a fonte]A produción destes compostos chámase cetoxénese, e é un proceso necesario en pequenas cantidades. Os corpos cetónicos fórmanse en maiores cantidades en situacións nas que o metabolismo da glicosa non pode funcionar normalmente, como son: descompensación diabética (con cifras elevadas de glicosa en sangue), hipoglicemias, xaxún prolongado.
Cando se acumula un exceso de corpos cetónicos, este estado chámase cetose. A cetose pode cuantificarse analizando o aire que expulsa o paciente, e comprobando a presenza de acetona por cromatografía de gas.[8] Os diabéticos poden comprobar eles mesmos a presenza de acetona nos seus corpos utilizando kits para a análise de urina ou sangue.
Tanto o ácido acetoacético coma o ácido beta-hidroxibutírico, cando se acumulan grandes cantidades, fan que o pH sanguíneo baixe perigosamente (acidificación); este estado denomínase cetoacidose. Isto ocorre fundamentalmente na diabete tipo I non tratada, e menos frecuentemente na diabete tipo II (cetoacidose diabética) ou tamén na cetoacidose alcohólica. A causa desta acetoacidose é en ambos os casos a mesma: a célula non ten suficiente glicosa; no caso da diabete a falta de insulina evita que a célula reciba glicosa, entanto que no caso da cetoacidose alcohólica, a inanición provoca que haxa menos glicosa dispoñible en xeral.
Notas
[editar | editar a fonte]- ↑ Mary K. Campbell, Shawn O. Farrell (2006). Cengage Learning, ed. Biochemistry (5th ed.). pp. 579. ISBN 0534405215.
- ↑ Lori Laffel (1999). "Ketone bodies: a review of physiology, pathophysiology and application of monitoring to diabetes". Diabetes/Metabolism Research and Reviews 15 (6): 412–426. PMID 10634967. doi:10.1002/(SICI)1520-7560(199911/12)15:6<412::AID-DMRR72>3.0.CO;2-8.
- ↑ Devlin, T. M. 2004. Bioquímica, 4ª edición. Reverté, Barcelona. ISBN 84-291-7208-4
- ↑ Mochel F, DeLonlay P, Touati G, et al. (April 2005). "Pyruvate carboxylase deficiency: clinical and biochemical response to anaplerotic diet therapy". Mol. Genet. Metab. 84 (4): 305–12. doi:10.1016/j.ymgme.2004.09.007. PMID 15781190
- ↑ Kodde IF, van der Stok J, Smolenski RT, de Jong JW (2007). "Metabolic and genetic regulation of cardiac energy substrate preference". Comp. Biochem. Physiol., Part a Mol. Integr. Physiol. 146 (1): 26–39. PMID 17081788. doi:10.1016/j.cbpa.2006.09.014.
- ↑ Hasselbalch, SG; Knudsen, GM; Jakobsen, J; Hageman, LP; Holm, S; Paulson, OB (1994). "Brain metabolism during short-term starvation in humans.". Journal of cerebral blood flow and metabolism 14 (1): 125–31. PMID 8263048. doi:10.1038/jcbfm.1994.17.
- ↑ "American Diabetes Association-Ketoacidosis". Arquivado dende o orixinal o 29 de abril de 2010. Consultado o 14 de xaneiro de 2012.
- ↑ K. Musa-Veloso, S. S. Likhodii and S. C. Cunnane (2002). "Breath acetone is a reliable indicator of ketosis in adults consuming ketogenic meals". Am J Clin Nutr 76 (1): 65–70. PMID 12081817.
Véxase tamén
[editar | editar a fonte]Outros artigos
[editar | editar a fonte]Ligazóns externas
[editar | editar a fonte]- eMedicine|emerg|135 Diabetic Ketoacidosis
- Fat metabolism at unisanet.unisa.edu.au
- NHS Direct Online Health Encyclopaedia, Ketosis
- BioCarta Pathways Formation of Ketone Bodies
- MeshName - Ketone+Bodies [1]