Cells 2024, 13(19), 1618; https://doi.org/10.3390/cells13191618 - 26 Sep 2024
Viewed by 1412
Abstract
►
Show Figures
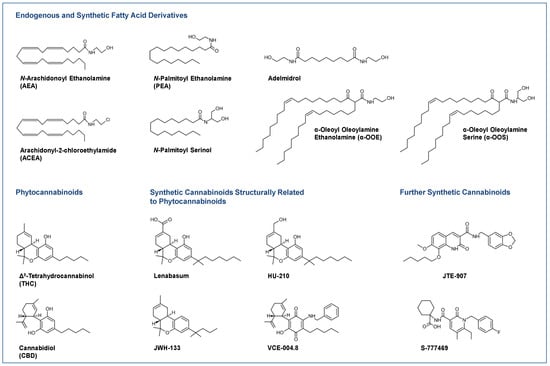
Cannabidiol (CBD) is a major non-psychotropic phytocannabinoid that exists in the Cannabis sativa plant. CBD has been found to act on various receptors, including both cannabinoid and non-cannabinoid receptors. In addition, CBD has antioxidant effects that are independent of receptors. CBD has demonstrated
[...] Read more.
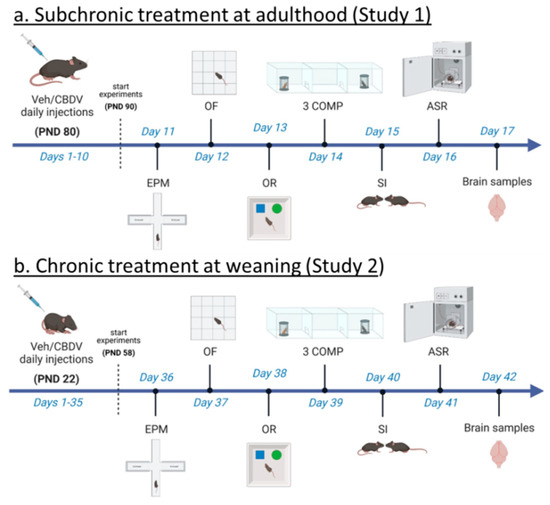
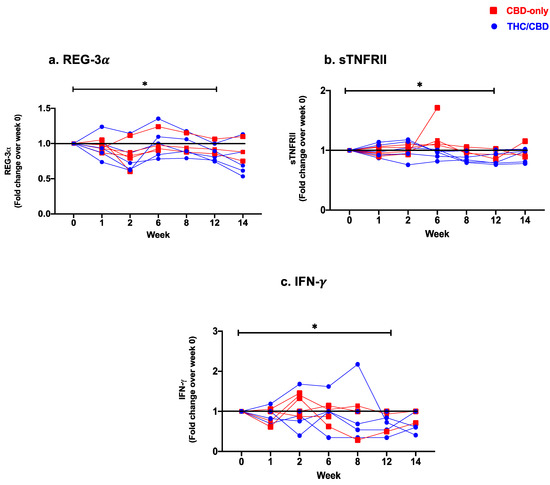
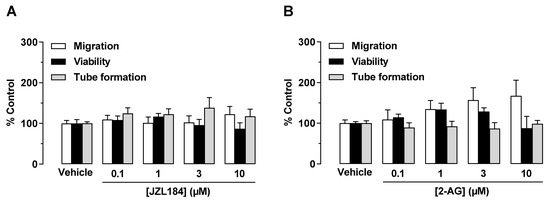
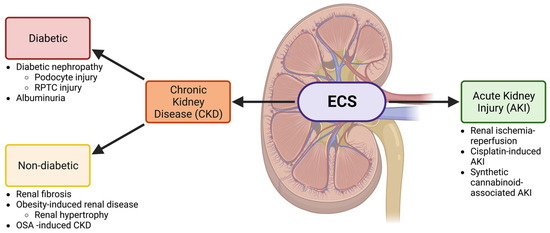
Cannabidiol (CBD) is a major non-psychotropic phytocannabinoid that exists in the Cannabis sativa plant. CBD has been found to act on various receptors, including both cannabinoid and non-cannabinoid receptors. In addition, CBD has antioxidant effects that are independent of receptors. CBD has demonstrated modulatory effects at different organ systems, such as the central nervous system, immune system, and the gastrointestinal system. Due to its broad effects within the body and its safety profile, CBD has become a topic of therapeutic interest. This literature review summarizes previous research findings with regard to the effect of CBD on the gastrointestinal (GI) system, including its effects at the molecular, cellular, organ, and whole-body levels. Both pre-clinical animal studies and human clinical trials are reviewed. The results of the studies included in this literature review suggest that CBD has significant impact on intestinal permeability, the microbiome, immune cells and cytokines. As a result, CBD has been shown to have therapeutic potential for GI disorders such as inflammatory bowel disease (IBD). Furthermore, through interactions with the gut, CBD may also be helpful in the treatment of disorders outside the GI system, such as non-alcoholic liver disease, postmenopausal disorders, epilepsy, and multiple sclerosis. In the future, more mechanistic studies are warranted to elucidate the detailed mechanisms of action of CBD in the gut. In addition, more well-designed clinical trials are needed to explore the full therapeutic potential of CBD on and through the gut.
Full article

Figure 1