Hepatitis delta virus ribozyme
| Hepatitis delta virus ribozyme | |
|---|---|
 Predicted secondary structure and sequence conservation of HDV ribozyme | |
| Identifiers | |
| Symbol | HDV_ribozyme |
| Rfam | RF00094 |
| Other data | |
| RNA type | Gene; ribozyme |
| Domain(s) | Viruses |
| SO | SO:0000374 |
| PDB structures | PDBe |
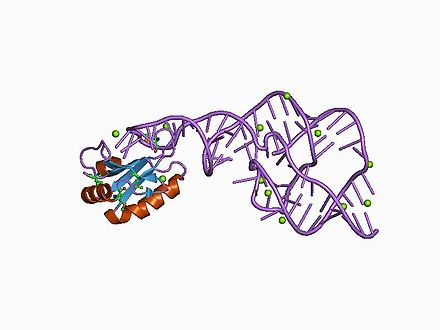
The hepatitis delta virus (HDV) ribozyme is a non-coding RNA found in the hepatitis delta virus that is necessary for viral replication. Hepatitis delta virus is the only known human virus that utilizes ribozyme activity to infect its host.[1] The ribozyme acts to process the RNA transcripts to unit lengths in a self-cleavage reaction during replication of the hepatitis delta virus, which is thought to propagate by a double rolling circle mechanism.[2][3] The ribozyme is active in vivo in the absence of any protein factors and was the fastest known naturally occurring self-cleaving RNA at the time of its discovery.[4]
The crystal structure of this ribozyme has been solved using X-ray crystallography and shows five helical segments connected by a double pseudoknot.[1]
In addition to the sense (genomic version), all HDV viruses also have an antigenomic version of the HDV ribozyme.[5] This version is not the exact complementary sequence but adopts the same structure as the sense (genomic) strand. The only "significant" differences between the two are a small bulge in P4 stem and a shorter J4/2 junction. Both the genomic and antigenomic ribozymes are necessary for replication.[2]
HDV-like ribozymes
[edit]The HDV ribozyme is structurally and biochemically related to many other self-cleaving ribozymes. These other ribozymes are often referred to as examples of HDV ribozymes, because of these similarities, even though they are not found in hepatitis delta viruses. They can also be referred to as "HDV-like" to indicate this fact.[6]
HDV-like ribozymes include the mammalian CPEB3 ribozyme, theta ribozymes in bacteriophages, retrotransposons members (e.g. in the R2 RNA element in insects and in the L1Tc and probably other retrotransposons in trypanosomatids) and sequences from bacteria.[7][8][6][9][10][11] The grouping is probably a result of convergent evolution: Deltavirus found outside of humans also possess a DV ribozyme, and no horizontal gene transfer scenarios proposed can yet explain this.[12][13][14]
Mechanism of catalysis
[edit]The HDV ribozyme catalyzes cleavage of the phosphodiester bond between the substrate nucleotide or oligonucleotide and the 5′-hydroxyl of the ribozyme. In the hepatitis delta virus, this substrate nucleotide sequence begins with uridine and is known as U(-1), however, the identity of the -1 nucleotide does not significantly change the rate of catalysis.[1] There is only a requirement for its chemical nature, since as shown by Perrotta and Been, substitution of the U(-1) ribose with deoxyribose abolishes the reaction, which is consistent with the prediction that the 2′-hydroxyl is the nucleophile in the chemical reaction.[15] Hence, unlike many other ribozymes, such as the hammerhead ribozyme, the HDV ribozyme has no upstream requirements for catalysis and requires only a single -1 ribonucleotide as a substrate to efficiently react.[1]
Initially, it was believed that the 75th nucleotide in the ribozyme, a cytosine known as C75, was able to act as a general base with the N3 of C75 abstracting a proton from the 2′-hydroxyl of the U(-1) nucleotide to facilitate nucleophilic attack on the phosphodiester bond.[1] However, although it is well established that the N3 of C75 has a pKa perturbed from its normal value of 4.45 and is closer to about 6.15 or 6.40,[16][17] it is not neutral enough to act as a general base catalyst. Instead, the N3 of C75 is believed to act as a Lewis acid to stabilize the leaving 5′-hydroxyl of the ribozyme; this is supported by its proximity to the 5′-hydroxyl in the crystal structure.[1][18] Substitution of the C75 nucleotide with any other nucleotide abolishes or substantially impairs ribozyme activity, although this activity can be partially restored with imidazole, further implicating C75 in catalytic activity.[19]
The C75 in the HDV ribozyme has been the subject of several studies because of its peculiar pKa. The typical pKa values for the free nucleosides are around 3.5 to 4.2; these lower pKa values are acidic and it is unlikely that they would become basic. However, it is likely that the structural environment within the ribozyme, which includes a desolvated active site cleft, provides negative electrostatic potential that could perturb the pKa of cytosine enough to act as a Lewis acid.[20][21][22][23]

In addition to Lewis acid stabilization of the 5′-hydroxyl leaving group, it is also now accepted that the HDV ribozyme can use a metal ion to assist in activation of the 2′-hydroxyl for attack on the U(-1) nucleotide. A magnesium ion in the active site of the ribozyme is coordinated to the 2’-hydroxyl nucleophile and an oxygen of the scissile phosphate, and may act as a Lewis acid to activate the 2′-hydroxyl.[19][18] In addition, it is possible that the phosphate of U23 can act as a Lewis acid to accept a proton from the 2′-hydroxyl with the magnesium serving as a coordinating ion.[24] Because the HDV ribozyme does not require metal ions to have activity, it is not an obligate metalloenzyme, but the presence of magnesium in the active site significantly improves the cleavage reaction. The HDV ribozyme does seem to have a nonspecific requirement for low amounts of divalent cations to fold, being active in Mg2+, Ca2+, Mn2+, and Sr2+.[1] In the absence of metal ions, it seems likely that water can replace the role of magnesium as a Lewis acid.
Regulation by upstream RNA
[edit]As limited by the rapid self-cleaving nature of HDV ribozyme, the previous ribonuclease experiments were performed on the 3′ product of self-cleavage rather than the precursor.[25] However, flanking sequence is known to participate in regulating the self-cleavage activity of HDV ribozyme.[26][27][28] Therefore, the upstream sequence 5′ to the self-cleavage site has been incorporated to study the resultant self-cleavage activity of the HDV ribozyme.[26] Two alternative structures have been identified.
The first inhibitory structure is folded by an extended transcript (i.e. -30/99 transcript, coordinates are referenced against the self-cleavage site) spanning from 30 nt upstream of the cleavage site to 15 nt downstream of the 3′-end.[26] The flanking sequence sequesters the ribozyme in a kinetic trap during transcription and results in the extremely diminished self-cleavage rate.[26] This self-cleavage-preventing structure includes 3 alternative stems: Alt1, Alt2 and Alt3, which disrupt the active conformation. Alt1 is a 10-bp Long-Range-Interaction formed by an inhibitory upstream stretch (-25/-15 nt) and the downstream stretch (76/86 nt).[26] The Alt1 disrupts the stem P2 in the active conformation wherein P2 is proposed to have an activating role for both genomic and antigenomic ribozyme.[26][29][30] Alt2 is an interaction between upstream flanking sequence and the ribozyme, and Alt3 is a nonnative ribozyme-ribozyme interaction.[26]
The secondary structure of this inhibitory conformation is supported by various experimental approaches.[26] First, direct probing via ribonucleases was performed and the subsequent modeling via mfold 3.0 using constraints from the probing results agrees with the proposed structure.[26] Second, a series of DNA oligomer complementary to different regions of AS1/2 were used to rescue the ribozyme activity; the results confirms the inhibitory roles of AS1/2.[26] Third, mutational analysis introduces single/double mutations outside the ribozyme to ensure the observed ribozyme activity is directly associated with the stability of the Alt1.[26] The stability of AS1 is found to be inversely related to the self-cleavage activity.[26]
The second permissive structure enables the HDV ribozyme to self-cleave co-transcriptionally and this structure further includes the -54/-18 nt portion of the RNA transcript.[26] The upstream inhibitory -24/-15 stretch from the aforementioned inhibitory conformation is now sequestered in a hairpin P(-1) located upstream of the cleavage site.[26][31][32] The P(-1) motif, however, is only found in the genomic sequence, which may be correlated with the phenomenon that genomic HDV RNA copies are more abundant in the infected liver cells.[26][5] Experimental evidence also supports this alternative structure. First, structural mapping via ribonuclease is used to probe the -54/-1 fragment instead of the whole precursor transcript due to the fast-cleaving nature of this structure, which agrees with the local hairpin P(-1) (between -54/-40 and -18/-30 nt).[26] Secondly, evolutionary conservation is found in P(-1) and the linking region between P(-1) and P1 among 21 genomic HDV RNA isolates.[26]
Use in RNA transcript preparation
[edit]The special properties of the HDV ribozyme's cleavage reaction make it a useful tool to prepare RNA transcripts with homogenous 3′ ends, an alternative to transcription of RNA with T7 RNA polymerase than can often produce heterogenous ends or undesired additions. The cDNA version of the ribozyme may be prepared adjacent to cDNA of the target RNA sequence and RNA prepared from transcription with T7 RNA polymerase. The ribozyme sequence will efficiently cleave itself with no downstream requirements, as the -1 nucleotide is invariant, leaving a 2′–3′ cyclic phosphate that can easily be removed by treatment with a phosphatase or T4 polynucleotide kinase.[33] The target RNA can then be purified with gel purification.
References
[edit]- ^ a b c d e f g h Ferré-D'Amaré AR, Zhou K, Doudna JA (October 1998). "Crystal structure of a hepatitis delta virus ribozyme". Nature. 395 (6702): 567–574. Bibcode:1998Natur.395..567F. doi:10.1038/26912. PMID 9783582. S2CID 4359811.
- ^ a b Modahl LE, Lai MM (July 1998). "Transcription of hepatitis delta antigen mRNA continues throughout hepatitis delta virus (HDV) replication: a new model of HDV RNA transcription and replication". Journal of Virology. 72 (7): 5449–5456. doi:10.1128/JVI.72.7.5449-5456.1998. PMC 110180. PMID 9621000.
- ^ Macnaughton TB, Shi ST, Modahl LE, Lai MM (April 2002). "Rolling circle replication of hepatitis delta virus RNA is carried out by two different cellular RNA polymerases". Journal of Virology. 76 (8): 3920–3927. doi:10.1128/JVI.76.8.3920-3927.2002. PMC 136092. PMID 11907231.
- ^ Kuo MY, Sharmeen L, Dinter-Gottlieb G, Taylor J (December 1988). "Characterization of self-cleaving RNA sequences on the genome and antigenome of human hepatitis delta virus". Journal of Virology. 62 (12): 4439–4444. doi:10.1128/JVI.62.12.4439-4444.1988. PMC 253552. PMID 3184270.
- ^ a b Chen PJ, Kalpana G, Goldberg J, Mason W, Werner B, Gerin J, Taylor J (November 1986). "Structure and replication of the genome of the hepatitis delta virus". Proceedings of the National Academy of Sciences of the United States of America. 83 (22): 8774–8778. Bibcode:1986PNAS...83.8774C. doi:10.1073/pnas.83.22.8774. PMC 387014. PMID 2430299.
- ^ a b Webb CH, Lupták A (2011). "HDV-like self-cleaving ribozymes". RNA Biology. 8 (5): 719–727. doi:10.4161/rna.8.5.16226. PMC 3256349. PMID 21734469.
- ^ Eickbush DG, Eickbush TH (July 2010). "R2 retrotransposons encode a self-cleaving ribozyme for processing from an rRNA cotranscript". Molecular and Cellular Biology. 30 (13): 3142–3150. doi:10.1128/MCB.00300-10. PMC 2897577. PMID 20421411.
- ^ Webb CH, Riccitelli NJ, Ruminski DJ, Lupták A (November 2009). "Widespread occurrence of self-cleaving ribozymes". Science. 326 (5955): 953. Bibcode:2009Sci...326..953W. doi:10.1126/science.1178084. PMC 3159031. PMID 19965505.
- ^ Sánchez-Luque FJ, López MC, Macias F, Alonso C, Thomas MC (October 2011). "Identification of an hepatitis delta virus-like ribozyme at the mRNA 5′-end of the L1Tc retrotransposon from Trypanosoma cruzi". Nucleic Acids Research. 39 (18): 8065–8077. doi:10.1093/nar/gkr478. PMC 3185411. PMID 21724615.
- ^ Sánchez-Luque F, López MC, Macias F, Alonso C, Thomas MC (January 2012). "Pr77 and L1TcRz: A dual system within the 5′-end of L1Tc retrotransposon, internal promoter and HDV-like ribozyme". Mobile Genetic Elements. 2 (1): 1–7. doi:10.4161/mge.19233. PMC 3383444. PMID 22754746.
- ^ Kienbeck K, Malfertheiner L, Zelger-Paulus S, Johannsen S, von Mering C, Sigel RK (February 2024). "Identification of HDV-like theta ribozymes involved in tRNA-based recoding of gut bacteriophages". Nat Commun. 15 (1): 1559. doi:10.1038/s41467-024-45653-w. PMC 10879173. PMID 38378708.
- ^ Hetzel U, Szirovicza L, Smura T, Prähauser B, Vapalahti O, Kipar A, Hepojoki J (April 2019). "Identification of a Novel Deltavirus in Boa Constrictors". mBio. 10 (2). doi:10.1128/mBio.00014-19. PMC 6445931. PMID 30940697.
- ^ Chang WS, Pettersson JH, Le Lay C, Shi M, Lo N, Wille M, Eden JS, Holmes EC (July 2019). "Novel hepatitis D-like agents in vertebrates and invertebrates". Virus Evolution. 5 (2): vez021. doi:10.1093/ve/vez021. PMC 6628682. PMID 31321078.
- ^ Paraskevopoulou S, Pirzer F, Goldmann N, Schmid J, Corman VM, Gottula LT, et al. (July 2020). "Mammalian deltavirus without hepadnavirus coinfection in the neotropical rodent Proechimys semispinosus". Proceedings of the National Academy of Sciences of the United States of America. 117 (30): 17977–17983. Bibcode:2020PNAS..11717977P. doi:10.1073/pnas.2006750117. PMC 7395443. PMID 32651267.
- ^ Perrotta AT, Been MD (January 1992). "Cleavage of oligoribonucleotides by a ribozyme derived from the hepatitis delta virus RNA sequence". Biochemistry. 31 (1): 16–21. doi:10.1021/bi00116a004. PMID 1731868.
- ^ Lee TS, Radak BK, Harris ME, York DM (2016). "A Two-Metal-Ion-Mediated Conformational Switching Pathway for HDV Ribozyme Activation". ACS Catalysis. 6 (3): 1853–1869. doi:10.1021/acscatal.5b02158. PMC 5072530. PMID 27774349.
- ^ Gong B, Chen JH, Chase E, Chadalavada DM, Yajima R, Golden BL, Bevilacqua PC, Carey PR (October 2007). "Direct measurement of a pK(a) near neutrality for the catalytic cytosine in the genomic HDV ribozyme using Raman crystallography". Journal of the American Chemical Society. 129 (43): 13335–13342. doi:10.1021/ja0743893. PMID 17924627.
- ^ a b Chen JH, Yajima R, Chadalavada DM, Chase E, Bevilacqua PC, Golden BL (August 2010). "A 1.9 A crystal structure of the HDV ribozyme precleavage suggests both Lewis acid and general acid mechanisms contribute to phosphodiester cleavage". Biochemistry. 49 (31): 6508–6518. doi:10.1021/bi100670p. PMID 20677830.
- ^ a b Nakano S, Proctor DJ, Bevilacqua PC (October 2001). "Mechanistic characterization of the HDV genomic ribozyme: assessing the catalytic and structural contributions of divalent metal ions within a multichannel reaction mechanism". Biochemistry. 40 (40): 12022–12038. doi:10.1021/bi011253n. PMID 11580278.
- ^ Rajagopal P, Feigon J (June 1989). "Triple-strand formation in the homopurine:homopyrimidine DNA oligonucleotides d(G-A)4 and d(T-C)4". Nature. 339 (6226): 637–640. Bibcode:1989Natur.339..637R. doi:10.1038/339637a0. PMID 2733796. S2CID 4313895.
- ^ Sklenár V, Feigon J (June 1990). "Formation of a stable triplex from a single DNA strand". Nature. 345 (6278): 836–838. Bibcode:1990Natur.345..836S. doi:10.1038/345836a0. PMID 2359461. S2CID 4233036.
- ^ Connell GJ, Yarus M (May 1994). "RNAs with dual specificity and dual RNAs with similar specificity". Science. 264 (5162): 1137–1141. Bibcode:1994Sci...264.1137C. doi:10.1126/science.7513905. PMID 7513905.
- ^ Legault P, Pardi A (September 1994). "In situ Probing of Adenine Protonation in RNA by 13C NMR". Journal of the American Chemical Society. 116 (18): 8390–8391. doi:10.1021/ja00097a066.
- ^ Kasprowicz A, Kempińska A, Smólska B, Wrzesiński J, Ciesiołka J (2015). "Application of a fluorescently labeled trans-acting antigenomic HDV ribozyme to monitor antibiotic–RNA interactions". Analytical Methods. 7 (24): 10414–10421. doi:10.1039/C5AY02953H.
- ^ Rosenstein SP, Been MD (October 1991). "Evidence that genomic and antigenomic RNA self-cleaving elements from hepatitis delta virus have similar secondary structures". Nucleic Acids Research. 19 (19): 5409–5416. doi:10.1093/nar/19.19.5409. PMC 328906. PMID 1923826.
- ^ a b c d e f g h i j k l m n o p q Chadalavada DM, Knudsen SM, Nakano S, Bevilacqua PC (August 2000). "A role for upstream RNA structure in facilitating the catalytic fold of the genomic hepatitis delta virus ribozyme". Journal of Molecular Biology. 301 (2): 349–367. doi:10.1006/jmbi.2000.3953. PMID 10926514.
- ^ Perrotta AT, Been MD (December 1990). "The self-cleaving domain from the genomic RNA of hepatitis delta virus: sequence requirements and the effects of denaturant". Nucleic Acids Research. 18 (23): 6821–6827. doi:10.1093/nar/18.23.6821. PMC 332737. PMID 2263447.
- ^ Perrotta AT, Been MD (April 1991). "A pseudoknot-like structure required for efficient self-cleavage of hepatitis delta virus RNA". Nature. 350 (6317): 434–436. Bibcode:1991Natur.350..434P. doi:10.1038/350434a0. PMID 2011192. S2CID 4331028.
- ^ Matysiak M, Wrzesinski J, Ciesiołka J (August 1999). "Sequential folding of the genomic ribozyme of the hepatitis delta virus: structural analysis of RNA transcription intermediates". Journal of Molecular Biology. 291 (2): 283–294. doi:10.1006/jmbi.1999.2955. PMID 10438621.
- ^ Perrotta AT, Nikiforova O, Been MD (February 1999). "A conserved bulged adenosine in a peripheral duplex of the antigenomic HDV self-cleaving RNA reduceskinetic trapping of inactive conformations". Nucleic Acids Research. 27 (3): 795–802. doi:10.1093/nar/27.3.795. PMC 148249. PMID 9889275.
- ^ Mathews DH, Sabina J, Zuker M, Turner DH (May 1999). "Expanded sequence dependence of thermodynamic parameters improves prediction of RNA secondary structure". Journal of Molecular Biology. 288 (5): 911–940. doi:10.1006/jmbi.1999.2700. PMID 10329189.
- ^ Zuker M, Mathews D, Turner D (1999). "Algorithms and thermodynamics for RNA secondary structure prediction practical guide". In Clark JB (ed.). RNA Biochemistry and Biotechnology. NATO ASI Series. Dordrecht, the Netherlands: Kluwer Academic Publishers.
- ^ Wichlacz A, Legiewicz M, Ciesiołka J (February 2004). "Generating in vitro transcripts with homogenous 3′ ends using trans-acting antigenomic delta ribozyme". Nucleic Acids Research. 32 (3): 39e–39. doi:10.1093/nar/gnh037. PMC 373431. PMID 14973333.