혈관신생성
Angiogenesis| 혈관신생성 | |
|---|---|
 혈관형성에 따른 혈관형성 | |
| 해부학 용어 |

혈관신생은 혈관신생의 초기 단계에서 형성된 기존의 [1][2][3]혈관으로부터 새로운 혈관이 형성되는 생리적인 과정이다.혈관신생은 싹트고 [4]갈라지는 과정에 의해 혈관조직의 성장을 계속한다.혈관 형성은 중배엽 세포 전구체 [5]및 신생 혈관 형성으로부터 내피 세포의 태아 형성이다. 비록 논의가 항상 정확한 것은 아니지만(특히 오래된 문헌에서).발달 중인 배아의 첫 번째 혈관은 혈관 형성을 통해 형성되는데, 혈관 형성은 발달 과정과 [6]질병에서 혈관 성장을 전부는 아니더라도 대부분 담당합니다.
혈관신생은 성장과 발달, 상처 치유 및 과립조직 형성에 있어 정상적이고 중요한 과정이다.그러나 양성 상태에서 악성 상태로의 종양 이행의 기본 단계이기도 하며,[7] 암 치료에 혈관신생 억제제를 사용하게 된다.종양 성장에서 혈관신생의 본질적인 역할은 1971년 유다 포크먼에 의해 처음 제안되었는데, 그는 종양이 "뜨겁고 유혈"[8]이라고 묘사했고, 적어도 많은 종양 유형에서 홍조 관류, 심지어 충혈이 특징이라는 것을 보여주었다.
종류들
혈관 신생의 싹트기
싹트기 혈관신생은 혈관신생의 첫 번째 확인된 형태였고, 이로 인해 혈관신생은 관장성 혈관신생보다 훨씬 더 많이 이해되고 있다.그것은 몇 가지 특징적인 단계에서 발생합니다.초기 신호는 혈관 구조가 없는 조직 부위에서 발생합니다.이러한 영역에서 지적되는 저산소증은 조직이 신진대사 활동을 수행할 수 있도록 하는 영양소와 산소의 존재를 요구하게 만든다.이로 인해 실질세포는 혈관신생성장인자인 혈관내피성장인자([9]VEGF-A)를 분비한다.이러한 생물학적 신호는 기존의 혈관에 존재하는 내피세포의 수용체를 활성화시킨다.둘째, 팁 세포로도 알려진 활성화된 내피 세포는 원래 혈관 벽에서 내피 세포가 빠져나갈 수 있도록 하기 위해 기저막을 분해하는 단백질 분해효소라고 불리는 효소를 방출하기 시작합니다.내피 세포는 주변 기질로 증식하여 인접한 혈관을 연결하는 단단한 새싹을 형성합니다.증식하는 세포는 끝 세포 뒤에 위치하며 줄기 세포로 알려져 있습니다.이 세포들의 증식은 모세관 싹의 길이를 동시에 늘릴 수 있게 해준다.
새싹이 혈관신생 자극의 근원을 향해 확장되면서, 내피 세포는 인테그린이라고 불리는 접착 분자를 사용하여 함께 이동합니다.이 새싹들은 세포가 혈관신생 부위로 이동하면서 본격적인 혈관 내강이 되기 위해 고리를 형성합니다.새싹은 하루에 몇 밀리미터의 비율로 발생하며, 혈관 구조의 틈새에서 새로운 혈관이 자랄 수 있게 합니다.기존 혈관을 분할하는 것이 아니라 완전히 새로운 혈관을 형성하기 때문에 혈관신생과는 확연히 다릅니다.
관장성 혈관신생
혈관신생으로도 알려진 관입성 혈관신생은 기존의 혈관을 둘로 쪼개서 새로운 혈관을 형성하는 것이다.
관입은 신생아 쥐에게서 처음 관찰되었다.이런 유형의 혈관 형성에서 모세혈관 벽은 내강으로 확장되어 단일 혈관을 두 개로 나눕니다.관장성 혈관신생에는 4가지 단계가 있다.첫째, 마주보는 두 개의 모세관 벽이 접촉 구역을 설정합니다.둘째, 내피세포 접합부를 재구성하고 혈관 이중층을 천공하여 성장인자 및 세포가 내강에 침투할 수 있도록 한다.셋째, 페리시테와 근섬유아세포로 채워진 접촉부 2개의 새로운 용기 사이에 코어가 형성된다.이 세포들은 혈관 내강 성장을 위한 세포 외 기질을 제공하기 위해 콜라겐 섬유를 핵심에 쌓기 시작합니다.마지막으로 기본구조를 변경하지 않고 코어를 살코기로 한다.삽입은 기존 세포를 재구성하는 것이기 때문에 중요합니다.그것은 내피세포의 수가 증가하지 않고 모세혈관의 수를 크게 증가시킬 수 있게 한다.이것은 새로운 혈관이 [10]발달할 때마다 새로운 세포로 풍부한 미세 혈관을 만들 수 있는 충분한 자원이 없기 때문에 배아 발달에서 특히 중요하다.
생리학
기계적 자극
혈관신생의 기계적 자극은 잘 특징지어지지 않는다.근육 수축이 증가하면 [11]혈관신생을 증가시킬 수 있다는 현재의 지식은 있지만 모세혈관에 작용하는 전단 응력에 관해서는 상당한 논란이 있다.운동 중 산화질소 생산량이 증가했기 때문일 수 있습니다.일산화질소는 혈관의 혈관 확장을 일으킨다.
화학적 자극
혈관신생의 화학적 자극은 여러 성장인자(예: VEGF, FGF)를 포함한 다양한 혈관신생 단백질(예: 인테그린 및 프로스타글란딘)에 의해 수행된다.
개요
| 자극기 | 메커니즘 | |
|---|---|---|
| Fgf. | 내피세포, 평활근세포 및 섬유아세포의 증식과 분화를 촉진합니다. | |
| VEGF | 투과성에 영향을 주다 | |
| VEGFR 및 NRP-1 | 서바이벌 신호 통합 | |
| Ang1 및 Ang2 | 혈관을 안정시키다 | |
| PDGF(BB-Homodimer) 및 PDGFR | 평활근 세포를 배양하다 | |
| TGF-β, 엔도글린 및 TGF-β 수용체 | ↑세포내 매트릭스 생성 | |
| CCL2 | 염증 부위에 림프구를 모집한다. | |
| 히스타민 | ||
| 인테그린αβV3,αβV5(?)[12] 및αβ51 | 결합매트릭스 고분자 및 단백질분해효소 | |
| VE-cadherin 및 CD31 | 내피 접합 분자 | |
| 에프린 | 동맥 또는 정맥의 형성을 결정합니다. | |
| 플라스미노겐활성화제 | 세포외 매트릭스를 리모델링하고 성장 인자를 방출하고 활성화한다. | |
| 플라스미노겐활성화억제제-1 | 인근 선박을 안정시키다 | |
| eNOS 및 COX-2 | ||
| AC133 | 혈관아세포 분화를 조절하다 | |
| ID1/ID3 | 내피 전이 분화를 조절합니다. | |
| 제3종 세마포린 | 내피 세포 접착, 이동, 증식 및 아포토시스를 조절합니다.혈관[13] 투과성 변경 | |
| 노고에이 | 내피 세포의 이동 및 [14]증식을 조절합니다.혈관 [15]투과성을 변경합니다. |
Fgf.
FGF-1(산성 FGF) 및 FGF-2(기본 FGF)의 프로토타입 멤버를 가진 섬유아세포 성장인자(FGF) 패밀리는 현재까지 알려진 22개 이상의 [16]멤버로 구성되어 있다.대부분은 16~18kDa의 단일 사슬 펩타이드이며, 헤파린 및 헤파란 황산염에 높은 친화력을 보인다.일반적으로 FGF는 헤파린 프로테오글리칸의 존재 하에서 세포표면 FGF 수용체에 결합함으로써 다양한 세포기능을 자극한다.FGF 수용체 패밀리는 7개의 멤버로 구성되어 있으며, 모든 수용체 단백질은 FGF 매개 수용체 이합체화 메커니즘에 의해 유도되는 자가인산화를 통해 활성화되는 단일 사슬 수용체 티로신 키나아제이다.수용체 활성화는 유전자 활성화와 세포 분화, 증식, 매트릭스 용해를 포함한 다양한 생물학적 반응을 유도하는 신호 전달 캐스케이드를 발생시켜 내피세포, 섬유아세포 및 평활근세포의 성장에 중요한 승모세포 활성 과정을 개시한다.FGF 패밀리의 모든 22개 멤버 중 유일한 FGF-1은 7개의 FGF 수용체 서브타입 모두에 결합할 수 있으며, FGF 패밀리의 가장 광범위한 작용 멤버이며, FG 수용체 상부의 손상(저독성) 조직에 혈관신생 반응을 탑재하는 데 필요한 다양한 세포 타입에 대한 강력한 마이토겐이다.FGF-1, 이 사실은 주로 새로운 모세 혈관의 형성을 혈관 내피 성장 인자(VEGF), 같은 다른pro-angiogenic 성장 인자에서 FGF-1를 구별하는 내피 세포와 부드러운 근육 세포를 포함 모든 유형의 세포들 동맥이 되는 선박 건설을 위하여 필요한 경우의 확산과 분화를 자극한다.[18][19]
FGF-1 외에 섬유아세포증식인자-2(FGF-2 또는 bFGF)의 가장 중요한 기능 중 하나는 내피세포 증식을 촉진하고 내피세포를 튜브상 구조로 물리적 조직을 형성하여 혈관형성을 촉진하는 것이다.FGF-2는 VEGF 또는 PDGF(혈소판 유래 성장인자)보다 더 강력한 혈관신생인자이지만 FGF-1보다는 덜 강력하다.혈관 성장을 자극하는 것 외에 aFGF(FGF-1)와 bFGF(FGF-2)는 상처 치유에 중요한 역할을 한다.그들은 혈관신생과 과립조직을 발생시키는 섬유아세포와 내피세포의 증식을 자극합니다. 둘 다 혈액 공급을 증가시키고 상처 치유 과정 초기에 상처 공간/캐비티를 채웁니다.
VEGF
혈관내피성장인자(VEGF)는 혈관신생의 주요 원인이며 주어진 네트워크에서 모세혈관의 수를 증가시키는 것으로 입증되었습니다.초기 체외 연구에서 소의 모세혈관 내피세포가 증식하고 VEGF와 bFGF에 의한 자극에 의해 튜브 구조의 징후를 보이는 것으로 나타났으나 결과는 VEGF에서 [20]더 뚜렷했다.VEGF의 상향조절은 운동에 대한 생리반응의 주요 요소이며 혈관신생에서 VEGF의 역할은 혈관손상의 [21][22][23][24]가능한 치료법일 것으로 의심된다.체외 연구는 VEGF가 혈관신생의 강력한 자극제라는 것을 명확히 보여준다. 왜냐하면 이 성장인자의 존재 하에서 도금된 내피세포가 증식하고 이동하며 결국 모세혈관과 [11]유사한 튜브 구조를 형성하기 때문이다.VEGF는 내피세포에서 대규모 시그널링 캐스케이드를 일으킵니다.VEGF 수용체-2(VEGFR-2)에 결합하면 혈관 투과성(eNOS, NO 생성), 증식/생존(bFGF), 이행(ICAMs/VCAMS/MMPs) 및 최종적으로 혈관으로 분화하는 인자의 생산을 자극하는 티로신 키나아제 시그널링 캐스케이드가 시작된다.기계적으로, VEGF는 영향을 받는 부위로의 혈류 증가의 결과로 근육 수축과 함께 상향 조절됩니다.증가된 흐름은 또한 VEGF 수용체 1과 2의 mRNA 생산의 큰 증가를 야기한다.수용체 생산 증가는 근육 수축이 혈관신생과 관련된 신호 캐스케이드의 상향 조절을 야기할 수 있다는 것을 의미한다.NO의 억제는 혈관신생성장인자의 영향을 유의하게 감소시키기 때문에 혈관신생신호 캐스케이드의 일부로서 NO는 혈관신생반응의 주요 요인으로 널리 간주된다.그러나 운동 중 NO의 억제는 혈관신생을 억제하지 않으며, 이는 혈관신생 [11]반응에 다른 요인이 관여함을 나타낸다.
앤지오포이에틴류
Angiopoietins, Ang1 및 Ang2는 마우스 녹아웃 [25]연구에서 입증되었듯이 성숙한 혈관을 형성하기 위해 필요합니다.Ang1과 Ang2는 수용체인 Tie-1과 Tie-2와 결합함으로써 작용하는 단백질 성장인자이다. 이것은 다소 논란이 있지만, 세포 신호는 Tie-2에 의해 주로 전달되는 것으로 보인다. 그러나 일부 논문들은 Tie-1을 통해서도 생리학적 신호를 보여준다.이 수용체들은 티로신 키나아제이다.따라서, 그들은 리간드 결합이 주요 티로신에서 인산화를 시작하는 이합체를 야기할 때 세포 신호를 시작할 수 있다.
MMP
혈관신생의 또 다른 주요 기여자는 매트릭스 메탈로프로테아제(MMP)입니다. MMP는 혈관 벽을 단단하게 유지하는 단백질을 분해하는 데 도움이 됩니다.이 단백질 분해는 내피세포가 발아하는 혈관신생에서 볼 수 있는 간질 매트릭스로 빠져나갈 수 있게 해준다.MMP의 억제는 새로운 모세혈관의 [26]형성을 방해한다.이러한 효소는 세포외 기질을 파괴하면 미세 [11]혈관의 무결성이 감소하기 때문에 혈관 형성 과정에서 고도로 조절됩니다.
Dll4
델타 라이크 리간드 4(Dll4)는 [27][28]혈관신생에 부정적인 조절 효과를 가진 단백질이다.Dl4는 수용체의 노치 패밀리를 위한 막 통과 배위자이다.델타 유사 리간드 4의 결과를 결정하는 데 도움이 되는 많은 연구가 수행되었습니다.한 연구는 특히 Dl4가 종양 혈관 및 [29]성장에 미치는 영향을 평가했다.종양이 자라고 발전하기 위해서는 적절한 혈관 구조가 있어야 한다.VEGF 경로는 종양 성장을 돕는 혈관 구조의 발달에 필수적이다.VEGF와 Dl4의 결합 차단은 종양 전체의 종양 진행과 혈관신생을 억제한다.이는 이러한 내피세포의 증식과 발아를 차단하는 내피세포 시그널링의 장애에 기인한다.이 억제 작용으로 세포는 통제할 수 없을 정도로 성장하지 않기 때문에 이 시점에서 암은 멈춘다.그러나 봉쇄가 풀리면 세포는 다시 [30]증식을 시작할 것이다.
제3종 세마포린
클래스 3 세마포린(SEMA3)은 내피세포 접착, 이동, 증식, 생존 및 주변세포 [13]유입을 조절하여 혈관신생을 조절한다.또한 SEMA3 및 VEGF-A 모두 내피세포에서 뉴로실린 [31][32]수용체 결합을 위해 경쟁하기 때문에 세마포린은 VEGF 매개 혈관신생을 방해할 수 있다.따라서 SEMA3와 VEGF-A의 상대적 발현 수준은 혈관신생에 [13]중요할 수 있습니다.
화학적 억제
혈관신생억제제는 내인성일 수도 있고 약물 또는 식이성분으로서 외부에서 올 수도 있다.
의학에서의 응용
치료 대상으로서의 혈관 형성
혈관신생은 혈관 형성 불량 또는 비정상적인 혈관 [33]구조로 특징지어지는 심장 질환과 같은 질병과 싸우는 표적이 될 수 있다.체내에서 새로운 혈관의 생성을 억제하거나 유도할 수 있는 특정 화합물을 사용하는 것은 그러한 질병과 싸우는 데 도움이 될 수 있다.혈관이 없어야 하는 곳에 혈관이 있으면 조직의 기계적 특성에 영향을 미쳐 기능 상실의 가능성을 높일 수 있습니다.복구 중이거나 신진대사가 활발한 조직에 혈관이 없으면 복구 또는 기타 필수 기능이 저해될 수 있습니다.허혈성 만성 상처와 같은 몇 가지 질병은 혈관 형성이 실패하거나 불충분한 결과이며 혈관의 국소적인 확장에 의해 치료될 수 있으며, 따라서 새로운 영양소를 현장에 가져와 복구를 용이하게 한다.노화와 관련된 황반변성과 같은 다른 질병은 혈관의 국소적인 확장에 의해 생성되어 정상적인 생리 과정을 방해할 수 있다.
혈관신생 원리의 현대적인 임상 적용은 크게 혈관신생 연구가 시작된 항혈관신생 치료와 혈관신생 치료의 두 가지 영역으로 나눌 수 있다.항혈관신생요법이 암과 악성종양을 [34][35]퇴치하기 위해 사용되고 있는 반면, 서양에서는 심혈관질환을 치료하기 위한 대안으로 항혈관신생요법이 연구되고 있다.인간에 대한 혈관신생방법의 첫 번째 적용 중 하나는 관상동맥질환 [18][36][37]치료를 위해 섬유아세포 성장인자 1(FGF-1)을 사용한 독일 실험이었다.
작용 메커니즘과 관련하여 혈관신생에 대한 방법은 크게 세 가지 범주로 구분할 수 있다: 증폭 또는 억제를 위해 관심 있는 유전자를 대상으로 하는 유전자 치료, FGF-1 또는 혈관 내피 성장인자(VEGF)와 같은 혈관신생 성장인자를 주로 조작하는 단백질 대체 치료, 그리고 세포 기반 치료.ich는 특정 세포 유형의 이식을 포함한다.
유전자 치료와 관련하여 여전히 심각하고 해결되지 않은 문제들이 있다.치료용 유전자의 표적 세포의 게놈으로의 효과적인 통합, 바람직하지 않은 면역 반응, 잠재적 독성, 면역 발생성, 염증 반응, 그리고 유전자를 이식하는데 사용되는 바이러스 벡터와 관련된 종양 발생의 위험과 혈관 발생의 유전적 기초의 순수한 복잡성을 포함한다.심장병, 고혈압, 당뇨병, 알츠하이머병과 같은 인간에게 가장 흔하게 발생하는 질환은 많은 유전자의 변이가 복합적으로 작용하여 발생할 가능성이 높으며, 따라서 단일 유전자를 주입하는 것은 그러한 [citation needed]질병에서 크게 이롭지 않을 수 있다.
대조적으로, 혈관신생 단백질 치료는 잘 정의되고 정확하게 구조화된 단백질을 사용하며, 질병 상태에 대한 개별 단백질의 미리 정의된 최적 용량과 잘 알려진 생물학적 효과를 [1]가지고 있다.반면 단백질 요법의 장애물은 분만 방식이다.치료용 단백질이 표적 조직에 들어가기 전에 대사되거나 클리어될 수 있기 때문에 경구, 정맥 내, 동맥 내 또는 근육 내 단백질 투여 경로가 항상 효과적인 것은 아닙니다.세포 기반 혈관 형성 요법은 여전히 연구의 초기 단계이며, 사용하기에 가장 좋은 세포 유형과 용량에 대한 많은 열린 질문들이 있다.
종양 혈관 형성
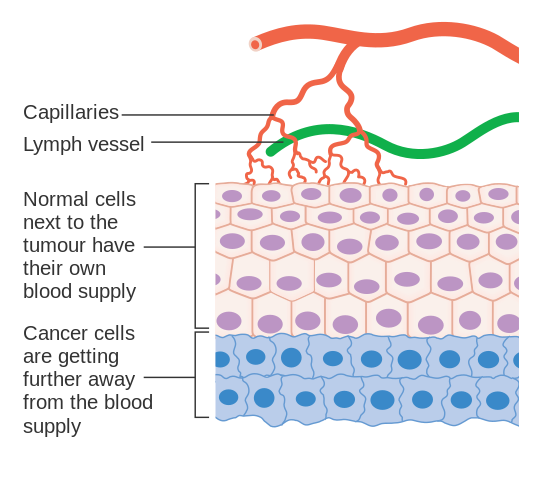
암세포는 조절된 방식으로 분열하는 능력을 상실한 세포이다.악성종양은 돌연변이를 점진적으로 축적하는 빠르게 분열되고 성장하는 암세포 집단으로 구성된다.하지만, 종양은 특정 크기 이상으로 자라기 위해 필요한3 [38][39]산소와 다른 필수 영양소를 공급하기 위해 전용 혈액 공급이 필요합니다.
종양은 다양한 성장인자(예: VEGF)와 단백질을 분비하여 혈관 성장(혈관 형성)을 유도한다.bFGF와 VEGF와 같은 성장 인자는 종양에 모세혈관 성장을 유도할 수 있으며, 일부 연구자들은 종양 확장이 가능하도록 필요한 영양분을 공급한다고 의심하고 있다.종양 혈관은 일반 혈관과 달리 불규칙한 [40]형태로 확장된다.다른 임상의들은 혈관신생이 암세포가 빠르게 분열하면서 분비되는 생물학적 최종 산물을 제거하는 폐기물 통로 역할을 한다고 믿는다.어느 경우든 혈관신생은 종종 볼펜 끝에 있는 금속공 크기라고 불리는 무해한 세포의 작은 클러스터에서 큰 종양으로 이행하기 위해 필요하고 필요한 단계이다.종양의 확산이나 [7]전이를 위해서도 혈관신생이 필요하다.단일 암세포는 확립된 고형종양에서 벗어나 혈관으로 들어가 멀리 떨어진 곳으로 옮겨져 2차 종양의 성장을 시작할 수 있다.이제 증거는 주어진 고형종양의 혈관이 사실 내피세포와 [7]종양세포로 구성된 모자이크 혈관일 수도 있다는 것을 암시한다.이 모자이시티는 종양세포를 혈관구조로 상당히 떨어뜨려 악성종양 [41]환자의 말초혈액에서 순환하는 종양세포의 출현에 기여할 수 있다.이러한 전이의 후속 성장은 또한 영양소와 산소의 공급과 폐기물 처리 경로를 필요로 할 것이다.
내피세포는 암세포보다 유전적으로 더 안정적이라고 오랫동안 여겨져 왔다.이 유전체 안정성은 항혈관신생요법을 이용한 내피세포를 대상으로 하는 화학요법에 비해 항혈관신생요법을 사용하는 데 유리하며, 암세포를 대상으로 하는 화학요법은 빠르게 변이하여 치료에 대한 약물 내성을 획득한다.이러한 이유로, 내피 세포는 그들에게 [42]대항하는 치료법의 이상적인 표적으로 여겨진다.
종양 혈관 형성
혈관신생에 의한 혈관형성의 메커니즘은 돌연변이에 의한 종양세포의 자연분열에 의해 시작된다.혈관신생 자극기는 종양세포에 의해 방출된다.그런 다음 이것들은 이미 확립된 근처 혈관으로 이동하고 그들의 내피 세포 수용체를 활성화시킨다.이것은 혈관 구조에서 단백질 분해 효소의 방출을 유도한다.이 효소들은 혈관의 특정 지점을 목표로 하고 모공을 형성하게 한다.여기가 새로운 혈관이 자라는 지점입니다.종양세포가 혈액공급을 필요로 하는 이유는 약 50-100개의 [43]세포에 해당하는 혈액공급이 확립되지 않으면 지름이 23밀리미터 이상 자랄 수 없기 때문이다.
심혈관 질환에 대한 혈관신생성
혈관신생은 심혈관질환 치료에 훌륭한 치료대상이 된다.이것은 우리 몸이 중요한 장기로의 혈액 공급 감소, 즉 신혈관 형성: 허혈성 [18]모욕을 극복하기 위한 새로운 방부 혈관의 생성에 반응하는 자연스러운 방법의 기초가 되는 강력하고 생리적인 과정이다.말초 동맥 질환 모델뿐만 아니라 심장 허혈 동물 모델에서도 단백질, 유전자 및 세포 기반 치료법으로 많은 임상 전 연구가 수행되었다.이러한 초기 동물 연구에서 재현 가능하고 신뢰할 수 있는 성공은 이 새로운 치료적 접근법이 이러한 장애를 가진 서양의 수백만 명의 환자들을 위한 임상적 이익으로 빠르게 번역될 수 있다는 높은 열정으로 이어졌다.그러나 10년 동안 유전자 및 단백질 기반 치료법을 모두 사용해 저농도 조직과 장기에서 혈관신생을 자극하도록 설계된 임상시험은 실망을 거듭하고 있다.동물에서 인간으로 혈관신생 치료의 전환에 큰 가능성을 제공한 이러한 모든 임상적 판독치가 어떤 식으로든 초기 임상시험에 통합되었지만, FDA는 현재까지(2007년) 혈관신생 치료제의 승인을 위한 주요 끝점은 운동 개선이어야 한다고 주장해왔다.치료된 환자의 [44]수행.
이러한 실패는 이것들이 신생혈관계화를 유도하는 잘못된 분자 목표물이거나, 그것들이 올바르게 조제되고 투여되어야만 효과적으로 사용될 수 있거나, 또는 전체적인 세포 미세환경의 맥락에서 그들의 표시가 효용에 중요한 역할을 할 수 있다는 것을 시사했다.이러한 단백질은 농도, 공간 및 시간 프로파일, 그리고 다른 적절한 [45]인자와 동시에 또는 순차적으로 나타나는 것을 포함한 자연적인 신호 이벤트를 모방하는 방식으로 제시해야 할 수 있다.
운동
혈관신생은 일반적으로 유산소 운동과 지구력 운동과 관련이 있다.동맥신생은 네트워크의 총 흐름량을 크게 증가시키는 네트워크 변화를 발생시키는 반면 혈관신생은 장기간에 걸쳐 더 많은 영양분 공급을 가능하게 하는 변화를 일으킵니다.모세혈관은 최대의 영양소 전달 효율을 제공하도록 설계되어 있기 때문에 모세혈관의 수가 증가하면 네트워크는 같은 시간 내에 더 많은 영양소를 전달할 수 있습니다.모세혈관의 수가 많을수록 네트워크 내 산소 교환도 활발해집니다.이것은 사람이 장기간의 훈련을 계속할 수 있게 해주기 때문에 지구력 훈련에 매우 중요하다.그러나 최대 산소 [11]공급을 증가시키기 위해 지구력 운동에서 모세혈관의 증가가 필요하다는 실험 증거는 없다.
황반변성
VEGF의 과잉 발현은 혈관신생을 자극하는 것 외에도 혈관의 투과성을 증가시킨다.습식 황반변성에서는 VEGF가 망막으로 모세혈관의 증식을 일으킨다.혈관신생의 증가는 또한 부종을 일으키기 때문에 혈액과 다른 망막 액체가 망막으로 새어나와 시력을 잃는다.VEGF 경로를 대상으로 하는 항혈관신생 약물은 현재 이러한 유형의 황반변성을 치료하기 위해 성공적으로 사용되고 있다.
조직공학적 구조
이식된 조직 공학 구조물에 대한 숙주의 혈관의 혈관 형성은 필수적이다.성공적인 통합은 종종 산소와 영양분을 제공하고 [46]임플란트 중앙 부분의 괴사를 방지하기 때문에 구조의 철저한 혈관화에 달려 있습니다.PDGF는 콜라겐 글리코사미노글리칸 스캐폴드에서 [47]혈관화를 안정시키는 것으로 나타났다.
정량화
미세혈관 밀도와 같은 혈관 구조 매개변수를 정량화하는 것은 조직학적 단면에 의한 조직의 선호 염색 또는 제한된 표현으로 인해 다양한 합병증을 가지고 있다.최근 연구는 동물 모델에서 [48]종양 혈관 구조의 완전한 3D 재구성과 전체 종양에서 혈관 구조의 정량화를 보여주었다.
「 」를 참조해 주세요.
레퍼런스
- ^ a b Angiogenesis insights from a systematic overview. New York: Nova Science. 2013. ISBN 978-1-62618-114-4.
- ^ Birbrair A, Zhang T, Wang ZM, Messi ML, Mintz A, Delbono O (January 2015). "Pericytes at the intersection between tissue regeneration and pathology". Clinical Science. 128 (2): 81–93. doi:10.1042/CS20140278. PMC 4200531. PMID 25236972.
- ^ Birbrair A, Zhang T, Wang ZM, Messi ML, Olson JD, Mintz A, Delbono O (July 2014). "Type-2 pericytes participate in normal and tumoral angiogenesis". American Journal of Physiology. Cell Physiology. 307 (1): C25-38. doi:10.1152/ajpcell.00084.2014. PMC 4080181. PMID 24788248.
- ^ "Lymphatic vasculature development" (PDF). www.columbia.edu. Retrieved 17 December 2018.
- ^ Risau W, Flamme I (1995). "Vasculogenesis". Annual Review of Cell and Developmental Biology. 11: 73–91. doi:10.1146/annurev.cb.11.110195.000445. PMID 8689573.
- ^ Flamme I, Frölich T, Risau W (November 1997). "Molecular mechanisms of vasculogenesis and embryonic angiogenesis". Journal of Cellular Physiology. 173 (2): 206–10. doi:10.1002/(SICI)1097-4652(199711)173:2<206::AID-JCP22>3.0.CO;2-C. PMID 9365523.
- ^ a b c Milosevic, Vladan; Edelmann, Reidunn J.; Fosse, Johanna Hol; Östman, Arne; Akslen, Lars A. (2022), Akslen, Lars A.; Watnick, Randolph S. (eds.), "Molecular Phenotypes of Endothelial Cells in Malignant Tumors", Biomarkers of the Tumor Microenvironment, Cham: Springer International Publishing, pp. 31–52, doi:10.1007/978-3-030-98950-7_3, ISBN 978-3-030-98950-7, retrieved 2022-07-13
- ^ John S. Penn (11 March 2008). Retinal and Choroidal Angiogenesis. Springer. pp. 119–. ISBN 978-1-4020-6779-2. Retrieved 26 June 2010.
- ^ 아데어 TH, 몬타니 JP혈관신생.San Rafael (CA): Morgan & Claypool 생명과학; 2010.1장, 혈관신생의 개요.입수처: https://www.ncbi.nlm.nih.gov/books/NBK53238/
- ^ Burri PH, Hlushchuk R, Djonov V (November 2004). "Intussusceptive angiogenesis: its emergence, its characteristics, and its significance". Developmental Dynamics. 231 (3): 474–88. doi:10.1002/dvdy.20184. PMID 15376313. S2CID 35018922.
- ^ a b c d e Prior BM, Yang HT, Terjung RL (September 2004). "What makes vessels grow with exercise training?". Journal of Applied Physiology. 97 (3): 1119–28. doi:10.1152/japplphysiol.00035.2004. PMID 15333630.
- ^ 혈관신생의 억제제일 수 있습니다.
- ^ a b c Mecollari V, Nieuwenhuis B, Verhaagen J (2014). "A perspective on the role of class III semaphorin signaling in central nervous system trauma". Frontiers in Cellular Neuroscience. 8: 328. doi:10.3389/fncel.2014.00328. PMC 4209881. PMID 25386118.
- ^ Rust, Ruslan; Grönnert, Lisa; Gantner, Christina; Enzler, Alinda; Mulders, Geertje; Weber, Rebecca Z.; Siewert, Arthur; Limasale, Yanuar D. P.; Meinhardt, Andrea; Maurer, Michael A.; Sartori, Andrea M.; Hofer, Anna-Sophie; Werner, Carsten; Schwab, Martin E. (9 July 2019). "Nogo-A targeted therapy promotes vascular repair and functional recovery following stroke". Proceedings of the National Academy of Sciences. 116 (28): 14270–14279. doi:10.1073/pnas.1905309116. PMC 6628809. PMID 31235580.
- ^ Rust, Ruslan; Weber, Rebecca Z.; Grönnert, Lisa; Mulders, Geertje; Maurer, Michael A.; Hofer, Anna-Sophie; Sartori, Andrea M.; Schwab, Martin E. (27 December 2019). "Anti-Nogo-A antibodies prevent vascular leakage and act as pro-angiogenic factors following stroke". Scientific Reports. 9 (1): 20040. Bibcode:2019NatSR...920040R. doi:10.1038/s41598-019-56634-1. PMC 6934709. PMID 31882970.
- ^ Ornitz DM, Itoh N (2001). "Fibroblast growth factors". Genome Biology. 2 (3): REVIEWS3005. doi:10.1186/gb-2001-2-3-reviews3005. PMC 138918. PMID 11276432.
- ^ Blaber M, DiSalvo J, Thomas KA (February 1996). "X-ray crystal structure of human acidic fibroblast growth factor". Biochemistry. 35 (7): 2086–94. CiteSeerX 10.1.1.660.7607. doi:10.1021/bi9521755. PMID 8652550.
- ^ a b c Stegmann TJ (December 1998). "FGF-1: a human growth factor in the induction of neoangiogenesis". Expert Opinion on Investigational Drugs. 7 (12): 2011–5. doi:10.1517/13543784.7.12.2011. PMID 15991943.
- ^ Khurana R, Simons M (April 2003). "Insights from angiogenesis trials using fibroblast growth factor for advanced arteriosclerotic disease". Trends in Cardiovascular Medicine. 13 (3): 116–22. doi:10.1016/S1050-1738(02)00259-1. PMID 12691676.
- ^ Goto F, Goto K, Weindel K, Folkman J (November 1993). "Synergistic effects of vascular endothelial growth factor and basic fibroblast growth factor on the proliferation and cord formation of bovine capillary endothelial cells within collagen gels". Laboratory Investigation; A Journal of Technical Methods and Pathology. 69 (5): 508–17. PMID 8246443.
- ^ Ding YH, Luan XD, Li J, Rafols JA, Guthinkonda M, Diaz FG, Ding Y (December 2004). "Exercise-induced overexpression of angiogenic factors and reduction of ischemia/reperfusion injury in stroke". Current Neurovascular Research. 1 (5): 411–20. doi:10.2174/1567202043361875. PMID 16181089. Archived from the original on April 19, 2012.
- ^ Gavin TP, Robinson CB, Yeager RC, England JA, Nifong LW, Hickner RC (January 2004). "Angiogenic growth factor response to acute systemic exercise in human skeletal muscle". Journal of Applied Physiology. 96 (1): 19–24. doi:10.1152/japplphysiol.00748.2003. PMID 12949011. S2CID 12750224.
- ^ Kraus RM, Stallings HW, Yeager RC, Gavin TP (April 2004). "Circulating plasma VEGF response to exercise in sedentary and endurance-trained men". Journal of Applied Physiology. 96 (4): 1445–50. doi:10.1152/japplphysiol.01031.2003. PMID 14660505.
- ^ Lloyd PG, Prior BM, Yang HT, Terjung RL (May 2003). "Angiogenic growth factor expression in rat skeletal muscle in response to exercise training". American Journal of Physiology. Heart and Circulatory Physiology. 284 (5): H1668-78. doi:10.1152/ajpheart.00743.2002. PMID 12543634.
- ^ Thurston G (October 2003). "Role of Angiopoietins and Tie receptor tyrosine kinases in angiogenesis and lymphangiogenesis". Cell and Tissue Research. 314 (1): 61–8. doi:10.1007/s00441-003-0749-6. PMID 12915980. S2CID 2529783.
- ^ Haas TL, Milkiewicz M, Davis SJ, Zhou AL, Egginton S, Brown MD, Madri JA, Hudlicka O (October 2000). "Matrix metalloproteinase activity is required for activity-induced angiogenesis in rat skeletal muscle". American Journal of Physiology. Heart and Circulatory Physiology. 279 (4): H1540-7. doi:10.1152/ajpheart.2000.279.4.H1540. PMID 11009439. S2CID 2543076.
- ^ Lobov IB, Renard RA, Papadopoulos N, Gale NW, Thurston G, Yancopoulos GD, Wiegand SJ (February 2007). "Delta-like ligand 4 (Dll4) is induced by VEGF as a negative regulator of angiogenic sprouting". Proceedings of the National Academy of Sciences of the United States of America. 104 (9): 3219–24. Bibcode:2007PNAS..104.3219L. doi:10.1073/pnas.0611206104. PMC 1805530. PMID 17296940.
- ^ Hellström M, Phng LK, Hofmann JJ, Wallgard E, Coultas L, Lindblom P, Alva J, Nilsson AK, Karlsson L, Gaiano N, Yoon K, Rossant J, Iruela-Arispe ML, Kalén M, Gerhardt H, Betsholtz C (February 2007). "Dll4 signalling through Notch1 regulates formation of tip cells during angiogenesis". Nature. 445 (7129): 776–80. Bibcode:2007Natur.445..776H. doi:10.1038/nature05571. PMID 17259973. S2CID 4407198.
- ^ Segarra M, Williams CK, Sierra ML, Bernando M, McCormick PJ, Meric D, Regino C, Choyke P, T