Profase

Profase (del grec; πρό (pró), «abans» + φάσις (phásis) «fase» ; «abans de la fase») és la primera etapa de la divisió cel·lular, tant en la mitosi com en la meiosi, que comença després de la fase G₂ de la interfase. L'ADN ja s'ha replicat quan la cèl·lula comença la profase. Els principals esdeveniment de la profase són la condensació de la cromatina i la desaparició del nuclèol.[2]
Mitosi
[modifica]La profase és la primera etapa de la mitosi a les cèl·lules animals i la segona etapa de la mitosi a les cèl·lules vegetals.[Nota 1][3] A l'inici de la profase, hi ha dues còpies idèntiques de cada cromosoma a la cèl·lula a causa de la replicació durant la interfase. Aquestes còpies es coneixen com a cromàtides germanes i estan unides per un element de l'ADN anomenat centròmer.[4]
-
Cicle cel·lular complet. La profase és una part de la mitosi (M) que comença després de la fase G₂ de la interfase
-
Canvis que es produeixen en una cèl·lula durant la mitosi (I,II,III: Profase; IV: Prometafase; V: Metafase; VI: Anafase; VII,VIII: Telofase)
-



Els principals esdeveniments de la profase són: la condensació de cromosomes, el moviment dels centrosomes, la formació del fus acromàtic, i l'inici de la ruptura del nuclèol.[2]
- La condensació dels cromosomes: L'ADN que s'ha replicat durant la interfase es condensa a partir de molècules amb longituds que arriben als 4 cm de llarg fins a cromosomes que es mesuren en micròmetres (µm).[2] Aquest procés utilitza el complex condensina.[4] Els cromosomes condensats consisteixen en dues cromàtides germanes unides al centròmer.[5]
- El moviment dels centrosomes: Durant la profase en cèl·lules animals, els centrosomes es comencen a desplaçar, allunyant-se entre ells.[2] L'activitat dels microtúbuls a cada centrosoma s'incrementa a causa de la recepció de la γ-tubulina. Els centrosomes replicats durant la interfase se separen cap als pols oposats de la cèl·lula amb l'ajut de les proteïnes motrius enganxades al centrosoma.[6] Els microtúbuls interpolars enganxats a cada centrosoma interaccionen entre ells, ajudant a traslladar els centrosomes cap als pols oposats.[2][6]
- La formació del fus acromàtic: Els microtúbuls implicats en les estructures temporals de la interfase es descomponen a mesura que se separen els centrosomes replicats.[2] El moviment dels centrosomes cap a pols oposats s'acompanya en les cèl·lules animals per l'organització de diferents matrius de microtúbuls radials (microtúbuls astrals) en cada centrosoma. Els microtúbuls interpolars dels dos centresomes interaccionen, unint els conjunts de microtúbuls i formant l'estructura bàsica del fus acromàtic. Les cèl·lules sense centríols, els cromosomes poden nuclear el conjunt de microtúbuls als fus acromàtic.[6] A les cèl·lules vegetals, els microtúbuls es reuneixen en pols oposats i comencen a formar el fus acromàtic en els llocs anomenats focus.[3] El fus acromàtic té una gran importància en el procés de la mitosi i acabarà separant les cromàtides germanes en la metafase.[2]
- Inici de la ruptura del nuclèol: El nuclèol comença a descompondre's en la profase, amb la qual cosa es deixa de produir ribosoma. Això indica una redirecció de l'energia cel·lular des del metabolisme cel·lular general cap a la divisió cel·lular.[2] La membrana nuclear es manté intacta durant aquest procés.[3]
Meiosi
[modifica]Durant la meiosi succeeixen dos processos de divisió del nucli, la meiosi I (dividida en profase I, metafase I, anafase I i telofase I), i la meiosi II (dividida en profase II, metafase II, anafase II i telofase II). Per tant la cèl·lula se sotmet dues vegades a la profase, donant lloc la profase I i la profase II.[5] La profase I és la fase més complexa de tota la meiosi perquè els cromosomes homòlegs s'han d'aparellar i intercanviar informació genètica.[2] La profase II és molt similar a la profase de la mitosi.[5]
Profase I
[modifica]La profase I es divideix en cinc estadis: leptotè, zigotè, paquitè, diplotè i diacinesi. A més dels esdeveniments que es produeixen en la profase de la mitosi, es produeixen diversos esdeveniments crucials dins d'aquests estadis, com l'aparellament de cromosomes homòlegs i l'intercanvi recíproc de material genètic entre aquests cromosomes homòlegs. La profase I es produeix a diferents velocitats en funció de l'espècie i el sexe. Moltes espècies aturen la meiosi a l'estadi diplotè de la profase I fins a l'ovulació.[2] Els humans, poden aturar els oocits a la profase I durant dècades i completar ràpidament la meiosi I abans de l'ovulació.[5]
-
Cromosomes en la fase zigotè
-
Cromosomes en la fase paquitè
-
Cromosomes en la fase diplotè
-
Cromosomes en la fase diacinesi




- Leptotè (del grec; λεπτος (leptos), «prim» + ταινία (tainía), «cinta» ; «cinta prima»): Al primer estadi de la profase I, els cromosomes (que són llargs, prims i sense espiralització) comencen a condensar-se. Cada cromosoma es troba en estat haploide i consta de dues cromàtides germanes; tanmateix, la cromatina de les cromàtides germanes encara no està suficientment condensada com per a ser visible al microscopi òptic.[2] Les regions homòlogues dins de parells de cromosomes homòlegs comencen a unir-se entre elles.[7]
- Zigotè (del grec; ζυγοῦν (zygoun), «que s'uneix» + ταινία (tainía), «cinta» ; «cinta que s'uneix»): Al segon estadi de la profase I, tots els cromosomes derivats maternalment i paternalment han trobat la seva parella homòlega.[2] A continuació, els parells homòlegs se sotmeten a una sinapsi per mitjà d'un complex sinaptinemal (una estructura proteïcea) que alinea les regions corresponents d'informació genètica sobre cromàtides no-germanes derivades maternalment i paternalment del parells de cromosomes homòlegs.[2] Els cromosomes homòlegs aparellats units pel complex sinaptinemal es coneix com a bivalents o tètrades.[2][3] Els cromosomes sexuals (X i Y) no realitzen totalment la sinapsi perquè només una petita regió dels cromosomes és homòloga.[2][5] El nuclèol es mou del centre cap a una posició perifèrica dins del nucli.[8]

- Paquitè (del grec; παχύς (pachys), «gruixut» + ταινία (tainía), «cinta» ; «cinta gruixuda»): Al tercer estadi de la profase I, que és el més llarg i el més estable, comença a completar-se la sinapsi.[2] La cromatina s'ha condensat prou com per poder-la veure amb un microscopi.[3] Les estructures anomenades nòduls de recombinació es formen sobre el complex sinaptinemal dels bivalents. Aquests nòduls de recombinació faciliten l'intercanvi genètic entre les cromàtides no-germanes del complex sinaptinemal en un esdeveniment conegut com a encreuament o recombinació genètica.[2] Es poden produir múltiples esdeveniments de recombinació a cada bivalent. En humans, es produeixen una mitjana de 2-3 esdeveniments en cada cromosoma.[6]
- Diplotè (del grec; διπλόος (diploos), «doble» + ταινία (tainía), «cinta» ; «doble cinta»): Al quart estadi de la profase I s'ha completat l'encreuament.[2][3] Els cromosomes homòlegs conserven un conjunt complet d'informació genètica; tanmateix, els cromosomes homòlegs són ara una barreja de descendència materna i paterna.[2] Les unions visibles anomenades quiasmes mantenen els cromosomes homòlegs en els llocs on es va produir la recombinació a mesura que el complex sinaptinemal es dissol.[2][5] És en aquest estadi on es produeix l'aturada de la meiosi en moltes espècies.[2]
- Diacinesi (del grec; δια (dia), «a través» + κίνημα (kínēma), «moviment» ; «moviment a través»): Al cinquè i últim estadi de la profase I ja s'ha produït una condensació completa de la cromatina i es poden observar les quatre cromàtides germanes en frma bivalent amb el microscòpi.
La resta de la profase I s'assembla a les primeres etapes de la prometafase de la mitosi, ja que la profase de la meiosi s'acaba amb l'inici de la formació del fus acromàtic i l'inici de la descomposició de la membrana nuclear.[2][3]
Profase II
[modifica]La profase II de la meiosi és molt similar a la profase de la mitosi. La diferència més notable és que la profase II es produeix amb un nombre haploide de cromosomes (n cromosomes) en contraposició al nombre diploide (2n cromosomes) de la profase de la mitosi.[3][5] Tant en cèl·lules animals com les cèl·lules vegetals, els cromosomes es poden descondensar durant la telofase I, requerint-los que es condensin de nou en la profase II.[2][3]
Si els cromosomes no necessiten tornar a condensar-se, la fase II procedeix sovint amb molta rapidesa, com es pot veure a l'organisme model Arabidopsis.[3]
Els esdeveniments nuclears
[modifica]Es pot observar la condensació progressiva de la cromatina dels cromosomes. Els cromosomes estan formats per dues cromàtides germanes (amb la mateixa informació genètica) que encara no podem visualitzar completament, ja que es troben estirades en tota la seva longitud per complexos proteics en forma d'anell, les cohesines (assegurant la cohesió de les cromàtides germanes des de la fase S fins a la transició de la metafase / anafase). Aquestes cohesines intervenen durant la meiosi / mitosi, però també durant la interfase realitzant diversos papers (regulació de la transcripció, reparació de l'ADN). Durant la metafase, les cohesineses substitueixen per condensines (responsables de la condensació) i es tradueix en un augment de la condensació i una associació de cromàtides germanes decreixents. Aquests canvis estan sota el control del complex CDK1-Ciclina B1 (fixa les condensines) i de la cinasa cdc5/Plk1 (elimina les cohesines).


També es pot observar una desaparició gradual de tots els constituents del nuclèol (a excepció del centre fibril·lar).
La membrana nuclear adopta un aspecte ondulat en determinades cèl·lules. Després de la profase, per tant durant la prometafase, la làmina nuclear desapareix, fent que la membrana nuclear es trenqui i es dissocii en «vesícules de membrana nuclear» (similars a les del reticle endoplasmàtic).
Els esdevediments citoplasmàtics
[modifica]Un orgànul (la definició d'orgànul varia entre els autors) important de la profase és el centrosoma. Igual que els cromosomes, el centrosoma es va duplicar durant les fases S i G2 de la interfase, abans de l'inici de la profase. A l'inici de la profase, el centrosoma conté quatre centríols, però es dividirà per la meitat per donar dos centrosomes i dos centríols. Cadascun dels dos centrosomes migrarà cap a un pol de la cèl·lula lliscant per sobre de la membrana nuclear. A partir de la següent fase, es trobaran posició diametralment oposada, als dos pols de la cèl·lula. Algunes cèl·lules eucariotes, sobretot les cèl·lules vegetals, estan desproveïdes de centríols.
El citoesquelet de microtúbuls es reorganitza per formar el fus acromàtic, una estructura bipolar que s'estén entre els dos centrosomes, i roman fora del nucli (en el citosol). Aquest fus acromàtic profàsic es descompon en microtúbuls astrals i microtúbuls interpolars:
- Els microtúbuls astrals es troben al voltant de cada centrosoma. Es tracta de fibres formades a partir de feixos de microtúbuls que surten del centrosoma en forma de raig, semblant a una estrella.
- Els microtúbuls interpolars permeten el creixement de l'eix mitòtic i, per tant, la migració dels centrosomes. Es pot observar una extrema inestabilitat d'aquests microtúbuls (el complex CDK1-Ciclina B1 fosforilitza diferents proteïnes associades als microtúbuls (MAPs)), compensat per un augment de la capacitat de nucleació dels centrosomes (augment del nombre de ƔTuRC). La cinesina 5 (o les proteïnes relacionades amb les cinesines) són les responsables de la separació dels centrosomes caminant cap a l'extrem positiu (+) dels microtúbuls. Aquestes fibres es qualifiquen de «contínues» a causa d'una il·lusió òptica present durant la seva observació amb un microscopi òptic.
Punts de control de la cèl·lula
[modifica]La profase I de la meiosi és la iteració més complexa de la profase que es produeix tant en cèl·lules vegetals com en cèl·lules animals.[2] Per garantir que aparegui correctament l'aparellament de cromosomes homòlegs i la recombinació del material genètic, hi ha punts de control cel·lulars. La xarxa de punts de control de la meiosi és un sistema de resposta al dany de l'ADN que controla la reparació de trencaments de doble cadena, l'estructura de la cromatina, i el moviment i l'aparellament de cromosomes.[9] El sistema consta de múltiples vies (incloent el punt de control de la recombinació meiòtica) que impedeixen que la cèl·lula entri en la metafase I amb errors a causa de la recombinació.[10]
Tinció i microscòpia
[modifica]Es pot utilitzar la microscòpia per visualitzar els cromosomes condensats a mesura que es mouen per meiosi o per mitosi.[11] S'utilitzen diverses tincions d'ADN per tractar cèl·lules de manera que es poden visualitzar els cromosomes condensats a mesura que es mouen durant la profase.[11]
S'utilitza habitualment la tinció del bandatge G per identificar cromosomes de mamífers; la utilització d'aquesta tecnologia sobre cèl·lules vegetals és difícil a causa de l'alt grau de compactació dels cromosomes en cèl·lules vegetals.[11][12] Les bandes G es va realitzar completament per als cromosomes vegetals el 1990.[13] Durant la profase meiòtica i mitòtica, es pot aplicar una tinció de Giemsa a les cèl·lules per obtenir una banda G en cromosomes.[7] La tinció de plata, una tecnologia més moderna, juntament amb la tinció de Giesma, es pot utilitzar per visualitzar el complex sinaptinemal en les diverses etapes de la profase meiòtica.[14] Per realitzar la banda G, s'han de fixar els cromosomes i, per tant, no és possible realitzar sobre cèl·lules vives.[15]
-
Dos nuclis de cèl·lules de ratolí en profase. Es poden veure els cromosomes condensats (vermell), la membrana nuclear (blau), i els microtúbuls (verd)
-
-
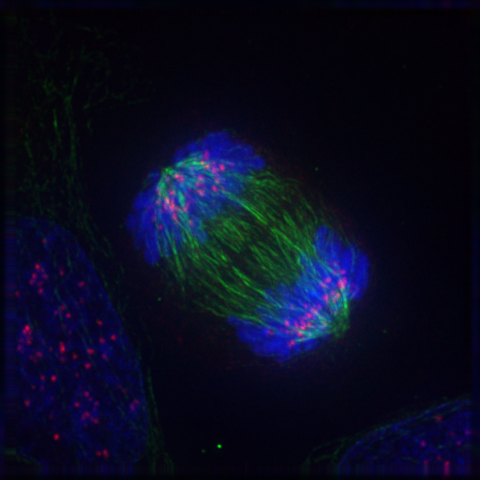
Imatge dels tres components d'una cèl·lula cancerosa humana amb càncer dividint-se; ADN (blau), una proteïna anomenada INCENP (verd) i els microtúbuls (vermell)



Es poden utilitzar tints fluorescents, com DAPI, tant en cèl·lules vives vegetals com en animals. En aquests tints no és necessari l'agrupament dels cromosomes, i permeten fer sondeigs d'ADN de regions específiques i de gens. L'ús de la microscòpia de fluorescència ha millorat àmpliament la resolució espacial.[16]
Notes
[modifica]- ↑ La diferència més notable entre la profase de cèl·lules vegetals i les cèl·lules animals es produeix perquè les cèl·lules vegetals no tenen centríols. En canvi, la membrana nuclear actua com un centre organitzador de microtúbuls (MTOC) per a la formació del fus acromàtic durant la preprofase. Una altra diferència notable és la preprofase, un pas addicional en la mitosi vegetal que es tradueix en la formació de la banda de preprofase, una estructura composta de microtúbuls. En la profase I de la meiosi de les plantes aquesta banda desapareix.[3]
Referències
[modifica]- ↑ Nussbaum, Robert L.; McInnes, Roderick R.; Huntington, F. Thompson & Thompson Genetics in Medicine (en anglès). Philadelphia: Elsevier, 2016, p. 12–20. ISBN 9781437706963.
- ↑ 2,00 2,01 2,02 2,03 2,04 2,05 2,06 2,07 2,08 2,09 2,10 2,11 2,12 2,13 2,14 2,15 2,16 2,17 2,18 2,19 2,20 2,21 2,22 2,23 Hartwell, Leland H; Hood, Leroy; Goldberg, Michael L; Reynolds, Ann E; et al.. Genetics From Genes to Genomes (en anglès). Nova York: McGraw-Hill, 2008, p. 90-103. ISBN 978-0-07-284846-5.
- ↑ 3,00 3,01 3,02 3,03 3,04 3,05 3,06 3,07 3,08 3,09 3,10 Taiz, Lincoln; Zeiger, Eduardo; Moller, Ian Max; Murphy, Angus. Plant Physiology and Development (en anglès). Sunderland MA: Sinauer Associates, 2015, p. 35–39. ISBN 978-1-60535-255-8.
- ↑ 4,0 4,1 Zeng, X.; Jiao, M.; Wang, X.; Song, Z.; Hao, S. «Electron microscopic studies on the Silver-stained Nucleolar Cycle of Physarum Polycephalum» (
PDF) (en anglès). Acta Botanica Cinica, 43(7), 2001, pàg. 680–685.
- ↑ 5,0 5,1 5,2 5,3 5,4 5,5 5,6 Nussbaum, Robert L; McInnes, Roderick R; Willard, Huntington F. Thompson & Thompson Genetics in Medicine (en anglès). Philadelphia: Elsevier, 2016, p. 12–20. ISBN 978-1-4377-0696-3.
- ↑ 6,0 6,1 6,2 6,3 Alberts, Bruce; Bray, Dennis; Hopkin, Karen; Johnson, Alexander; et al.. Essential Cell Biology (en anglès). Nova York NY: Garland Science, 2004, p. 639-658. ISBN 978-0-8153-3481-1.
- ↑ 7,0 7,1 Schermelleh, L.; Carlton, P. M.; Haase, S.; Shao, L.; et al. «Subdiffraction Multicolor Imaging of the Nuclear Periphery with 3D Structured Illumination Microscopy» (en anglès). Science, 320(5881), 2008, pàg. 1332–6. Bibcode: 2008Sci...320.1332S. DOI: 10.1126/science.1156947. PMID: 18535242.
- ↑ Zickler, D.; Kleckner, N. «The lepotene-zygotene transition of meiosis» (en anglès). Annu Rev Genet, 32, 1998, pàg. 619–697. DOI: 10.1146/annurev.genet.32.1.619. PMID: 9928494.
- ↑ Hochwagen, A; Amon, A «Checking your breaks: Surveillance mechanisms of meiotic recombination» (en anglès). Current Biology, 16(6), matç 2006, pàg. R217– R228. DOI: 10.1016/j.cub.2006.03.009. PMID: 16546077.
- ↑ MacQueen, Amy J; Hochwagen, Andreas «Checkpoint mechanisms: the puppet masters of meiotic prophase» (en anglès). Trends in Cell Biology, 21(7), 7-2011, pàg. 393–400. DOI: 10.1016/j.tcb.2011.03.004. PMID: 21531561.
- ↑ 11,0 11,1 11,2 Singh, Ram J. Plant Cytogenetics (en anglès). Boca Raton, FL: CBC Press, Taylor & Francis Group, 2017, p. 19. ISBN 9781439884188.
- ↑ Wang, H. C.; Kao, K. N. «G-banding in plant chromosomes» (en anglès). Genome, 30, 1988, pàg. 48–51. DOI: 10.1139/g88-009.
- ↑ Kakeda, K; Yamagata, H; Fukui, K; Ohno, M; et al. «High resolution bands in maize chromosomes by G-banding methods» (en anglès). Theor Appl Genet, 30, 1990, pàg. 265–272.
- ↑ Pathak, S; Hsu, T. C. «Silver-stained structures in mammalian prophase» (en anglès). Chromosoma, 70(2), 9-1978, pàg. 195–203. DOI: 10.1007/bf00288406. PMID: 85512.
- ↑ Sumner, A.T. «The nature and mechanisms of chromosome banding» (en anglès). Cancer Genetics and Cytogenetics, 6(1), 1982, pàg. 59–87. DOI: 10.1016/0165-4608(82)90022-x. PMID: 7049353.
- ↑ De Jong, Hans «Visualizing DNA domains and sequences by microscopy: a fifty-year history of molecular cytogenetics» (en anglès). Genome, 46(6), 12-2003, pàg. 943–946. DOI: 10.1139/g03-107. PMID: 14663510.
Vegeu també
[modifica]-
Profase
-
-
-
-




| Ciclina | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| CDK | |||||||||
| Inhibidor CDK | |||||||||
| Família P53 p63 p73 | |||||||||
| Altres | |||||||||
| Fases i punts de control |
| ||||||||