종양괴사인자
Tumor necrosis factor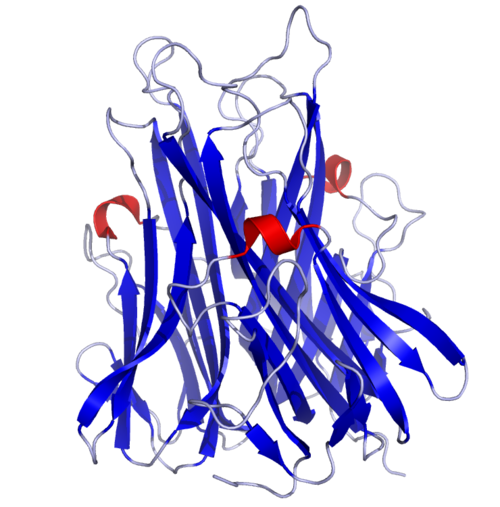





종양 괴사인자(TNF, 카섹신 또는 카섹틴, 종종 종양 괴사인자 알파 또는 TNF-α)는 아디포카인과 사이토카인이다.TNF는 상동 TNF 도메인을 가진 다양한 막 통과 단백질로 구성된 TNF 슈퍼 패밀리의 구성원이다.
아디포킨으로서 TNF는 인슐린 저항을 촉진하고 비만유발형 2형 [5]당뇨병과 관련된다.사이토카인으로서 TNF는 세포 신호 전달을 위해 면역계에 의해 사용된다.대식세포(특정 백혈구)가 감염을 감지하면 염증 [5]반응의 일부로 TNF를 방출하여 다른 면역 체계 세포에 경고를 보냅니다.
TNF 시그널링은 TNFR1과 TNFR2의 [6][7]두 가지 수용체를 통해 발생합니다.TNFR1은 대부분의 세포 유형에서 구성적으로 발현되는 반면, TNFR2는 주로 면역 세포의 [6][7]내피, 상피 및 서브셋으로 제한된다.TNFR1 시그널링은 항염증 및 아포토시스인 반면 TNFR2 시그널링은 항염증이며 세포 증식을 [6][7]촉진합니다.TNFR1 시그널링의 억제는 자가면역질환 [8]치료에 중요했지만 TNFR2 시그널링은 상처치유를 [7]촉진한다.
TNF-α 투과 성막 형태(mTNF-α)과 해결 가능한 형태(sTNF-α).. mTNF-α는 그것은 세포 간 접촉에 의해 조직 수용체와 상호 작용 monocytes/macrophages에서 찾을 수 있는 mTNF-α,[9]과정 기질 발표라 불리는 효소에 의해 효소의 분열에서 sTNF-α 결과 존재한다.[9]sTNF-α 선택적으로 TNFR1에, 모두에게mTNF-α 주는 반면에 바인딩 합니다.TNFR1 및 TNFR2.[10]TNFR1에 대한 TNF-α 결합은 불가역적인 반면 TNFR2에 대한 결합은 불가역적이다.[11]
TNF의 주된 역할은 면역세포의 조절이다.TNF는 내인성 발열제로서 발열, 아포토시스 세포사망, 악세사 및 염증을 유발하고 종양유전 및 바이러스 복제를 억제하며 IL-1 및 IL-6 생성세포를 통해 패혈증에 반응할 수 있다.TNF 생산 불량은 알츠하이머병,[12][13] 암, [14]주요 우울증, 건선[15], 염증성 [16]장질환을 포함한 다양한 인간의 질병과 관련이 있다.논란의 여지가 있지만, 일부 연구는 우울증과 IBD를 TNF의 증가 [17][18]수치와 연관짓고 있다.
Tasonermin이라는 이름으로 TNF는 특정 암 치료에 면역자극제로 사용된다.TNF의 작용에 대항하는 약물은 류마티스 관절염과 같은 다양한 염증성 질환의 치료에 사용된다.
특정 암은 TNF의 과잉 생산을 유발할 수 있다.TNF는 2차 고칼슘혈증을 일으키는 부갑상선 호르몬과 유사하며 과도한 생산과 관련된 암에서도 유사하다.
검출
생체 내 면역계의 항종양 반응 이론은 내과의사 윌리엄 B에 의해 인정되었다. 1968년 어바인 캘리포니아 대학의 Gale A Granger는 림프구에 의해 생성된 세포독성 인자를 보고하고 그것을 림프독소([19]LT)라고 명명했다.이 발견에 대한 공적은 예일 대학의 낸시 H 러들이 공유하고 있는데, 그는 같은 달에 [20]연속해서 발행된 일련의 기사에서 같은 활동을 보고했다.이후 1975년 뉴욕주 메모리얼 슬론-케터링 암 센터의 로이드 J. 올드(Lloyd J. Old)는 대식세포에 의해 생성된 또 다른 세포독성 인자를 보고하고 이를 종양괴사인자(TNF)[21]라고 명명했다.두 인자는 모두 생쥐 섬유육종 L-929 세포를 죽이는 능력에 기초해 기술되었다.이러한 개념은 1981년 호주 국립대학의 이안 A. 클라크가 Old의 그룹의 엘리자베스 카스웰과 협력하여 TNF의 과잉 생산이 말라리아 질병과 엔도톡신 [22][23]중독을 유발한다고 판단했을 때 전신 질환으로 확장되었다.
LT와 TNF를 코드하는 cDNA는 1984년에[24] 복제되었으며 유사한 것으로 밝혀졌다.TNF의 수용체에 대한 결합과 LT에 의한 변위는 두 요인 사이의 기능적 호몰로지를 확인하였다.TNF와 LT의 순차적 기능적 호몰로지는 TNFα로, LT는 TNFβ로 이름을 바꾸게 했다.1985년, 브루스 A. 뷰틀러와 앤서니 세라미는 카섹틴이 실제로 [25]TNF라는 것을 발견했다.그리고 그들은 TNF가 치사성 엔도톡신 [26]중독의 매개체임을 확인했다.케빈 J. Tracy와 Cerami는 치사성 패혈증 쇼크에서 TNF의 핵심 매개자 역할을 발견했으며 모노클로널 항TNF [27][28]항체의 치료 효과를 확인했다.
마크 매트슨 연구소의 연구는 TNF가 항산화 효소와 Bcl-2의 [29][30]발현을 유도하는 전사 인자 NF-γB의 활성화를 수반하는 메커니즘에 의해 뉴런의 죽음/아포토시스를 예방할 수 있다는 것을 보여주었다.
진
인간의 TNF 유전자는 [31]1985년에 복제되었다.그것은 6p21.3 염색체에 매핑되며, 약 3킬로베이스에 걸쳐 있으며 4엑손이 포함되어 있다.마지막 엑손은 림프토신 알파(LTA, 한때 TNF-β로 [32]명명됨)와 유사성을 공유한다.TNF의 3개의 주요 미번역 영역(3'-UTR)에는 AU가 풍부한 원소(ARE)가 포함되어 있습니다.
구조.
TNF는 주로 안정된 호모트리머로 [33][34]배열된 233-아미노산 길이 II형 트랜스막 단백질로 생산된다.이 막 일체형 형태에서 용해성 호모트리머사이토카인(sTNF)은 금속단백질가수분해효소 TNF 알파 변환효소(TACE, [35]ADAM17이라고도 함)에 의한 단백질 분해분열을 통해 방출된다.용해성 51kDa 삼량체 sTNF는 나노몰 범위 이하의 농도에서 분리되는 경향이 있으며, 이로 인해 생체 활성이 상실됩니다.인간 TNF의 분비 형태는 삼각형 모양의 피라미드 모양이며 무게는 약 17kDa이다.분비된 형태와 막 결합 형태 모두 생물학적으로 활성화되어 있지만, 각각의 특정 기능은 논란의 여지가 있다.하지만, 두 형태 모두 중복되고 뚜렷한 생물학적 [36]활동을 합니다.
일반적인 쥐의 TNF와 사람의 TNF는 구조적으로 다르다.[37]17킬로달톤(kDa) TNF 프로토머(185-아미노산 길이)는 반팔레알 β 가닥을 가진 2개의 반팔레알 β-환원 시트로 구성되며, TNF 계열에 전형적인 '젤리 롤' β 구조를 형성하지만 바이러스 캡시드 단백질에서도 발견된다.
세포 신호
tumornecrosisfactor2수용체, TNFR1(TNF 수용체 1;CD120a, p55/60)과 TNFR2(TNF 수용체 2;CD120b, p75/80)바인딩 할 수 있다.TNFR1은 55-kDa과 TNFR2은 75-kDa.[38]TNFR1 대부분의 조직에서 발생하고, TNFR2은 일반적으로 면역 체계의 세포에서 발견된다 양쪽 모두 충분히 그 막결합과 종양의 가용성 삼량체의. 형식에 의해, 활성화될 수 있고, 전구 핵부대 homotrimer의 막 경계 형태에 반응하다고 표현된다.으로서 종양 신호에 관한 대부분의 정보 TNFR1에서 파생된 것 TNFR2의 역할이 과소 평가다.부분에서 TNFR2이 세포 내 죽음 도메인이 있다면, 그것은neuroprotective 속성을 보여 줍니다.[30]

리간드와 접촉하면 TNF 수용체도 삼량체를 형성하며, TNF 단량체 사이에 형성된 홈에 팁이 들어맞습니다.이 결합은 수용체에서 구조 변화를 일으켜 억제 단백질 SOD를 세포 내 사망 도메인에서 분리시킨다.이 해리는 어댑터 단백질 TRADD가 사망 영역에 결합할 수 있도록 하며, 후속 단백질 결합을 위한 플랫폼 역할을 합니다.TRADD 결합에 이어 세 가지 경로를 [39][40]시작할 수 있습니다.
- NF-bB 활성화 : TRADD는 TRAF2와 RIP를 모집합니다.TRAF2는 다성분 단백질 키나제 IKK를 차례로 모집하여 세린-트레오닌 키나제 RIP가 활성화되도록 한다.통상 NF-δB에 결합해 그 전위를 억제하는 억제단백질 IδBα는 IKK에 의해 인산화되어 분해되어 NF-δB를 방출한다.NF-γB는 세포 생존과 증식, 염증 반응 및 항아포토시스 인자와 관련된 광범위한 단백질 배열의 전사를 매개하는 이단체 전사 인자입니다.
- MAPK 경로 활성화:세 가지 주요 MAPK 캐스케이드 중 TNF는 스트레스 관련 JNK 그룹의 강한 활성화를 유도하고 p38-MAPK의 중간 응답을 유발하며, 고전 ERK의 최소 활성화를 담당한다.TRAF2/Rac은 MLK2/MLK3,[41] TAK1, MEKK1 및 ASK1의 JNK 유도 업스트림 키나아제(각각 직접 또는 GCKs 및 Trx를 통해)를 활성화한다.SRC-Vav-Rac 축은 MLK2/MLK3를 활성화하고 이러한 키나아제들은 MKK7을 인산화하며, 이는 JNK를 활성화한다. JNK는 핵으로 전이되고 c-Jun 및 ATF2와 같은 전사 인자를 활성화한다.JNK 경로는 세포 분화, 증식에 관여하며 일반적으로 세포자멸을 촉진한다.
- 사망 신호 유도: 사망 도메인을 포함하는 모든 TNFR 슈퍼 패밀리의 구성원과 마찬가지로, TNFR1은 사망 [42]신호에 관여합니다.그러나 TNF 유도 세포사는 염증 과정에서의 압도적 기능에 비해 작은 역할을 할 뿐이다.다른 가족(Fas 등)에 비해 사망 유발 능력이 약하고 NF-δB의 항아포토시스 효과로 인해 종종 가려집니다.그럼에도 불구하고 TRADD는 FADD를 결합하고, FADD는 시스테인 단백질 분해효소 카스파아제-8을 신병합한다.카스파아제-8의 고농도는 카스파아제-8의 자동단백 활성화와 이펙터 카스파아제 분할을 유도하여 세포자멸을 일으킨다.
위의 경로에 의해 매개되는 무수하고 종종 충돌하는 효과는 광범위한 교차 대화의 존재를 나타낸다.예를 들어 NF-δB는 사망신호를 방해하는 억제단백질인 C-FLIP, Bcl-2 및 cIAP1/cIAP2의 전사를 촉진한다.한편 활성 캐스페이스는 RIP, IKK 및 NF-γB 자체의 서브유닛을 포함한 NF-γB 경로의 여러 성분을 분해한다.세포 유형, 다른 사이토카인의 동시 자극 또는 활성산소종(ROS)의 양과 같은 다른 요소들은 균형을 한 경로 또는 [citation needed]다른 경로로 바꿀 수 있습니다.이러한 복잡한 시그널링은 TNF가 방출될 때마다 매우 다양한 기능과 조건을 가진 다양한 세포가 [citation needed]염증에 적절하게 반응할 수 있음을 보증합니다.구강점막하섬유증의[43] 경우 단백질 분자 종양 괴사인자 알파와 케라틴 17이 모두 관련이 있는 것으로 보입니다
동물 모델에서 TNF는 선택적으로 자기반응성 T세포를 [44]죽인다.
또한 TNF-α 시그널링이 세포 [45][46][47][48]내 염증반응의 지속적인 향상을 초래하는 하류 후생유전학적 변형을 유발한다는 증거가 있다.
효소조절
이 단백질은 알로스테릭 [49]조절의 모르핀 모델을 사용할 수 있다.
임상적 의의
TNF는 주로 대식세포에 [50]의해 생성되는 것으로 생각되었지만 림프세포, 돛대세포, 내피세포, 심근세포, 지방조직, 섬유아세포, [51][unreliable medical source?]뉴런을 포함한 광범위한 세포 유형에 의해서도 생성된다.다량의 TNF는 리포다당류, 다른 세균 생성물 및 인터류킨-1(IL-1)에 반응하여 방출된다.피부에서 비만 세포는 염증 자극(예: LPS)[52]에 의해 방출될 수 있는 사전 형성 TNF의 주요 공급원으로 보인다.
일반적으로 IL-1 및 인터류킨-6(IL-6)과 함께 다양한 장기 시스템에 여러 가지 작용이 있습니다.
- 시상하부:
- 코르티코트로핀 방출 호르몬(CRH)의 분비를 자극하여 시상하부-하수체-부신축 자극
- 식욕을 억제하다
- 열.
- 간: 급성상 반응을 자극하여 C-반응성 단백질과 다른 매개체의 증가를 유도합니다.또한 인슐린 신호를 손상시키는 인슐린 수용체 기질-1(IRS-1)의 세린인산화를 촉진함으로써 인슐린 저항을 유도한다.
- 그것은 호중구에 대한 강력한 화학 유인제이며, 내피 세포에서 접착 분자의 발현을 촉진하여 호중구의 이동을 돕는다.
- 대식세포상 : 식세포증 촉진, IL-1 산화제 및 염증성 지질 프로스타글란딘 E22(PGE) 생성
- 다른 조직: 인슐린 저항성 증가.TNF는 인슐린 수용체 세린 잔기를 인산화하여 신호 전달을 차단합니다.
- 신진대사와 음식 섭취: 쓴맛 인식을 [53]조절합니다.
TNF 농도가 국소적으로 증가하면 염증의 주요 징후인 열, 붓기, 홍조, 통증 및 기능 상실이 발생합니다.
고농도의 TNF는 쇼크와 같은 증상을 유발하는 반면, 저농도의 TNF에 장기간 노출되면 소모성 증후군인 캐시샤를 초래할 수 있다.예를 들어, 이것은 암 환자들에게서 발견될 수 있다.
상기 등에서는 TNF가 단구에서의 PD-1 수치를 상향 조절함으로써 CD4 T세포의 확장 및 기능에 대한 IL-10 의존적 억제를 유발하고, PD-1이 [54]PD-L에 의해 결합된 후 단구들에 의해 IL-10이 생성되는 것으로 나타났다.
Pedersen 등의 연구에 따르면 패혈증에 대한 반응의 TNF 증가는 미오킨의 운동 유발 생성에 의해 억제된다.급성 운동이 진정한 항염증 반응을 유도하는지를 연구하기 위해, '저급 염증' 모델을 확립했다. 이 모델은 내독소 투여 전에 휴식 또는 운동을 무작위로 선택한 건강한 지원자에게 저용량의 대장균 내독소를 투여했다.휴식 중인 피험자에서 엔도톡신은 TNF의 순환 수치를 2~3배 증가시켰다.이와는 대조적으로 피험자가 3시간의 에르고미터 사이클링을 수행하고 2.5시간에 엔도톡신 볼루스를 투여받았을 때 TNF 반응은 완전히 [55]무뎌졌다.이 연구는 급성 운동이 TNF [56]생성을 억제할 수 있다는 몇 가지 증거를 제공한다.
뇌에서 TNF는 [30]흥분독성으로부터 보호할 수 있다.TNF는 시냅스를 [6]강화합니다.뉴런의 TNF는 그들의 생존을 촉진하는 반면, 대식세포와 마이크로글리아의 TNF는 아포토시스를 [30]유도하는 신경독을 일으킨다.
비만은 TNF-α 및 IL-6 농도가 높아진다.[57][58][59]TNF-α에 대한 모노클로널 항체는 비만의 감소보다는 증가와 관련이 있어 염증이 비만의 [59]원인이라기보다는 결과임을 나타낸다.TNF와 IL-6은 COVID-19 심각도와 [5]사망을 예측하는 가장 두드러진 사이토카인이다.
약리학
TNF는 염증 반응을 촉진하고, 이는 차례로 류마티스 관절염, 강직성 척추염, 염증성 장질환, 건선, 화농성 은색선염, 내화성 천식과 같은 자가면역 질환과 관련된 많은 임상적 문제를 일으킨다.이러한 장애들은 때때로 TNF 억제제를 사용하여 치료된다.이러한 억제는 TNF에 직접 결합하는 인플릭시맵(레미케이드)과 같은 모노클로널 항체, 아달리맵(휴미라), 서톨리주맵 페골(Cimzia)과 같은 TNF에 TFRN보다 높은 친화력으로 결합하는 유인 순환 수용체 융합 단백질(Etanercept, Enbrel)과 같은 유인체 순환 수용체 융합 단백질에 의해 달성될 수 있다.
한편, TNF 억제제로 치료된 일부 환자는 질병의 악화 또는 자가면역성의 새로운 시작을 일으킨다.TNF는 면역 억제 측면도 가지고 있는 것 같습니다.가능한 메커니즘의 한 가지 설명은 TNF가 종양 괴사인자 수용체 2(TNFR2)[61]에 결합하기 때문에 조절 T세포(Tregs)에 양성 영향을 미친다는 관찰이다.
항TNF 치료는 암 치료에서 약간의 효과만 보여왔다.인플릭시맙으로 신장세포암을 치료한 결과 일부 환자의 질병이 장기간 안정되었다.Etanercept는 IL-6와 CCL2의 하향조절을 통해 특정 환자의 유방암과 난소암에 대한 치료가 장기화되었는지 검사했다.한편, 진행성 췌장암 환자를 치료하기 위해 gemcitabine에 infliximab 또는 etanercept를 첨가하는 것은 [62]위약과 비교했을 때 효과의 차이와 관련이 없었다.
상호 작용
TNF는 TNFRSF1A와 [63][64]상호작용하는 것으로 나타났습니다.
명명법
LTα는 더 이상 TNFβ로 [65]지칭되지 않기 때문에, HGNC(HUGO 유전자 명명 위원회) 데이터베이스에 나타나 있듯이, 이전의 유전자 기호로서 TNFα는 단순히 TNF로 불린다.
레퍼런스
- ^ a b c ENSG00000230108, ENSG00000223952, ENSG00000204490, ENSG00000228321, ENSG00000232810, ENSG00000228849, ENSG00000206439 GRCh38: Ensembl 릴리즈 89: ENSG000000002278008989, ENSG
- ^ a b c GRCm38: 앙상블 릴리즈 89: ENSMUSG000024401 - 앙상블, 2017년 5월
- ^ "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- ^ "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- ^ a b c Sethi JK, Hotamisligil GS (2021). "Metabolic Messengers: tumour necrosis factor". Nature Metabolism. 3 (10): 1302–1312. doi:10.1038/s42255-021-00470-z. PMID 34650277.
- ^ a b c d Heir R, Stellwagen D (2020). "TNF-Mediated Homeostatic Synaptic Plasticity: From in vitro to in vivo Models". Frontiers in Cellular Neuroscience. 14: 565841. doi:10.3389/fncel.2020.565841. PMC 7556297. PMID 33192311.